Cannabidiol Decreases Neuropathic Pain in Mice by Release of Opioid Peptides
Background: More than 100 phytocannabinoids have already been identified present in varying percentages of composition in the various plants of the genus Cannabis, including cannabidiol (CBD). Some evidences have demonstrated the effectiveness of CBD in treating neuropathic pain. We evaluated the participation of the opioid system in peripheral antinociception induced by the administration of CBD in the face of neuropathic pain induced by constriction of the sciatic nerve. Methods: Neuropathic pain was induced with the constriction of the sciatic nerve while the nociceptive threshold was measured using the pressure test of the mice paw. All drugs were administered by intraplantar injections in the right hind paw of mice. Results: Bestatin, an inhibitor of the enzyme aminopeptidase N, which degrades opioid peptides, potentiated the antinociceptive effect of the lowest administered dose of CBD (5 μg/paw). The antinociceptive effect of CBD was reversed by the administration of naloxone, a non-selective antagonist for opioid receptor. The selective antagonist for μ opioid receptor, clocinnamox, reversed the antinociceptive effect of CBD. Naltrindole, a selective antagonist for δ opioid receptors, and nor-binaltorphymine, a selective antagonist for κ opioid receptors, partially reversed the antinociception induced by CBD. Conclusions: Our data suggest the participation of the opioid system in CBD-induced peripheral antinociception in the model of neuropathic pain induced by sciatic nerve constriction in mice.
Introduction
More than 100 phytocannabinoids have already been identified present in varying percentages of composition in the various plants of the genus Cannabis, with cannabidiol (CBD) being the most abundant non-psychoactive phytocannabinoid [1]. CBD has its therapeutic potential evidenced, in animal models, for the treatment of various diseases such as anxiety, depression, neoplasms, cardiac arrhythmias, psychosis and pain [2]. Due to its low affinity for cannabinoid receptors, data from the literature indicate that CBD has affinity for many other molecular targets, including receptors for glycine, transient potential receptor family (TRP), PPARγ receptors, 5-HT1A receptors, and opioid receptors [2, 3, 4]. The objective of this study was to evaluate the participation of the opioid system in CBD-induced antinociception against neuropathic pain induced by sciatic nerve constriction.
Materials and Methods
Animals
Male Swiss mice (30-40g) from the Center for Experimental Animals at Institute of Biological Sciences (ICB) of the Federal University of Minas Gerais (CEBIO-ICB/ UFMG) were used in the experiments. Housed in standard cages and kept at a constant temperature of 23 to 25°C with a 12h light-dark cycle, the mices had free access to food and water. After the experimental procedures the animals were killed with an intraperitoneal injection of a lethal dose of a mixture of ketamine (300 mg/kg - Cayman Chemical) and xylazine hydrochloride (25 mg/kg - Sigma Aldrich), in concentrations 3 times higher than the anesthetic concentration (180 mg / kg and 24 mg / kg of body weight respectively). Following the guidelines of the ARRIVES and the National Research Council›s Guide for the Care and Use of Laboratory Animals, and submitted to the Ethics Committee on Animal Experiments (CETEA) of the Federal University of Minas Gerais (UFMG) with the Protocol nº 57/2017, the project was approved.
Neuropathic Pain Induction Model: Sciatic Nerve Constriction (SNC)
The animals were anesthetized with 10% ketamine hydrochloride solution (Dopalen, Vetbrands, Brazil) at dose 100 mg/kg and 2% xylazine hydrochloride (Dopaser, Calier, Brazil) at dose 5 mg/kg, with an intraperitoneal injection in a final volume of 150 μl of solution. The surgical technique used was adapted from the method described by Yalcin I, et al. [5]. After the application of the anesthetics, we laid each supine mouse down on the surgical table. We then performed a local epilation and by using tweezers and sharp-edged scissors, made a small incision parallel to the femur. We then dilated the muscles to locate and expose the sciatic nerve.
Unlike the procedure performed by Yalcin I, et al. [5], who implanted a polyethylene cuff in the sciatic nerve, in our model we made two loose ligatures around the nerve separated one from the other at a distance of approximately 2 millimeters, and tied them with a loose resinous flexible thread. Afterwards, we sutured the surgical wound with sterile 5.0-needled surgical nylon thread. Then we divided the experimental groups into “naive”, “operated”, “wireless sham” and “wired sham” groups. “Naïve” animals did not undergo any surgery. The sham-operated groups were submitted to all experimental procedures performed in the “operated” group, except for sciatic nerve constriction.
