Beneficial Effect of Ursodeoxycholic Acid on High Glucose-Induced Long QT Interval Arrhythmia in Isolated Rat Heart
The aim of the present study was to investigate whether ursodeoxycholic acid (UDCA) could prevent the high glucose-induced long QT interval arrhythmia in isolated perfused rat heart Moreover, we studied whether UDCA could affect the cardiac expression of two arrhythmias-related miRNAs, miR-1 and miR-133 together with monitoring of caspase-3, caspase-9, ubiquitin-protesome system, Mn-SOD levels and NO bioavailability. Rat hearts perfused with standard Krebs solution containing 11.1 mmol D-glucose showed a cardiac QT interval of 107±11 ms and a coronary perfusion pressure (CPP) of 72±5 mmHg. These were increased by a Krebs containing 33.3 mmol D-glucose (e.g +98% QT interval; +79% CPP). The pretreatment of the rats with UDCA (150 mg/kg in drinking water for 12 weeks) prior to the heart excision while not affecting the QT interval and CPP in rats perfused with 11.1 mmol D-glucose it significantly reduced these parameters of -46% and -43% in rat perfused with D-glucose 33.3 mmol. The beneficial effect of UDCA on the cardiac instability caused by high glucose was also observed in an in vivo setting of rats made diabetics with streptozotocin (65 mg/kg i.p.). In these rats the basal cardiac parameters R-R interval, P-R interval and QT interval were not affected by UDCA. In both the in vitro and in vivo settings the reduction of QT interval operated by UDCA was associated to a reduction of the expression of miR-1 and miR-133, a reduction of the expression of the ubiquitin, proteasome, caspase-3, and caspase-9 into the hearts and by an increase of the expression of eNOS and nitrate/nitrite levels.
Di filippo C, Trotta MC, Maisto R,Gaudino G, Accardo M, Ferraraccio F,
Berrino l, Rossi F, D'amico M*
Second University of Naples, Italy
Donatelli”, Second University of Naples, Italy, E-mail: michele.damico@unina2.it
Introduction
Bile acids are essential compounds for bile formation and cholesterol degradation; among them, ursodeoxycholic acid (UDCA) is an endogenous secondary bile acid recently implicated in the genesis of cardiac arrhythmias when present at low concentration in human serum [1]. The present study was aimed to investigate for the first time the use of UDCA as novel therapeutic option in preventing the high glucose-induced long QT interval arrhythmia and the increased coronary perfusion pressure. Cardiac arrhythmias are a variation from the normal heart rate or rhythm [2, 3, 4], due to electrical and structural remodelling mechanisms, for which hypertension, aging and diabetes mellitus are major risk factors [5]. In this last condition, hyperglycemia represents an important risk factor for ventricular instability characterized by QT interval prolongation and increased vascular tone, due to a oxidative stress response, derangement of cardiac structure and ion conductance into the heart [6, 7, 8, 9]. Despite several pharmacological approaches have been tried the hyperglycemia-induced electrical instability and long cardiac QT interval is still a clinical challenge [10] and hence the understanding of the molecular pathways governing the pathophysiology of hyperglycemia-induced long QT interval and targeting them is essential to the prevention of catastrophic cardiac complication of diabetes. In diabetes, exogenous UDCA reduces the endoplasmic reticulum stress associated with elevated glucose by inhibiting caspase activation, by down-regulating the ubiquitin-proteasome system, and inhibiting reactive oxygen species [11]. Through its binding to Farnesoid X receptor UDCA also increases the nitric oxide (NO) bioavailability in congestive heart failure [12] and may counteract the arrhythmias, that constitutes a major cause for cardiac death in this setting [4]. All together these data may account for a possible interference on hyperglycemia-induced long cardiac QT interval. Therefore, we investigated this aspect by using a combination of biochemical and functional analyses. Moreover, since recent studies have shown that some microRNAs are involved in cardiac rhythm regulation by modifying the expression levels of key ion channels, transporters, and cellular proteins that lead ventricular arrhythmias in rat myocardial infarction as for example miR-1 [13], and contributes to QT interval prolongation in diabetic rats as for example miR-133 [14] we monitored the putative heart changes of these two miRNAs following high glucose exposure and from the mechanistic point of view by monitoring the local NO bioavailability, the ubiquitin-proteasome system, the apoptotic markers, and the MnSOD.
Materials and Methods
In vitro isolated rat hearts
20 healthy male Sprague-Dawley rats purchased from Harlan (San Pietro al Natisone, Udine, Italy), of five-six weeks and weighing about 100 g, were housed in individuals cage with controlled temperature (21-23°C), lighting (12-12 h light-dark cycle) and humidity (55-60%) and were fed on a standard chow pellet diet. 10 Sprague Dawley rats were treated with 150 mg/kg UDCA dissolved in drinking water for 6 weeks [15]; of these 5 hearts were excised and were perfused with D-glucose 11.1 mmol and 5 hearts were excised and perfused with D-glucose 33.3 mmol. 10 rats were not treated with UDCA but were divided into 5 rat heart perfused with D-glucose 11.1 mmol and served as control and 5 rat perfused with D-glucose 33.3 mmol. At the end of the pretreatment period, rats were anaesthetized with ketamine hydrochloride (100 mg/kg) and metedomidine (0.25 mg/kg) and then heparanized (sodium heparin, 250 IU, i.p., 10 min before heart excision). The hearts were rapidly excised and placed in ice-cold standard Krebs solution [16] prior to coronary perfusion. Hearts were cannulated via the aorta and perfused retrogradely under constant flow (10 ml min-1) as previously described by Ceriello et al. [16]. Each experiment lasted 2 h, and all the experimental procedures were approved by Animal Care Ethical Committee of the Naples University in accordance with Italian (Decree 116/92) and European Community (E.C. L358/1 18/12/86) guidelines on the use and protection of laboratory animals.
