Effect of Salinity Stress on Osmoprotectant (Glycine Betaine and Prline) of Two Maize (Zea mays L.). Varieties Cultivated in Sokoto Metropholis
Salinity is one of the principal abiotic stresses that affect plant productivity by inducing osmotic stress. Maize is an economically important cereal crop affected by high salinity. In this study two varieties of maize seedlings (Oba super 2 16-11-kd-155- 159 and Sammaz 37) were subjected to 50, 100 and 150 mM NaCl concentrations. The Root, young and mature leaves were harvested after 21 days of sowing. A significant (P
Introduction
Maize (Zea mays L.) is one of the most important cereal grains grown worldwide in a wider range of environments because of its greater adaptability [1]. Maize is a good source of carbohydrate, vitamins, minerals and dietary fiber [2]. It is mainly used as a food source and has become the most important raw material for animal feed [3]. Maize are glycophytes species and generally show limited growth and development due to soil salinity) [4]. It is an economically important cereal crop and its production is affected by soil salinity in various part of the world.
Salinity is defined as the presence of an excessive concentration of salt in the soil which suppresses plant growth and productivity [5]. Salinity stress induces a multitude of responses in plants including morphological, physiological, biochemical and molecular changes [6]. It causes ionic imbalance, which result in ionic toxicity, osmotic stress, and generation of reactive oxygen species [7]. One of the biochemical changes occurring in plant subjected to environmental stress condition is the production of reactive oxygen species (ROS) [8]. ROS attack protein, nucleic acids and lipid, and the degree of damage depends on the balance between formations of reactive oxygen species [9].
The ability of plants to cope with salinity stress is an important determinant of crop distribution and productivity in many areas, so it is important to understand the mechanisms that confer tolerance to saline environment [10]. Osmolyte could be used as potential growth regulator to improve salinity stress resistance in several plant species [11]. These glycine betainen and proline are very good biochemical makers of stress and their increased levels may attest to potential for remediation [12].
Different organs and/or leaves tissues of different developmental stages may respond differently to salinity stress and trigger specific defences mechanisms, However, differences in oxidative stress and osmoprotectants defences, in different organs and developmental stages of plant species are much less studied, therefore the aim of this study was carried out to evaluate the effect of salinity stress on glycine betaine and proline defense responses of roots and mature leaves of maize cultivars (Oba super 2 16- 11-kd-155-159 and Sammaz 37) cultivated at various NaCl concentration.
Materials and Methods
Plant Material and Stress Treatment
Two maize cultivars (Oba super 2 16-11-kd-155-159 and sammaz-37) were used in this study. The maize cultivars were obtained from Sokoto Agricultural Development Project (SADP), Sokoto State, Nigeria. seed were directly sown into sandy soil (85.9 % sand, pH 6.34) the soil initially contained 0.64 % carbon, 0.060 kg nitrate-nitrogen, 0.3 9 kg sodium, 0.82 kg potassium, 5.8 kg cation exchange capacity, 0.45 kg calcium and 0.50 kg magnesium. Germination was carried out in the Botanical Garden of the Department of Biological Sciences, Usmanu Danfodiyo University, Sokoto. Salinity irrigation water treatments were applied as sodium chloride (NaCl) solution at the following levels (0.00, 50, 100 and 150 mM). Plants were irrigated daily with different NaCl concentration. Each treatment was replicated three times. Roots and mature leaves were harvested after 21 days of salt treatment and samples were immediately taken for analysis.
Hydrogen Peroxide (H2O2)
Hydrogen peroxide content was estimated according to the method of [13]. Exactly 0.5 g fresh tissues of roots and matured leaves were homogenized in 5 ml of 0.1% (w/v) trichloroacetic acid (TCA) in an ice bath. The homogenate was centrifuged at 3000 rpm for 15 min. Then 0.5 ml of the supernatant was made up to 1 ml with 10 mm potassium Phosphate buffer (pH 7.0) to which 1 ml of 1M KI was added. The absorbance was measured at 390 nm against blank in three replicated sample. The blank sample was prepared by 0.4 ml of distilled water and 0.4 ml of working reagent. And 0.4 ml of working reagent was prepared as a standard test.