Hyperalgesia Measurements
Hyperalgesia was measured according to Randall and Selitto [6] and adapted to mice by Kawabata, et al. [7]. An algesimeter equipped with a cone-shaped paw-presser with a rounded tip was used (Ugo-Basile, Italy) in order to apply a linearly increasing force to the hind paw. The weight in grams (g) required to elicit the nociceptive response of paw flexion was determined to be the nociceptive threshold. A cutoff value of 160 g was used to reduce the possibility of damage to the paws. The nociceptive threshold of each animal was determined after the constriction of the sciatic nerve and before the administration of each drug. The threshold is always measured three times, the result being the mean of these measurements.
Experimental Procedure
Cannabidiol (THC Pharm, purity >98%) dissolved in Tocrisolve 5% in sterile aqueous solution of sodium chloride (NaCl) at 0.9% (physiological saline); Bestatin (Tocris), an inhibitor of the enzyme aminopeptidase N, an enzyme that degrades endogenous opioid peptides, dissolved in physiological saline; Naloxone; (Sigma), non-selective antagonist for opioid receptor, dissolved in physiological saline; Clocinnamox (Tocris), selective antagonist for μ opioid receptors, dissolved in physiological saline; Naltrindole (Tocris), selective antagonist for δ opioid receptors, dissolved in physiological saline; Nor-binaltorphymine (Tocris), a selective antagonist for κ opioid receptors, dissolved in physiological saline. The administration protocols were recorded in the legends of all graphics. The nociceptive threshold was always measured three times, in the right hind paw of the animals, and the result was the mean of these measurements. All drugs were administered by intraplantar injections in the right hind paw of mice, in a volume of 20 μl.
Statistical Analyses
Results obtained were analyzed by using the Graph Pad Prism 8.0.2 and presented as mean ± SEM of the measures and the analysis of variance two-way (ANOVA) followed by the Bonferroni post-test used to find out the value of the differences between the averages (multiple comparisons), being such a difference considered significant when P values were less than 0,05.
Results
In our data, bestatin (400 μg/paw), an inhibitor of the enzyme aminopeptidase N, involved in the degradation of opioid peptides, potentiated the peripheral antinociception induced by CBD at its lowest dose (5μg/paw) in the animals 15 days after constriction of the sciatic nerve (Figure 1). Bestatin, when administered alone, did not cause any change in the nociceptive threshold of the animals.
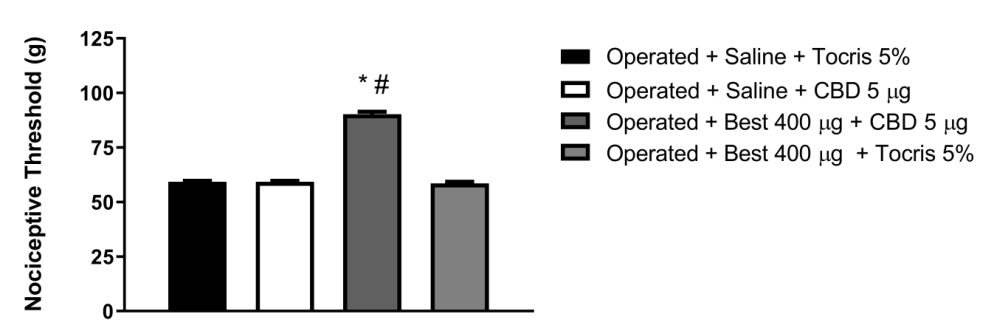
Figure 1: Intraplantar administration of Bestatin (Best) potentiates the peripheral antinociceptive effect of cannabidiol. Bestatin (μg) was administered 30 minutes before cannabidiol (5μg/paw) in the animals 15 days after constriction of the sciatic nerve. Each column represents the mean ± E.P.M. (n = 5). * and # indicate a statistically significant difference (p < 0.05) in relation to the Operated + Saline + Tocris 5% and Operated + Saline + CBD 5 μg group injected as controls, respectively, according to the one-way ANOVA, followed by Bonferroni`s post hoc test.