ECG and CPP measurements
For each heart, a unipolar electrocardiogram (ECG) was recorded by implanting a stainless steel wire electrode into the apex of the left ventricular muscle mass, with a second electrode connected to the aorta. A 3-min ECG (speed 50 mm/s) was recorded every 10 min for 2 h and the following parameters were calculated: (i) heart rate (R–R interval), (ii) the duration between onset of the P wave and onset of the ventricular complex (PR interval) and (iii) the width of the ventricular complex, measured at 100% repolarization and defined as QT. The QT value of each experiment was calculated as the mean of the ECG readings performed every 10 min. The coronary perfusion pressure (CPP) was monitored continuously during each study with a Statham
Spectramed pressure transducer connected to a chart recorder (Grass, 79E, Quincy, MA, USA). Air temperature was maintained by means of a heated (37 °C) water jacket. On establishing a stable CPP (20–30 min following cannulation), experiments were performed as previously described. CPP values used for statistical comparisons were calculated either as the mean of each 10-min value throughout the entire experiment or as the mean of the steady-state increase above baseline, if an increase in CPP was evident during an experiment.
Hematoxylin/eosin staining of the cardiac sections
At the end of each experiment the hearts were cut in two halves from the base to the apex and half was immediately frozen in liquid nitrogen for RNA extraction, WB and ELISA assay while the other half was paraffin embebbed and cut in 5µm section for the staining with hematoxylin/eosin.
Tissue nitrate/nitrite levels
To determinate NO bioavailability in isolated rat hearts, a nitrate/nitrite colorimetric assay Kit (Cayman Chemical, MI, USA) was used according to the manufacturer’s protocol.
Western Blotting Assay
After tissue homogenization in a solution of 0.5% hexadecyl- trimethyl-ammonium bromide dissolved in 10mM potassium phosphate buffer (pH 7) and centrifugation for 30 min at 4,000×g at 4°C, tissues protein concentration was determined using the Bio-Rad protein assay (Bio-Rad Laboratories, Milan, Italy). After the gel electrophoresis in a 8% PAGE separation gel, the protein sample were electro- transferred onto a PVDF membrane. Blots were blocked with 5% non-fat dry milk for 1h at room temperature, and then incubated with primary specific antibodies over-night, followed by incubation with a horseradish peroxidase-conjugated secondary antibody for 1 h at room temperature. The signal was expressed as densitometric units (DU).Western blots were performed to evaluate the expression levels of caspase-3, caspase-9, Ubiquitin (Ub) and Proteasome System (20S and 26S) by means of the following primary antibodies: anti-Caspase 3 (AB3623, Millipore, Milan, Italy), anti-Caspase-9 (ab32539, Abcam, USA), anti-Ub (sc- 9133, SantaCruz Biotec, USA), anti-20S Proteasome (sc- 58412, SantaCruz Biotec, USA), anti-26S Proteasome (sc- 58311, SantaCruz Biotec, USA) and anti-eNOS (NB300- 500, Novus Biologicals, Abingdon, UK). For all assays goat anti-rabbit (sc-2004) and anti-mouse (sc-2005) HRP horseradish peroxidase were used as secondary antibodies (SantaCruz Biotec, USA).
RNA extraction and miRNA profiling
Total RNA, including small RNAs, was extracted from 50 mg of heart tissue collected at the end of the experimental procedure using the miRNeasy Mini kit (Qiagen) according to the supplementary protocol Purification of Total RNA, including Small RNAs, from Animal Cells. In order to monitor the efficiency of miRNA recovery and to normalize miRNA expression in the subsequent Real-time PCR experiment, a 5µL aliquot of 5nM Syn-cel-miRNA-39 miScript miRNA Mimic was spiked into each sample before nucleic acid preparation. RNA was then quantificated using NanoDrop 2000c Spectophotometer (Thermo Fisher Scientific, Waltham, MA USA). Total RNA was then reverse-transcribed using the miScript II RT kit (SABiosciences) according the manifacturer’s protocol: cDNA synthesis was obteined adding 2 µg of total RNA to a mix reaction of 2 µl of 10x miScript Nucleics Mix, 2 µl of miScript Reverse Transcriptase Mix, 4 µl of 5x miScriptHiSpec Buffer and Rnase-free water for a final volume reaction of 20 µl. The samples were incubated at 37°C for 1 hour, 95°C for 5 minutes and kept at 4°C in the Mastercycler personal (Eppendorf) to synthetize the cDNAs. The quantization and normalization of mature miR-1 and miR-133 have been performed using Syn-cel-miR-39-3p both as a control for the extraction step and as a normalizer gene for the phase of Real-Time PCR. For a single PCR reaction we used 12.5µl of 2x QuantiTect SYBR Green PCR Master Mix (QIAGEN), 2.5µl of template cDNA (10 times diluted), 2.5µl of 10x miScript Primer Assays (QIAGEN), containing a primer for a specific miRNA (miR-1, miR-133 and Syn- cel-miR-39-3p), and RNAse free water (QIAGEN) to a final volume of 25µl. Each reaction was carried out in triplicate in Bio-rad CFX96 cycler (Bio-Rad Laboratories, Inc). Cycling conditions were as follows: 95°C for 15 min; 94°C for 15 sec; 58°C for 30 sec, 70°C for 30 sec, repeated for 40 cycles; melt curve: 55-95°C. Relative quantization of gene expression was performed calculating the DCt value for each miRNA as Ct miRNA – Ct Syn-cel-miR39; expression fold change was then obtained as 2^-DCt. The P-values are calculated based on a Student’s t-test of the replicate 2^-DCt values for each miRNA in the control and pre-treated groups. P<0.05 was considered significant.