Determination of Lipid Peroxidation Marker
Malondialdehyde (MDA): MDA was determined using the method of [14]. Exactly 0.5 g each of fresh roots and matured leaves were homogenized in 1 % metaphosphoric acid and then incubated at room temperature for 30 min. 1 ml of 10 % trichloroacetic acid (TCA) was added and centrifuged at 2000 rpm for 15 min and the supernatant was used for the assay. 1 ml of the supernatant was mixed with 1 ml of 5 % thiobarbituric acid (TBA) in the test tubes, followed by addition of equal volume of 40% trichloroacetic acid. The mixture was placed in boiling water for 30 min at 95 0C. The samples were allowed to cool at room temperature and then the absorbance were measured at 532 nm in three replicated sample. The blank sample was prepared by 1.5 ml of distilled water and 1.5 ml of working reagent. And 1.5 ml of working reagent was prepared as a standard test.
Determination Of Osmoprotectants
Glycine Betaine (GB): The glycine betaine level was determined according to the method of [15]. A 0.5 g of the fresh tissues (root and mature leaves) was homogenized for extract preparation with 5 ml of toluene-water mixture (0.05% toluene) and then filtered. After filtration 0.5 ml of extract was mixed with 1 ml of 2 N HCl solution and then 0.1 ml of potassium tri-iodide solution (containing 7.5 g iodine and 10 g potassium iodide in 100 ml of 1 N HCl) was added and shaken in an ice-cold water bath for 30 minute. Then 10 ml of 1, 2-dichloroethane was poured into the test tube and shake gently. Absorbance was measured at 365 nm against blank in three replicated sample. The blank sample was prepared by 0.1 ml of distilled water and 0.1 ml of working reagent. And 0.1 ml of working reagent was prepared as a standard test.
Proline: The proline content was estimated according the method of [16]. Exactly 0.5 g each of roots and matured leaves tissues of maize was homogenized in 5 ml 3% aqueous sulfosalicylic acid in an ice bath. The homogenate was centrifuged at 3000 rpm for 15 min at 4 oC and then filtered. 2ml of the filtrate was added to 2 ml acid ninhydrin and 2 ml of glacial acetic acid in a test tube. The mixture was boiled for 1 hour and then later the test tube was placed in an-ice cold water for the termination of the reaction. 4 ml of toluene was added to the reaction mixture and stirred for 30 sec. The toluene layer was separated and warmed at 37°C and the absorbance was measured at 520 nm against blank sample in three replicated. The blank sample was prepared by 2.0 ml of distilled water and 2.0 ml of working reagent. And 2.0 ml of working reagent was prepared as a standard test.
Statistical Analysis
Statistical analysis was carried out using the Instat software. Parameters were analysed statistically by one way analysis of variance (ANOVA). Results were presented as Means ± SEM and significant difference between means (p<0.05) was established using the Duncan multiple range test.
Results
Hydrogen Peroxide Levels in Salt-Stressed Maize Cultivars
Increase in salinity stress resulted a significant increase in hydrogen peroxide (H2O2) content in roots at 50 mM NaCl and young leaves at 50 and 100 mM NaCl of Oba super 2. In sammaz 37 variety significant increase in hydrogen peroxide content was observed only at 50 mM NaCl in young and matured leaves (Figure 1).
Malondialdehyde (MDA) Levels of Maize Seedlings Under Salinity Stress
MDA increased significantly (p<0.05) in root, mature leaves and young leaves (Figure 2). The effect of salt stress was observed at highest concentration (100 and 150 mM NaCl) in roots, matured and young leaves of both the varieties investigated.
Glycine Betaine Content of Maize Seedling under Salinity Stress
Figure 3 shows that salt stress resulted in no significant increase in GB level in root and mature leaves of Oba super 2 while in the young leaves there is slight decrease in GB level. Sammaz 37 variety shows no significant increase in GB by increasing salt concentration in all the tissues examined. Proline Levels in Salt Stress Maize Cultivars Levels of proline in root of oba super 2 variety decreased at all salt regimes when compared with control, but in young leaves proline levels decrease significantly only at 150 mM NaCl. While in the mature leaves increase in salinity resulted to a decrease in proline level at all salt regime when compared to the control Figure 4. In roots of sammaz 37 the decrease in proline levels was observed by increasing salt concentration. In young leaves Proline level shows significant decreased (p˂0.05) only at 150 mM NaCl whereas in the mature leaves proline levels increased significantly at 50, 100 and 150 mM NaCl.