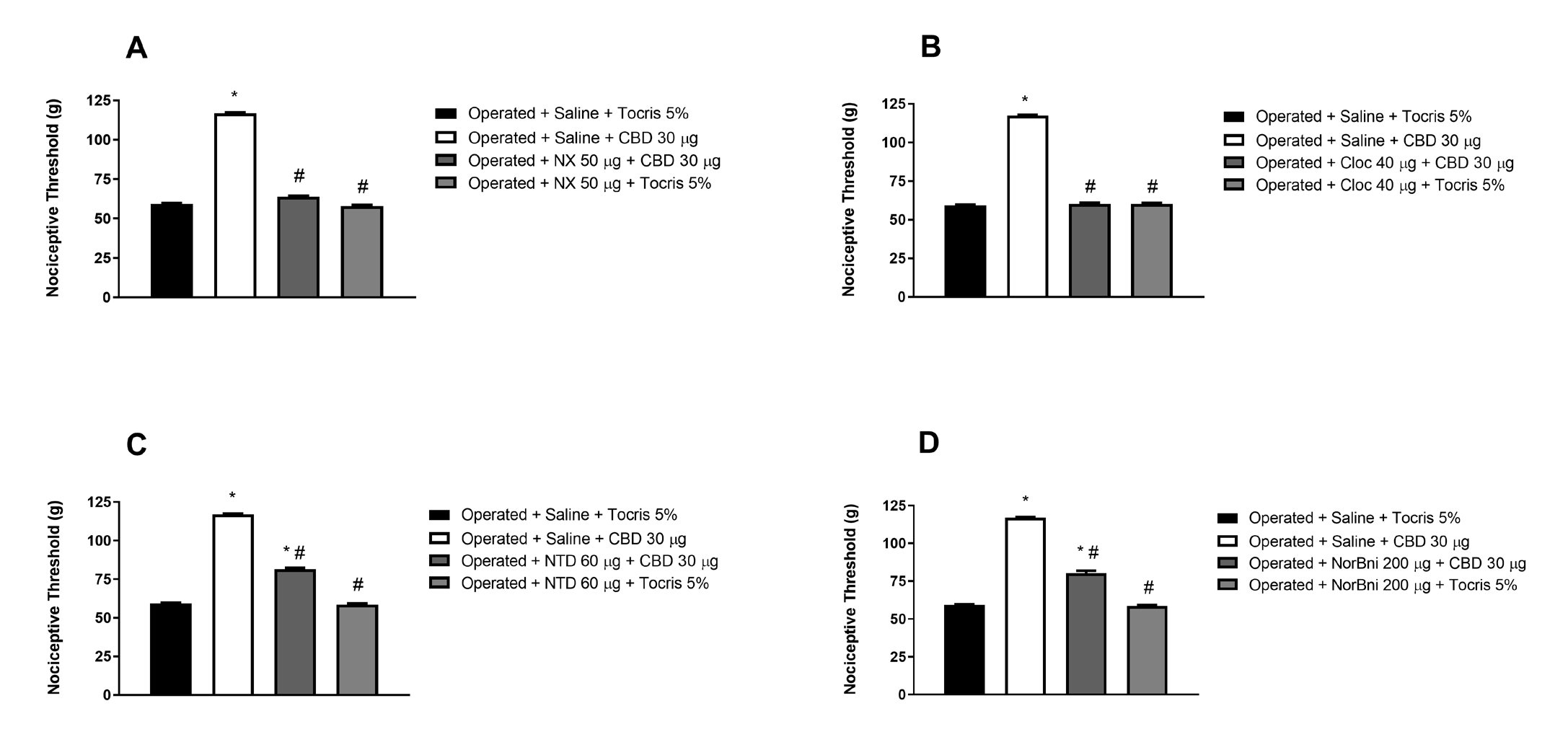
Figure 2: Reversal of the peripheral antinociceptive effect of cannabidiol by intraplantar administration of selective opioid receptor antagonists. Naloxone (NX, A), clocinnamox (Cloc, B), naltrindole (NTD, C) and nor-binaltorphymine (NorBni, D) were administered 30 minutes before cannabidiol (30 μg/paw) in the animals, 15 days after sciatic nerve constriction. Each column represents the mean ± E.P.M. (n = 5). * and # indicate a statistically significant difference (p < 0.05) in relation to the Operated + Saline + Tocris 5% and Operated + Saline + CBD 30 μg group injected as controls, respectively, according to the one-way ANOVA, followed by Bonferroni`s post hoc test.
As shown in (Figures 2A and 2B), respectively, naloxone (50 μg/paw), a non-selective antagonist for opioid receptor, and clocinnamox (40 μg/paw), a selective antagonist for μ opioid receptors, reversed CBD-induced peripheral antinociception (30 μg/paw) in animals 15 days after sciatic nerve constriction. The antagonists when administered alone did not cause alteration in the nociceptive threshold of the animals.