Statistical Analysis
Data values are expressed as mean ± SEM of n number of rats for the in vivo experiments. Coronary perfusion pressure and ECG values used for statistical comparisons are expressed either as the mean of each 3-min value throughout the entire experiment or as the mean of the steady-state increment above baseline, if an increase in coronary perfusion pressure or modifications of ECG parameters were evident during an experiment. Statistical analysis was assessed either by Student’s t-test (when only two groups were compared) or one-way ANOVA followed by Dunnett’s test (more than two experimental groups). A probability P value less than 0.05 was considered significant to reject the null hypothesis.
In vivo proof of concept
To confirm the role of UDCA in reducing high glucose- induced QT interval, we translated the in vitro experiments in a setting of in vivo ones. We investigated the effects of UDCA in diabetic Sprague-Dawley rats using a method already described [8] with some modifications. A first group of rats (n=5) received drinking water only. Other two groups (n=5 each) were treated with 150 mg/kg UDCA dissolved in drinking water for 6 weeks and one of these (STZ+UDCA) was administered at the end by a single injection of 65 mg/kg i.p. streptozotocin (Sigma Chemical Co., St. Louis, MO, USA) dissolved in 0.1 M citrate buffer (pH 4.3) associated to a daily subcutaneous administration of regular insulin (1.5 ± 0.5 units/day) applied in order to render it diabetic and yield blood glucose levels of ∼22 mmol/l (350 mg/dl) for 8 days (poor glycemic control rats). A fourth group (STZ) did not receive UDCA and was administered with streptozotocin+insulin only, serving as control for the group STZ+UDCA. At the end of the treatmens rats with plasma glucose levels above 300 mg/dl were included in the study and monitored for a 3 min lead II ECG at 50 mm/sec speed (Cardioline, Hellige, Germany) according to Di Filippo et al. [17]. The time points considered were 0-2-6-8 days from the STZ injection and i) heart rate (R-R interval); ii) atrio-ventricular conduction time (P-R interval); and iii) width of the ventricular complex (QT interval) measured as the average (ms±s.e.m.) of at least three measurements during the 3 min continuous ECG. The ECG analysis was carried out by a cardiologist blinded of the animal group. From the molecular point of view, the cardiac expression levels of miR-1 and miR-133 were monitored at day 8, in order to mirror the isolated heart.
Results
The perfusion of the rat hearts with a Krebs containing a high glucose concentration (33.3 mmol) caused appearance of several necrotic zones within the cardiac myocytes caused by local coagulation, and genesis of interstitial edema as observed after staining of the heart section with hematoxylin/eosin with respect to the heart perfused with glucose 11.1 mmol concentration (Figure 1). These signs were reduced in the hearts treated with UDCA 150 mg/kg in drinking water for 12 weeks harvested and perfused with the high glucose. Indeed, there was a reduced edema and reduced linfo-monocyte phlogosis accompanied by some regenerative myocytes aspects (Figure 1).
QT interval and CPP measurement
Rat hearts perfused with standard Krebs solution containing D-glucose 11.1 mmol (Normo) showed a cardiac QT interval of 107±11 ms and a coronary perfusion pressure of 72±5 mmHg. As shown in (Table 1), the perfusion of the hearts with a Krebs containing a high glucose concentration (High, 33.3 mmol) registered high values of QT interval (+98% vs Normo) and CPP (+79% vs Normo). The pretreatment of the rats with UDCA (150 mg/kg in drinking water for 12 weeks) prior to the heart excision, while not affecting the QT interval and CPP in rats perfused wih D-glucose 11.1 mmol (Normo),
| TREATMENT | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| High+Ud | Normo+ | |||||||||||||
| Normo | High | |||||||||||||
| ca | Udca | |||||||||||||
| QT interval (ms±s.e.m.) | 107 ± 10 | 172 ± 8** | 115 ± 12°° | 105 ± 7 | ||||||||||
| CPP (mmHg±s.e. m.) | 72 ± 5 | 129 ± 7** | 74 ± 6° | 71 ± 8 |
Table 1: In vitro effects of UDCA on QT interval and CPP in isolated heart. 20 rat hearts (n=5 each group) were perfused with 11.
Table 1: In vitro effects of UDCA on QT interval and CPP in isolated heart. 20 rat hearts (n=5 each group) were perfused with 11.1 mmol D-glucose (NORMO), 33.3 mmol D-glucose (HIGH) and pre-treated or not with ursodeoxycholic acid (UDCA, 150 mg/kg) in drinking water for 12 weeks. QT=cardiac QT interval; CPP=coronary perfusion pressure. Significant differences are indicated as **P<0.01 vs NORMO; °P<0.05 vs HIGH; °°P <0.01 vs HIGH.
A B
C
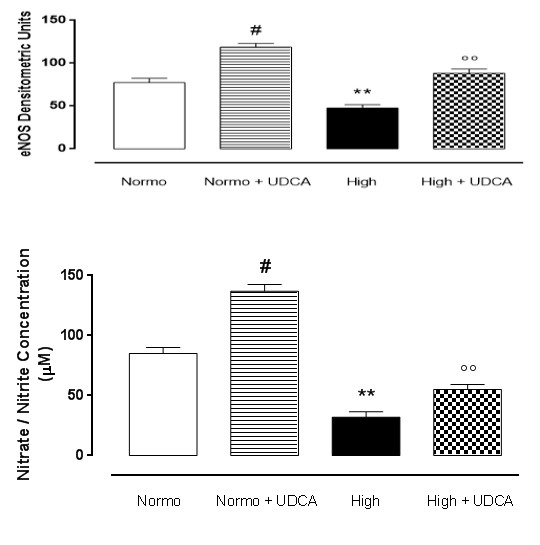
UDCA and eNOS expression into the heart
Perfusion of the rat hearts with a Krebs solution containing 11.1 mmol D-glucose showed high levels of Mn-SOD, which were sensibly decreased in hearts perfused with 33.3 mmol D-glucose (Figure 2). Pre- treatment of the rats with 150 mg/kg UDCA significantly reported the levels of Mn-SOD recorded in the hearts perfused with 33.3 D-glucose towards the values of isolated hearts perfused with standard Krebs (11.1 mmol D-glucose), almost recovering the expression of this enzyme. Interestingly, the hearts collected from rats treated with UDCA alone and perfused with normal glucose showed significantly (p<0.05) higher levels of Mn- SOD with respect to the levels observed in the heart without treatment and perfused with normal Krebs (Figure 2A,B).