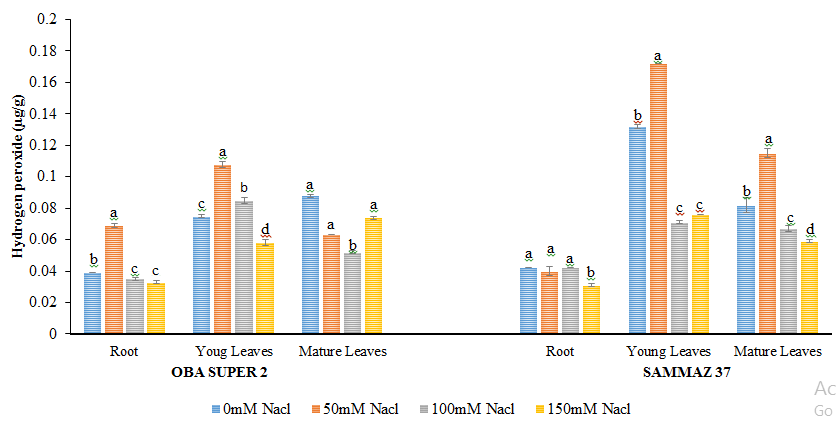
Figure 1: Hydrogen Peroxide levels (µg/g) of two varieties of maize exposed to the same level of different concentrations of NaCl for 21 days. Values are expressed as mean ± SEM of three replicates. Significant difference between the means was determined using Duncan’s test at (p < 0.05), mean in the column followed by the same letter (s) are not significantly difference at 5% level.
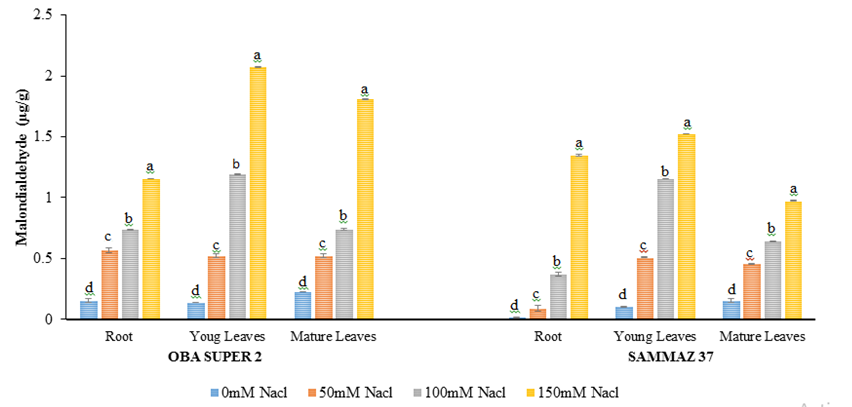
Figure 2: Malondialdehyde levels (µg/g) of two varieties of maize exposed to the same level of different concentrations of NaCl for 21 days. Values are expressed as mean ± SEM of three replicates. Significant difference between the means was determined using Duncan’s test at (p < 0.05), mean in the column followed by the same letter (s) are not significantly difference at 5% level.
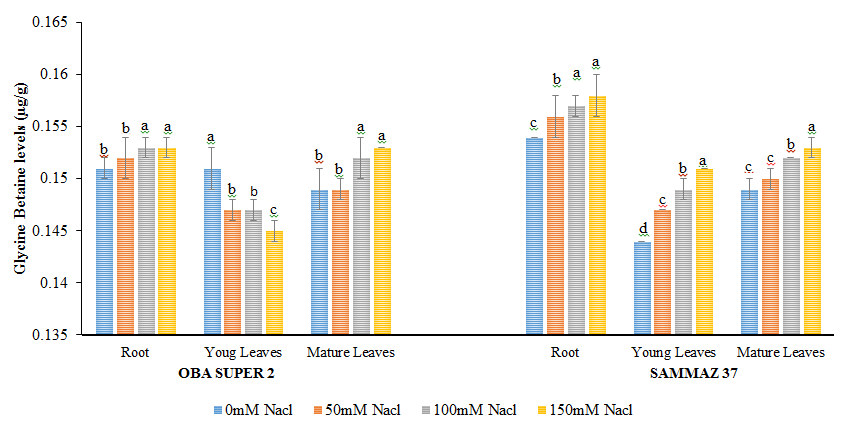
Figure 3: Glycine Betaine levels (µg/g) of two varieties of maize exposed to the same level of different concentrations of NaCl for 21 days. Values are expressed as mean ± SEM of three replicates. Significant difference between the means was determined using Duncan’s test at (p < 0.05), mean in the column followed by the same letter (s) are not significantly difference at 5% level.