On the other hand, (Figures 2C and 2D) show that naltrindole (60 μg/paw), a selective antagonist for δ opioid receptors, and nor-binaltorphymine (200 μg/paw), a selective antagonist for κ opioid receptors, partially reversed the peripheral antinociception induced by CBD (30 μg/paw) in the animals 15 days after constriction of the sciatic nerve. The antagonists when administered alone did not cause alteration in the nociceptive threshold of the animals.
Discussion
We have demonstrated in previous work that CBD has a peripheral antinociceptive effect against neuropathic pain induced by sciatic nerve constriction [4]. The mechanism of action of CBD is complex, and it involves several molecular targets. Among other physiological functions, it plays a role in the descending control of the nociceptive pathway. This involves the indirect activation of CB1 and CB2 cannabinoid receptors, inhibition of anandamide (AEA) reuptake, increased degradation of AEA, and activation/desensitization of vanilloid receptors type 1 (TRPV1). Additionally, CBD activates glycine receptors and serotonergic receptors type 5-HT1A [2, 4].Regarding the interaction between CBD and the opioid system, research has demonstrated, using kinetic binding studies, that CBD is an allosteric modulator of the μ and δ opioid receptors [3]. With regard to this interaction in pain models, Neelakantan H, et al. [8] reported that a CBD- opioid combination presents synergistic analgesia in the acetic acid induced writhing test.
The mechanism through which the opioid system produces analgesia is related to the action of endogenous ligands that bind with varying affinities to opioid receptors. β-endorphin exhibits a similar affinity for μ and δ receptors but has a low affinity for κ receptors. Enkephalins, on the other hand, have a high affinity for δ receptors, a low affinity for μ receptors, and no affinity for κ receptors. Dynorphin A and dynorphin B display a high affinity for κ receptors but also exhibit significant affinity for μ and δ receptors [9].
By using as a pharmacological tool bestatin, an inhibitor of the enzyme aminopeptity-N, which in our studies potentiated the peripheral antinociception induced by cannabidiol at its lowest dose, we can suggest the participation of endogenous opioid peptides in the effect of CBD, even without describing how this release process would happen Previous studies have reported that opioid peptides from immune cells can activate opioid receptors present in peripheral nerve termination in inflamed paw of rats [10, 11]. Another likely source of these peptides are resident cells, such as keratinocytes, which in addition to constitutively expressing CB1 and CB2 cannabinoid receptors [12], are a source of β-endorphin [13], dynorphin A [14], Met and Leucine-enkephalin [15] released through cellular activation. Although classically the CB1 and CB2 receptors are known for their coupling to the inhibitory transduction G protein (Gi), whose activation inhibits adenylate cyclase, promotes opening of potassium channels (K+) and closure sodium channels (Na+), which results in decreased cellular activity [16]. And Glass M, et al., Bonhaus DW, et al. [17, 18] bring evidence that CB1 receptor can also bind to the excitatory transduction G protein (Gs), which promotes the stimulation of adenylate cyclase and opening calcium channels, resulting in increased cellular activity. The activation of G protein-coupled receptors (GPCR) releases the α and βγ subunits, which will activate their respective effectors. The G-protein βγ dimers play an important role in signal transduction, and G βγ can stimulate the activities of fospolipase C (PLC) and mitogen-activated protein kinase (MAPK) pathway, increaseing the intracellular calcium concentration [19].
A probable mechanism of action for the antinociceptive effect of CBD may be the activation of CB1 and CB2 in immune cells and keratinocytes, inducing the release of opioid peptides, and these, in turn, activate opioid receptors (Gi), inhibiting primary afferent neurons and, consequently, reducing pain. Advancing in our experiments we evaluated the participation of opioid receptors from the use of naloxone, a non-selective antagonist for opioid receptors, which antagonized the antinociceptive response induced by CBD. To verify the selectivity of the opioid receptors involved, we used as pharmacological tools selective antagonists for these receptors. In response, we found that clocinnamox, a selective antagonist for type μ opioid receptors, reversed cannabidiol-induced peripheral antinociception. Naltrindole, a selective antagonist for type δ opioid receptors, and nor- binaltorphymine, a selective antagonist for type κ opioid receptors, partially reversed the peripheral antinociception induced by CBD. These data suggest the participation of opioid receptors in CBD-induced peripheral antinociception. The literature shows us that CBD evokes cytosolic elevations of Ca2+, both by releasing this ion from Ca2+ deposits of the endoplasmic reticulum (ER) in cultured neurons [20] and from mitochondria in oligodendrocytes and hippocampal neurons [21, 22].