Tissue nitrate/nitrite levels
As shown in Figure 2C, nitrate/nitrite concentration was lower in hearts perfused with high glucose Krebs compared to hearts perfused with standard normoglycemic Krebs, as expected. Interestingly, UDCA pretreatment significantly increased the tissue nitrate/nitrite levels in heart perfused with D-glucose 33.3 mmol, showing a higher NO bioavailability compared to the hearts perfused with high glucose Krebs only. The increase of nitrate/nitrite levels caused by UDCA was also evident in the hearts collected from rats treated with UDCA alone and perfused with normal glucose with respect to the levels observed in the heart without treatment and perfused with normal Krebs (Figure 2C).
$$ - - - - $$
e-NOS $$ - - - - $$
B-actin
Normo Normo + UDCA
High High + UDCA
Rat hearts (n=5 each group) were perfused with 11.1 mmol D-glucose (Normo), 33.3 mmol D-glucose (High) and pre-treated or not with ursodeoxycholic acid (UDCA, 150 mg/kg) in drinking water for 6 weeks. The values are expressed as the mean±s.e.m. Significant differences are indicated as **P<0.01 vs Normo; °°P <0.01 vs High; #P<0.01 vs Normo.
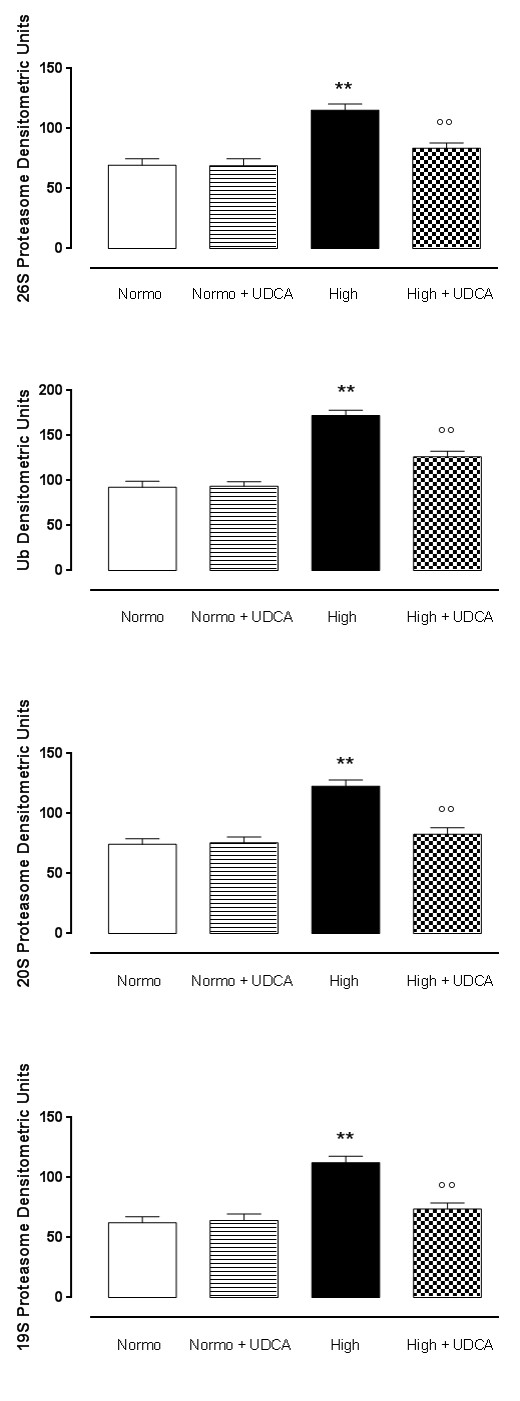
UDCA and ubiquitin proteasome system (UPS) expression The expression levels of UPS subunits were found to be significantly (p<0.01) augmented in isolated rat hearts A D
B
C
Figure 3: UDCA and ubiquitin proteasome system (UPS) expression. Depicted are the graphs concerning the levels, in densitometric units, of ubiquitin (panel A), 20S protesome (panel B), 19S proteasome (panel C), and 26S proteasome (panel D) subunits in rat hearts perfused with 11.1 mmol D-glucose (Normo), 33.3 mmol D-glucose (High) and pre- treated or not with ursodeoxycholic acid (UDCA, 150 mg/kg) in drinking water for 6 weeks. (E) Representative perfused with high glucose Krebs (Figure 3A-3E). UDCA pretreatment (150 mg/kg) resulted in a significant (p <0.1) decrease of the expression levels of all the UPS subunits Ubiquitin, 19S proteasome, 20S proteasome and 26S proteasome (Figure 3A-3E). The hearts collected from rats treated with UDCA alone and perfused with normal glucose show normal expression of UPS subunits if compared to the expression levels observed in the heart without treatment and perfused with normal Krebs (Figure 3A-3E).