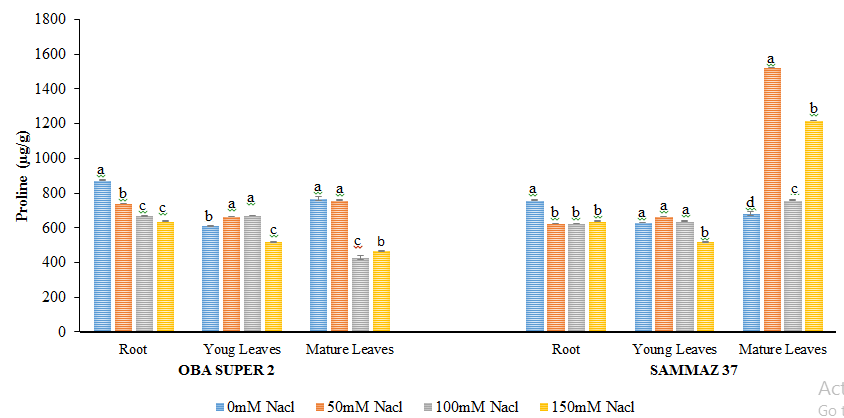
Figure 4: Proline levels (µg/g) of two varieties of maize exposed to the same level of different concentrations of NaCl for 21 days. Values are expressed as mean ± SEM of three replicates. Significant difference between the means was determined using Duncan’s test at (p < 0.05), mean in the column followed by the same letter (s) are not significantly difference at 5% level.
Discussion
Adverse effects of Salinity in plants include reduction in overall growth and productivity. To contribute the understanding of the mechanisms underlying salinity stress responses, maize seedlings were exposed to three different salinity levels (50, 100 and 150 mM), and investigated the responses in roots and mature leaves (M.L). Salinity tolerance is an important trait for plants such as maize that grow in arid and semi-arid areas where water has high concentrations of salts [17]. Malondialdehyde and Hydrogen peroxide: Salinity has been reported to induce oxidative stress in different plants and tissues [4, 18]. Under salinity Stress, the levels of ROS increases in the plant tissues as a result of irregularities in the electron transport chain and accumulation of photo reducing power. One of the early changes in plants physiology and metabolism during abiotic stresses such as salinity is the production of reactive oxygen species (ROS) which include O2 -, H2O2, and OH.- Among these, H2O2 appears to be one of the earliest stress signalling factor, and a stable ROS intermediate. This study observed increase in MDA and hydrogen peroxide (H2O2) in root and mature leaves of both the two varieties. Tatar, et al. have also reported high MDA content with increase in the degree of salt stress in wheat. However, report by Weisany, et al. showed an increase in MDA and H2O2 levels in soybeans. Sammaz 37 is more tolerant than Oba super 2 variety, because sammaz 37 showed lower level of MDA which indicate tolerant of the plant against environmental stresses compared to Oba super 2, Glycine betaine: salinity induced adjustment in osmolarity of plants involve of osmoprotectants such as glycine betaine and proline. GlyBet alleviated lipid peroxidation and facilitated the protection of membrane functions under salinity stress. This facilitation could be attributed to the GlyBet induced antioxidant responses that support the plant from oxidative. In this study progressive increase in GlyBet levels was observed in root, mature and young leaves of both the two variety, similar results showed increased in Glycine betaine reported in other plants like safflower by Erda, et al., wheat, tomato and purslane under salinity stress condition [19].
Slightly decreased in proline level was observed in the present study, this is in contrast to many previous reports where proline (Pro) levels were increased under Salinity stress to decrease the cellular water potential and improve water uptake, and possibly scavenge ROS molecules. However, similar to this study Sorghum bicolor leaves showed no significant change in Pro content under salinity stress while in S. sudanense proline decreased slightly with salinity. Moreover, different maize varieties differed in Pro accumulation under salinity stress. Variety K3615.1 did not accumulate significant amounts of Pro under 50–100mM NaCl, while K3653.2 showed decreased Pro content under elevated salinity.
Conclusion
This study revealed that Salt induced osmotic stress also seems to be countered by increased production of the osmolyte (Glycine betaine and proline). These accumulation of osmolytes could improve salinity tolerance in both the two varieties of maize.
References
-
Kogbe MB, Adediran C (2003) Soil compaction modifies characteristics of seminal Maize roots. Plant Soil Environment 55: 1-10.