A possible explanation for our results could be that CBD induces excitatory transduction via Gs when activating CB1 receptors, leading to an increase in intracellular levels of Ca2+.
When CBD activates CB2 receptors, it stimulates the activity of the βγ subunit of the Gi protein, which, in turn, enhances the activity of PLC and MAPK, further increasing Ca2+ levels. This intracellular increase in calcium may be associated with the release of endogenous opioids by immune cells and keratinocytes [19, 23]. Additionally, CBD could also increase intracellular Ca2+ concentrations by mobilizing this ion from the ER and mitochondria stock, thereby amplifying the production and release of opioids, causing potent analgesia. Our hypothesis can be supported by work of Ibrahim MM, et al. [13] who suggested that the activation of CB2 receptors by their agonist AM1241 produces antinociception by β-endorphin release. The result of Katsuyama S, et al. [24] also corroborate our hypothesis since antinociception produced by intraplantar injection β -caryophyllene is mediated by activation of CB2 receptors, which stimulates the local release from keratinocytes of the endogenous opioid β-endorphin. The release of β-endorphin appears to play a critical role in CB2 receptor-mediated antinociception as both the effects of AM1241 and β-caryophyllene were abolished after the use of an antiserum that sequesters β-endorphin.
Conclusion
In our study, we found that bestatin, an inhibitor of aminopeptidase N, which degrades opioid peptides, enhanced the peripheral pain-relieving effects of CBD at a low dose in animals with sciatic nerve constriction. Naloxone, a non-selective antagonist for opioid receptors and clocinnamox, a selective antagonist for µ-opioid receptor, reversed CBD-induced pain relief. However, naltrindole and nor-binaltorphymine, selective antagonists for δ and κ opioid receptors respectively, partially reversed the effect. Notably, when administered alone, these antagonists had no impact on the pain threshold, highlighting the role of the opioid system in mediating pain relieving by CBD effects in this context. Then, the present data suggest the participation of endogenous opioid peptides and opioid receptors in CBD-induced peripheral antinociception in the model of neuropathic pain induced by sciatic nerve constriction.
References
-
Pertwee R (2014) Handbook of Cannabis.
-
Ligresti A, De Petrocellis L, Marzo V Di (2016) From Phytocannabinoids to Cannabinoids Receptors and Endocannabinoids: Pleiotropic Physiological and Pathological Romes through Complex Pharmacology. Physiol Rev 96(4): 1593-1659.
-
Ibeas Bih C, Chen T, Nunn AVW, Bazelot M, Dallas M, et al. (2015) Molecular Targets of Cannabidiol in Neurological Disorders. Neurotherapeutics 12(4): 699-730.
-
Diniz DA, da Costa Oliveira, C Fonseca FCS, Almeida DL, Pereira WVC, et al. (2023) Peripherally Injected Canabidiol Reduces Neuropathic Pain in Mice: Role of the 5-Ht1a and Trpv1 Receptors. Biochem Biophys Res Commun 660: 58-64.
-
Yalcin I, Megat S, Barthas F, Waltisperger E, Kremer M, et al. (2014) The Sciatic Nerve Cuffing Model of Neuropathic Pain in Mice. Journal of Visualized Experiments 16(89): 51608.
-
Randall L, Selitto J (1957) A Method for Measurement of Analgesic Activity on Inflamed Tissue. Arch Int Pharmacodyn CXI: 111(4): 409-419.
-
Kawabata A, Nishimura Y, Takagi H (1992) L-Leucyl- L-Arginine, Naltrindole and D-Arginine Block Antinociception Elicited by L-Arginine in Mice with Carrageenin-Induced Hyperalgesia. Br J Pharmacol 107(4): 1096-1101.
-
Neelakantan H, Tallarida RJ, Reichenbach ZW, Tuma RF, Ward SJ, et al. (2015) Distinct Interactions of Cannabidiol and Morphine in Three Nociceptive Behavioral Models in Mice. Behavioural Pharmacology 26(3): 304-314.
-
Janecka A, Fichna J, Janecki T (2004) Opioid Receptors and their Ligands. Curr Top Med Chem 4(1): 1-17.