E
$$ - - - - $$
Ub
$$ - - - - $$
20S $$ - - - - $$
19S $$ - - - - $$
26S $$ \therefore \mathrm {一} \mathrm {一} \mathrm {一} \mathrm {一} $$
B-actin
Normo Normo + UDCA
High High + UDCA
western blotting traces for these markers. The values are expressed as the mean±s.e.m. of n=5 hearts per group. Significance is expressed as **P < 0.01 vs Normo; °P < 0.05 vs High; °°P < 0.01 vs High.
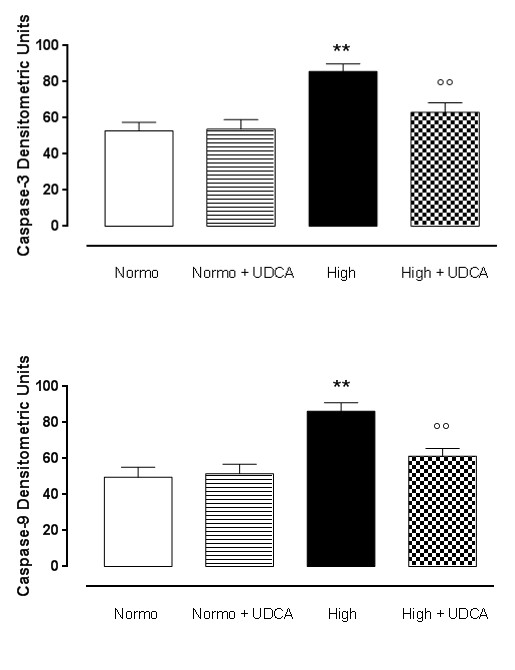
Effect of UDCA pre-treatment on apoptosis
High glucose conditions (33.3mmol) led to increased apoptotic phenomena into the heart, consisting in a higher expression of caspase-3 and caspase-9 compared to isolated rat hearts perfused with standard Krebs solution (Figure 4A-4C). These caspases were decreased (p<0.01) after treatment of the rat with 150 mg/kg UDCA A in drinking water for 12 weeks (Figure 4A-4C). The hearts collected from rats treated with UDCA alone and perfused with normal glucose did not show any sign of apoptosis with levels of caspase-3 and caspase-9 similar to those observed in the heart without treatment and perfused with normal Krebs (Figure 4A-4C).
B
$$ - - - - $$
Caspase-9
Figure 4: UDCA reduces the apoptosis caused by high glucose into the rat hearts. Panels A-C, western blottings and relative densitometric units of Caspase, Caspase-9 in rat hearts perfused with 11.1 mmol D-glucose (Normo), 33.3 mmol D-glucose (High) and pre-treated or not with ursodeoxycholic acid (UDCA, 150 mg/kg). Depicted are the graphs, in densitometric units expressed as the mean±s.e.m. of n=5 hearts per group. Significance is expressed as **P < 0.01 vs Normo; °P < 0.05 vs High; °°P < 0.01 vs High.
UDCA pre-treatment and QT-related cardiac miRNA
qRT-PCR showed that following perfusion of the hearts with high glucose (33.3 mmol) Krebs there was a significant (p<0.01) up-regulation of the expression of miR-1 and miR-133 compared to isolated hearts perfused with normal glucose (Normo, 11.1 mmol) Krebs (Figure 5). In contrast to this, hearts excised from rats pre-treated with High+UDCA (150 mg/kg) showed significant reduction of the up-regulation of miR-1 and miR-133 with levels addressed toward the values of heart perfused with D-glucose 11.1 mmol (Normo, Figure 5). The hearts collected from rats treated with UDCA alone and perfused with normal glucose (Normo+UDCA) did not produce any change of these miRNAs with respect to the expression levels observed in the heart without treatment, and perfused with normal Krebs (Figure 5).
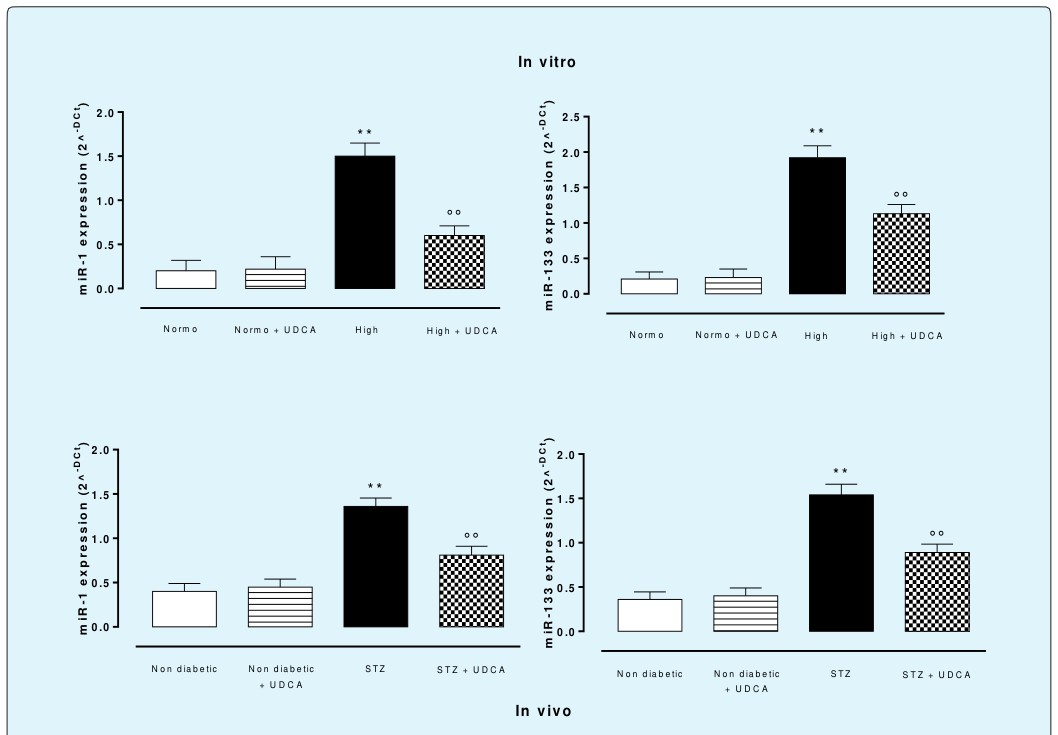
Figure 5: Effects of UDCA on cardiac miR-1 and miR-133 expression levels. (In vitro), rat hearts (n=5 each group) were perfused with 11.1 mmol D-glucose (Normo), 33.3 mmol D-glucose (High) and pre-treated or not with ursodeoxycholic acid (UDCA, 150 mg/kg) in drinking water for 6 weeks. (In vivo), non diabetic rats, non diabetic+UDCA rats, STZ- diabetic rats and STZ-diabetic+UDCA rats were used. 150 mg/kg UDCA, dissolved in drinking water, was administered for 6 weeks before the induction of 8 days diabetes with a single injection of STZ (65 mg/kg i.p.). miRNAs levels are expressed as 2^-Dct values ± s.e.m. of n=5 hearts per group. Significant differences are indicated as **P<0.01 vs Normo; °°P <0.01 vs High.