-
Allen MM, Ort DM (2011) Growth and nutrients status of wheat as affected by ascorbic acid and water salinity. Nature and Science 9: 64-69.
-
Pooja S, Rajesh K (2015) Soil Salinity: A Serious Environmental Issue and Plant Growth Promoting Bacteria as One of the Tools for Its Alleviation. Saudi Journal of Biological Science 22(2): 123-131.
-
Ashraf M, Harris PJC (2004) Potential biochemical indicators of salinity tolerance in Plants. Plant Science 166(1): 3-16.
-
Zaki F (2011) The determinat of salinity tolerance in maize (Zea mays L.). University of Groningen, Netherlands, pp: 11-15.
-
Ambede JG, Netondo GW, Mwai GN Musyimi DM (2012) NaCl salinity affects germination, growth, Physiology, and biochemistry of bambara groundnut. British Journal of Plant Physiology 24(3): 151-160.
-
Chaparzadeh N, D Amico ML, Khavari-Nejad RA, Izzo R, Navari-Izzo F (2004). Antioxidative responses of Calendula officinalis under salinity conditions. Plant Physiology and Biochemistry 42(9): 695-70.
-
Munne-Bosch S (2005) The role of alpha-tocopherol in plant stress tolerance. The Journal of Plant Physiology 162(7)**:** 743-748.
-
Verheul MJ, Vanhassel PR, Stamp P (2002) Comparison of maize inbred lines Differing in low-temperature tolerance - effect of acclimation at suboptimal temperature On chloroplast functioning. Annals of Botany 76(1): 7-14.
-
Gilbert GA, Gadush MV, Wilson C, Madore MA (1998) Amino acid Accumulation in sink and source tissues of Coleus blumei Benth During salinity stress. Journal of Botany 49(318): 107-114.
-
Gunes A, Inal A, Alpaslam M, Erslan F, Bagsi EG, et al. (2007) Salicylic acid induced Changes on some physiological parameters symptomatic for oxidative stress and mineral Nutrition inmaize (Zea mays L.) grown under salinity. Journal of Plant Physiology 164(6): 728-736.
-
Vangronsveld J, Clijsters H (1994) Toxic effects of metals. In: Plants and the Chemical Elements. Farago ME (Ed.), Biochemistry, Uptake, Tolerance and Toxicity pp: 150- 177.
-
Velikova V, Yordanov I, Edreva A (2000) Oxidative stress and some antioxidant systems in acid rain-treated bean plants: protective role of exogenous polyamines. Plant science 151(1): 59-66.
-
Hodges DM, DeLong JM, Forney CF, Prange RK (1999) Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 207: 604-611.
-
Grieve CM, Grattan SR (1983) Rapid assay for determination of water soluble quaternary ammonium compounds. Plant and soil 70: 303-307.
-
Bates LS, Waldren RP, Teare ID (1973) Rapid determination of free proline for water-stress studies. Plant and soil 39: 205-207.
-
Yaish MW, Kumar PP (2015) Salt tolerance research in date palm tree (Phoenix dactylifera L.), Past, present, and future perspectives. Front Plant Science 6: 348.
-
Chawla S, Jain S, Jain V (2013) Salinity induced oxidative stress and antioxidant system in salt-tolerant and salt- sensitive cultivars of rice (Oryza sativa L.). Journal of Plant Biochemistry Biotechnology 22**:** 27-34.
-
Yaish M (2017) Epigenetic Modifications Associated with Abiotic and Biotic Stresses in Plants: An Implication for Understanding Plant Evolution. Front Plant Science 8: 1983.
- Evaluation of Proximate and Mineral Compositions of Momordica charantia L. (Cucurbitaceae)
- Targeting Superbugs: Efficacy of Bacteriophage Therapy against Antibiotic-Resistant Pseudomonas Aeruginosa in Urinary Tract Infections
- Genetic Insights into Prepubertal Gynecomastia: A Comprehensive Analysis of a Rare 45,X[2]/ 46,X, + mar[28] Karyotype
- The Efficiency of Biological Treatment Plants in Some Private Hospitals in the City of Basra, Iraq
- Exploring the Combined Efficacy of Carvacrol and Friedelin against Multi-Drug Resistant Bacteria in Upper and Lower Respiratory Tract Infections
- Isolation, Identification and Comparative Analysis of Oral Microbial Communities in Smokers and Non-Smokers: A Scientific Investigation