-
Stein C, Hassant AHS, Przewlocki R, Gramsch C, Peter K, et al. (1990) Opioids From Immunocytes Interact with Receptors on Sensory Nerves to Inhibit Nociception in Inflammation. Proc Natl Acad Sci USA 87(15): 5935- 5939.
-
Mousa SA, Zhang Q, Sitte N, Ji RR, Stein C (2001) B-Endorphin-Containing Memory-Cells and M-Opioid Receptors Undergo Transport to Peripheral Inflamed Tissue. J Neuroimmunol 115(1-2): 71-78.
-
Casanova ML, Blázquez C, Martínez Palacio J, Villanueva C, Fernández Aceñero MJ, et al. (2003) Inhibition of Skin Tumor Growth and Angiogenesis in Vivo by Activation of Cannabinoid Receptors. Journal of Clinical Investigation 111(1): 43-50.
-
Ibrahim MM, Porreca F, Lai J, Albrecht PJ, Rice FL, et al (2005) Cb2 Cannabinoid Receptor Activation Produces Antinociception by Stimulating Peripheral Release of Endogenous Opioids. PNAS 102(8)2: 3093-3098.
-
Tominaga M, Ogawa H, Takamori K (2007) Possible Roles of Epidermal Opioid Systems in Pruritus of Atopic Dermatitis. Journal of Investigative Dermatology 127(9): 2228-2235.
-
Slominski AT, Zmijewski MA, Zbytek B, Brozyna AA, Granese J, et al. (2011) Regulated Proenkephalin Expression in Human Skin and Cultured Skin Cells. Journal of Investigative Dermatology 131(3): 613-622.
-
Neer EJ (1995) Heterotrimeric G Proteins: Organizers of Transmembrane Signals Review. Cell 80(2): 249-257.
-
Glass M, Felder CC (1997) Concurrent Stimulation of Cannabinoid Cb1 and Dopamine D2 Receptors Augments Camp Accumulation in Striatal Neurons: Evidence for A Gs Linkage to the Cb1 Receptor. The Journal of Neuroscience 17(14): 5327-5333.
-
Bonhaus DW, Chang LK, Kwan J, Martin GR (1998) Dual Activation and Inhibition of a Denylyl Cyclase by Cannabinoid Receptor Agonists: Evidence for Agonist- Specific Trafficking of Intracellular Responses. J Pharmacol Exp Ther 287(3): 884-888.
-
Gao F, Zhang LH, Su TF, Li L, Zhou R, et al. (2016) Signaling Mechanism of Cannabinoid Receptor-2 Activation- Induced β-Endorphin Release. Mol Neurobiol 53(6): 3616-3625.
-
Bouron A (2018) Phyto and Endocannabinoids Exert Complex Actions on Calcium and Zinc Signaling in Mouse Cortical Neurons. Biochem Pharmacol 152: 244-251.
-
Ryan D, Drysdale AJ, Lafourcade C, Pertwee RG, Platt B (2009) Cannabidiol Targets Mitochondria to Regulate Intracellular Ca2+ Levels. Journal of Neuroscience 29(7): 2053-2063.
-
Mato S, Sánchez Gómez MV, Matute C (2010) Cannabidiol Induces Intracellular Calcium Elevation and Cytotoxicity in Oligodendrocytes. Glia 58(14): 1739-1747.
-
Cabot PJ, Carter L, Gaiddon C, Zhang Q, Schäfer M, et al. (1997) Immune Cell-Derived-Endorphin: Production, Release, and Control of Inflammatory Pain in Rats. J Clin Invest 100(1): 142-148.
-
Katsuyama S, Mizoguchi H, Kuwahata H, Komatsu T, Nagaoka K, et al. (2012) Involvement of Peripheral Cannabinoid and Opioid Receptors in B-Caryophyllene- Induced Antinociception. European Journal of Pain 17(5): 664-675.
- Gallic and Citric Acid Present in the Peels of Tropical Fruits as an Alternative in the Fight against Cancer
- Treating the Forehead Lines with Combination of Forehead and Glabellar Botulinum Toxin Among Japanese Patients
- Clinical Evaluation of Patients Suffering from Breast Cancer & Determination of Treatment Therapies and Better Strategies Related to Breast Cancer
- Medieval Recipes by Al-Zahrāwī for Heart Palpitations Treatment
- Etiology and Prescription Errors of Myocardial Infarction in Different Health Care Systems of Azad Kashmir
- Early Diagnosis and Multidisciplinary Management of Turner Syndrome: A Paediatric Case Study