In vivo setting
The treatment of Sprague-Dawley rats with UDCA for 6 weeks did not affect per sé the body weight of the rats, the plasma glucose levels, and the insulin levels recorded in control rats (Table 2). Similarly, the basal cardiac parameters, R-R interval, P-R interval and QT interval were not affected by UDCA. 8 days of poor glycemic control in diabetic rats caused a hyperglycemia of 22±0.4 mmol/l with reduction of R-R interval from 154±9 ms to 110±12 ms, no changes of P-R interval (hyperglycemia 58±7 ms; normoglycemia 61±4 ms), and a significant prolongation (+77% at day 8) of cardiac QT interval compared with non diabetic normoglycemic rats (Table 3). UDCA markedly reduces the QT interval in diabetic rats during the 8 days of poor glycemic control (Table 3), having a maximum reduction of -42.5% at day 8. The effects of UDCA observed on isolated heart for that concerning the miRNAs expression was copied in the vivo STZ-diabetic setting after 8 days of poor glycemic control. In this case 6 weeek treatment with UDCA (150 mg/kg) before the induction of diabetes prevents both the upregulation of miR-1 and miR-133 (Figure 5).
| Parameters | Groups | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Drinking | ||||||||||||||
| Drinking | Drinking | Drinking | ||||||||||||
| water | ||||||||||||||
| water+UDCA | water+STZ | water+STZ+UDCA | ||||||||||||
| (control) | ||||||||||||||
| Δ Body weight | 89±7 | 91.3±5.5 | 104±2 | 90±7 | ||||||||||
| (gr) | ||||||||||||||
| Glycemia | 5.0±0.1 | 5.6±0.15 | 22.7±0.1* | 15.6±0.15° | ||||||||||
| (mmol/l±s.e.m.) | ||||||||||||||
| Insulin | 9.1±0.8 | 9.8±0.5 | 30.5±1.7* | 10.6±1.2° | ||||||||||
| (µU/ml±s.e.m.) | ||||||||||||||
| HOMA-IR | 2.02±0.08 | 2.43±0.9 | 30.8±2.5* | 7.3±1.3° | ||||||||||
| Sodium | 129±5 | 130±7 | 131±3 | 133±5 | ||||||||||
| (mmol/l±s.e.m.) | ||||||||||||||
| Potassium | 4.6±0.7 | 4.4±0.5 | 4.8±0.9 | 4.3±0.4 | ||||||||||
| (mmol/l±s.e.m.) | ||||||||||||||
| MABP | 106±4 | 104±5 | 118±5 | 104±9 | ||||||||||
| (mmHg±s.e.m.) | ||||||||||||||
| HR (beats/min- | 379±13 | 387±10 | 369±15 | 377±8 | ||||||||||
| 1) | ||||||||||||||
| (R-R interval, | 154±9 | 159±9 | 156±5 | 154±3 | ||||||||||
| ms±s.e.m.) | ||||||||||||||
| P-R interval | 59±3 | 56±2.5 | 57±4 | 56±3.1 | ||||||||||
| (ms±s.e.m.) |
Table 2: In vivo effect of ursodeoxycholic acid on plasma and systemic parameters of non diabetic and diabetic rats after 6 weeks
Table 2: In vivo effect of ursodeoxycholic acid on plasma and systemic parameters of non diabetic and diabetic rats after 6 weeks of treatment. Δ=increase in body weight (gr) after 12 weeks of treatments; UDCA= ursodeoxycholic acid (150 mg/kg ad libitum); MABP=mean arterial blood pressure; HR= Heart rate. Values are expressed as mean ± SEM (n = 5 rats per group). *p<0.01 vs drinking water (control); °p<0.01 vs drinking water+STZ.
Discussion
The bile acid UDCA, already approved by the Food and Drug Administration for the clinical pretreatment of cholestatic liver diseases [11], has recently inspired much interest because of his cytoprotective activity and potential therapeutic effects on several cardiovascular and non-liver diseases both in experimental and clinical settings [18, 19, 20, 21, 22]. However, despite the pioneering studies of Lee et al., [18] on the heart dynamics (e.g. heart rate, left ventricular developed pressure, coronary flow) the cardiac aspects of UDCA have not been further and deeply investigated. As for example, there is a scant knowledge on the effects of UDCA on the cardiac electrical instability caused by a metabolic dysfunction as it is diabetes. Diabetes is a condition that several researchers, including ourselves, have demonstrated be negative for the cardiac electrical stability mainly because of the high levels of glucose into the heart tissue responsible of a high stress to the heart cells and derangement of their function from the normal electrical conduction, thus causing arrhythmias [9]. The QT interval prolongation arrhythmia and increased vasomotor tone are the outcomes [9]. In light of the incomplete and still ongoing knowledge of the pathogenetic molecular mechanism underlying the biology of UDCA, the lack of structural, functional and biochemical studies in human subjects, and the need of new local therapeutic options for long cardiac QT interval [8], we show here that UDCA prevents the cardiac QT interval prolongation and the increased vasomotor tone caused by high glucose in isolated perfused rat hearts. This was accompanied by increased cardiac eNOS enzyme and nitrate/nitrite levels, by reduced ubiquitin- proteasome subunits, caspase-3 and caspase-9 into rat heart tissue. We propose that the increased NO bioavailability from UCDA together with the specific deactivation of inflammatory, oxidative and apoptotic signaling may be responsible of this. Interestingly enough for this point, Liu et al. [12] reported that UDCA could be considered a new therapeutic agent for treating reduced nitric oxide bioavailability in congestive heart failure and other cardiovascular diseases as a ligand for Farnesoid X receptor (FXR) present in cardiomyocytes and endothelial cells [23]. Therefore, UDCA may represent an alternative approach for the treatment of high glucose-induced long cardiac QT interval because it improves cardiac electrical performance compromised by high glucose. This latter effect also underlined by changes in the expression of some miRNAs related to cardiac electrical conduction and stability. These were miR-1 and miR-133, the most abundant miRNAs expressed in the heart [24, 25] whose targets include mRNA transcripts encoding proteins involved in cardiovascular development, hypertrophy and ion channel function [26, 27, 28, 29]. In our studies, UDCA significantly reduced the up-regulated miRNAs in parallel with reduced cardiac QT interval and reduced vascular parameters. Noteworthy, increased levels of miR-1 and miR-133a are related to QT interval prolongation in different experimental models [29, 30, 31, 32, 33, 34]: up-regulated miR- 1 leads to ventricular electrical derangement in rat myocardial infarction [14], while in diabetic rats up- regulated miR-133 contributes to QT interval prolongation [15].
Conclusion
In conclusion, thanks to its anti-apoptotic, anti- inflammatory properties and its role in maintaining cardiovascular NO bioavailability associated to a regulation of miRNAs expression, UDCA could represent a valid drug in the prevention of cardiac consequences of high glucose levels into the heart, and hyperglycemia in diabetes.
References
-
Rainer P, Primessnig U, Harenkamp S, Doleschal B, Wallner M, et al. (2013) Bile acids induce arrhythmias in human atrial myocardium: implications for altered serum bile acid composition in patients with atrial fibrillation. Heart 99(22): 1685-1692.
-
Kim GH (2013) MicroRNA Regulation of Cardiac Conduction and Arrhythmias. Transl Res 161(15): 381-392.
-
Kumar KR, Mandleywala SN, Link MS (2015a) Atrial and Ventricular Arrhythmias in Hypertrophic Cardiomyopathy. Card Electrophysiol Clin 7(2): 173- 186.
-
Kumar S, Stevenson WG, John RM (2015b) Arrhythmias in Dilated Cardiomyopathy. Card Electrophysiol Clin 7(2): 221-233.
-
Schoonderwoerd BA, Van Gelder IC, Van Veldhuisen DJ, Van den Berg MP, Crijns HJ (2005) Electrical and structural remodeling: role in the genesis and maintenance of atrial fibrillation. Prog Cardiovasc Dis 48(3): 153-168.
-
Kuusisto J, Mikkanen L, Pyorala K, Laakso M (1994) NIDDM and its metabolic control predict coronary artery disease in elderly people. Diabetes 43(8): 960- 967.
-
Capes SE, Hunt D, Malmberg GK, Gerstein HC (2000) Stress hyperglycaemia and increased risk of death after myocardial infarction in patients with and without diabetes: a systematic overview. Lancet 2000; 355(9206): 773-778.
-
D'Amico M, Marfella R, Nappo F, Di Filippo C, De Angelis L, et al. (2001) High glucose induces ventricular instability and increases vasomotor tone in rats. Diabetologia 44(4): 464-470.
-
Di Filippo C, Cuzzocrea S, Marfella R, Fabbroni V, Scollo G, et al. (2004) M40403 prevents myocardial injury induced by acute hyperglycaemia in perfused rat heart. European Journal of Pharmacology 497(1): 65-74.
-
Barrett T (2015) Why Are We Prolonging QT Interval Monitoring? Dimens Crit Care Nurs 34(3): 130-133.
-
Vang S, Longley K, Steer CJ, Low WC (2014) The Unexpected Uses of Urso- and Tauroursodeoxycholic Acid in the Treatment of Non-liver Diseases. Glob Adv Health Med 3(3): 58-69.
-
Liu X, Fassett J, Wei Y, Chen Y (2013) Regulation of DDAH1 as a Potential Therapeutic Target for Treating Cardiovascular Diseases. Evidence-Based Complementary and Alternative Medicine 2013: 619207.
-
Lu Y, Zhang Y, Shan H, Pan Z, Li X, et al. (2009) MicroRNA-1 downregulation by propranolol in a rat model of myocardial infarction: a new mechanism for ischaemic cardioprotection. Cardiovascular Research 84(3): 434-441.
-
Xiao J, Luo X, Lin H, Zhang Y, Lu Y, et al. (2007) MicroRNA miR-133 Represses HERG K Channel Expression Contributing to QT Prolongation in Diabetic Hearts. Journal of Biological Chemistry 282(17): 12363-12367.
-
Mahmoud AA, Elshazly SM (2014) Ursodeoxycholic acid ameliorates fructose-induced metabolic syndrome in rats. PLoS One 9(9): e106993.
-
Ceriello A, Quagliaro L, D'Amico M, Di Filippo C, Marfella R, et al. (2002) Acute hyperglycemia induces nitrotyrosine formation and apoptosis in perfused heart from rat. Diabetes 51(4): 1076-1082.
-
Di Filippo C, Marfella R, Cuzzocrea S, Piegari E, Petronella P, et al. (2005) Hyperglycemia in streptozotocin-induced diabetic rate increases infarct size associated with low levels of myocardial HO-1 during ischemia/reperfusion. Diabetes 54(3): 803- 810.
-
Lee WY, Han SH, Cho TS, Yoo YH, Lee SM (1999) Effect of ursodeoxycholic acid on ischemia/reperfusion injury in isolated rat heart. Arch Pharm Res 22(5): 479-484.
-
Suh H, Jung EJ, Kim TH, Lee HY, Park YH, et al. (1997) Anti-angiogenic activity of ursodeoxycholic acid and its derivatives. Cancer Lett 113(1-2): 117-122.
-
Ozel Coskun BD, Yucesoy M, Gursoy S, Baskol M, Yurci A, et al. (2015) Effects of ursodeoxycholic acid therapy on carotid intima media thickness, apolipoprotein A1, apolipoprotein B, and apolipoprotein B/A1 ratio in nonalcoholic steatohepatitis. Eur J Gastroenterol Hepatol 27(2): 142-149
-
von Haehling S, Schefold JC, Jankowska EA, Springer J, Vazir A, et al. (2012) Ursodeoxycholic acid in patients with chronic heart failure: a double-blind, randomized, placebo-controlled, crossover trial. J Am Coll Cardiol 59(6): 585-592.
-
Sinisalo J, Vanhanen H, Pajunen P, Vapaatalo H, Nieminen MS (1999) Ursodeoxycholic acid and endothelial-dependent, nitric oxide-independent vasodilatation of forearm resistance arteries in patients with coronary heart disease. Br J Clin Pharmacol 47(6): 661-665.
-
Zhang L, Li T, Yu D, Forman BM, Huang W (2012) FXR protects lung from lipopolysaccharide-induced acute injury. Molecular Endocrinology 26(1): 27-36.
-
Liang Y, Ridzon D, Wong L, Chen C (2007) Characterization of microRNA expression profiles in normal human tissues. BMC Genomics 8:166.
-
Luo X, Zhang H, Xiao J, Wang Z (2010) Regulation of human cardiac ion channel genes by microRNAs: theoretical perspective and pathophysiological implications. Cell Physiol Biochem 25(6): 571-586.
-
Care A, Catalucci D, Felicetti F, Bonci D, Addario A, et al. (2007) MicroRNA-133 controls cardiac hypertrophy. Nat Med 13(5): 613–618.
-
Ikeda S, He A, Kong SW, Lu J, Bejar R, et al. (2009) MicroRNA-1 negatively regulates expression of the hypertrophy-associated calmodulin and Mef2a genes. Mol Cell Biol 29(8): 2193-2204.
-
Liu N, Olson EN (2010) MicroRNA regulatory networks in cardiovascular development. Dev Cell 18(4): 510-525.
-
Matkovich SJ, Wang W, Tu Y, Eschenbacher WH, Dorn LE, et al. (2010) MicroRNA-133a protects against myocardial fibrosis and modulates electrical repolarization without affecting hypertrophy in pressure-overloaded adult hearts. Circ Res 106(1): 166-175.
-
Shan H, Zhang Y, Lu Y, Zhang Y, Pan Z, et al. (2009) Downregulation of miR-133 and miR-590 contributes to nicotine-induced atrial remodelling in canines. Cardiovasc Res 83(3): 465-472.
-
Hedley PL, Carlsen AL, Christiansen KM, Kanters JK, Behr ER, et al. (2014) MicroRNAs in cardiac arrhythmia: DNA sequence variation of MiR-1 and MiR-133A in long QT syndrome. Scand J Clin Lab Invest 74(6): 485-491.
-
Amaral JD, Viana RJ, Ramalho RM, Steer CJ, Rodrigues CM (2009) Bile acids: regulation of apoptosis by ursodeoxycholic acid. J Lipid Res 50(9): 1721-1734.
-
Ramalho RM, Viana RJ, Low WC, Steer CJ, Rodrigues CM (2008) Bile acids and apoptosis modulation: An emerging role in experimental Alzheimer’s disease. Trends Mol Med 14(2): 54-62.
-
Parry GJ, Rodrigues CM, Aranha MM, Hilbert SJ, Davey C, et al. (2010) Safety, tolerability and cerebrospinal fluid penetration of ursodeoxycholic Acid in patients with amyotrophic lateral sclerosis. Clin Neuropharmacol 33(1): 17-21.
- Investigation of Polymorphisms in PPAR-Ɣ and TRHR Genes and their Impact on Turkish Diabetic and Obese Individuals
- The Impact of Aircraft Noise Exposure on the Efficacy of Empagliflozin Therapy in an Animal Model of Obesity
- Rooibos Mitigates Metabolic and Inflammatory Dysfunctions in Mice Fed a High-Carbohydrate Diet
- Synergistic Effect of Combined Leaf Extract of Vernonia amygdalina, Ocimum gratissimum, and Zingiber officinale Tuber on Phytochemical Profile, Antioxidant Activity, Serum Insulin, and Biochemical Parameters in Streptozotocin-Induced Diabetic Rats
- Investigation of Cardiovascular Responses to Aerobic Exercise in Obese University Students
- A Look at the Phase Angle Obtained by Electrical Bioimpedance