Tomato: A Model Plant to Study Plant-Pathogen Interactions
Tomato (Solanum lycopersicum) is a very important vegetable plant in the worldwide because of its importance as food, quality of fruit, improves productivity, and resistance to biotic and abiotic stresses. Tomato has been extensively used not just for food however conjointly as a research (plant-pathogen interactions) material. Generally, most of the tomato traits are agronomically imperative and cannot be studied using other model plant systems. It belongs to family Solanaceae and intimately associated with several commercially important plants like potato, tobacco, peppers, eggplant, and petunias. Production of tomato yield is affected each year due to range of pathogenic diseases that square measure caused by fungi, bacteria, viruses and roundworm, enlarge all the methods through soil-borne, above-ground infections and in some instances are transmitted through insect feeding. This review is focused on the way to tomato-pathogen interactions analysis is very important and role of pathological processes connected factors and genes.
Introduction
Tomato (Solanum lycopersicum) is a member of the family Solanaceae. This is a small genus of annual or short lived perernnial herbs, indigenous to the western regions of tropical South America. One species Solanum lycopersicum is widely cultivated throughout the world. In India, tomatoes can be grown nearly throughout the year. The tomato is currently grownup worldwide for its edible fruits, with thousands of cultivars having been chosen with variable fruit varieties and for optimum growth in differing growing conditions. The tomato plant requires a warm growing season with plenty of sunshine and adequate moisture. It does not tolerate frost. It can be cultivated under irrigation in arid tropics, but hot and dry or hot and humid months do not favour its growth. High humidity with high temperature renders its susceptibility to foliage diseases. For the proper development of colour in the fruit, warm sunny days and moderately cool nights are necessary. As in most sectors of agriculture, there is increasing demand in developed and/or developing country for tomato. The chemical composition of tomato varies with variety and stage of harvest. In 2012, FAOSTAT reported that India is the second largest country for tomato production (17,500,000 MT) while China is the largest producer (50,000,000 MT). Because of its importance as food, tomato has been bred to enhance productivity, fruit quality and resistance to organic phenomenon and abiotic stresses. Carotenoids, β- carotene and lycopene constitute the chief colouring matter of tomato; their concentration in the fruit varies widely with varieties and the stages of ripeness.
Tomato (Lycopersicon esculentum Mill.) has provided excellent model systems to study plant-pathogen interactions Meissner, et al. [1], Emmanuel & Levy [2], Meena, et al. 2017 [3, 4] and its prospects for the future research are promising. An international consortium named International Solanaceae Genomics Project (SOL) is currently proceeding with whole genome sequencing of tomato. In order to be relevant in the post-genomic era, the accumulation of information on tomato-pathogen interactions are important. Tomato is an important vegetable crop plant grown worldwide for its edible fruits. Several varieties of tomatoes are available worldwide producing optimum growth and yield in different growth conditions. However, its cultivation has been limited by an abundance of diseases caused by fungi, bacteria, viruses and nematodes. In order to establish effective control methods to control them, the analysis of tomato-pathogen interactions is important. Jones, et al. [5] described the major diseases of tomato caused by 24 fungi, 7 bacteria, 10 viruses, 3 viroids and multiple nematodes. This large diversity of pathogens emphasizes the importance of the tomato-pathosystem as a favorable model for studying plant-pathogen interactions. There has also been substantial investigation of interactions between tomato and its pathogens, such as Fusarium oxysporum, Alternaria alternata, Pseudomonas syringae and Cucumber mosaic virus (CMV) [6]. The work on tomato research has recently been advanced by Kazusa DNA Research Institute (Kisarazu, Chiba, Japan), which is proceeding with whole genome sequencing of tomato cv. Moneymaker.
It is one of the important "protective foods" both because of its special nutritive value and widespread production. It is the world's largest vegetable crop afterward potato and sweet potato, however it first-rate the list of canned vegetables [7]. Current progress on the tomato genome sequencing project has generated helpful information. Additionally, the tomato belongs to the very massive potato family and is closely associated with several commercially vital plants like potato, eggplant, peppers, tobacco and petunias. Information obtained from studies conducted on tomato is simply applied to those plants that make tomato significant research material. Owing to these facts, tomato is a model organism for the Solanaceae family and specifically, for fleshy-fruited plants.
Tomatoes are rich in vitamin A and C and are naturally low in calories. They are also an excellent source of lycopene, which has been shown to protect oxidative damage in many epidemiological and experimental studies. In addition to its antioxidant activity, different metabolic effects of carotenoid (lycopene) have also conjointly been incontestable. Lycopene is that the richest supply within the diet tomato and tomato-derived merchandises. Tomatoes are often grown in greenhouse in cooler climates. Hydroponic tomatoes are offered and therefore the technique is commonly utilized in hostile growing environments, yet as high-density plantings. Apart from being used as a source of fruit and vegetable, the tomato plant is also an excellent model plant to study plant-microbe interaction. Tomato cultivars vary widely in their resistance to diseases.
Being one of the most popular vegetable throughout the world, the importance of its cultivation is threatened by a wide array of pathogens (fungi, bacteria, viruses and nematodes). This diversity of the pathogens emphasizes the importance of the tomato pathosystem as a favourable model for studying plant-pathogen interactions. Moreover, tomato carries several specific resistance (R) genes against a variety of pathogens, which make this plant suitable for genetic studies of plant host-specific resistance based on the gene for gene theory. The famous models are the interactions with the fungal mold Cladosporium fulvum [8], the bacterial speck Pseduomonas syringae pv. Tomato and the fungal wilt Verticillium dahliae [9]. Tomato expresses a large number of defense compounds and is also used as a model plant to test whether an elicitor or a particular pathogen is able to induce basal resistance or to activate forms of induced resistance through SA or JA/ET signaling pathways. In the last twenty years, this plant has been successfully used as a model plant to investigate the induction of defense pathways after exposure to fungal, bacterial and abiotic molecules, showing triggering of different mechanisms of resistance. Understanding these mechanisms in order to improve crop production is a main goal of this study.
Tomato-Pathogen Interactions
Plant-pathogen interactions can be explained by two stages. The first includes interactions between general constitutive plant defense mechanisms and virulence factors produced by the pathogen aimed at destroying the defense. Second, following initial recognition, the plant induces acquired resistance while the pathogen tries to escape from the resistance
General Defense
A plant’s general defense consists of physical and chemical factors. Physical defenses include cutins, which are hardy polymers covering plant external surfaces, pectins that exist in cell walls and middle lamellae to effect adherence between cells, and cell walls, which protect plant cells from external harm. Examples of general chemical defenses are the phytoanticipins such as saponins. Tomato has an antifungal saponin tomatine.
General Pathogenicity-Related Factors and Genes in Tomato Pathogens
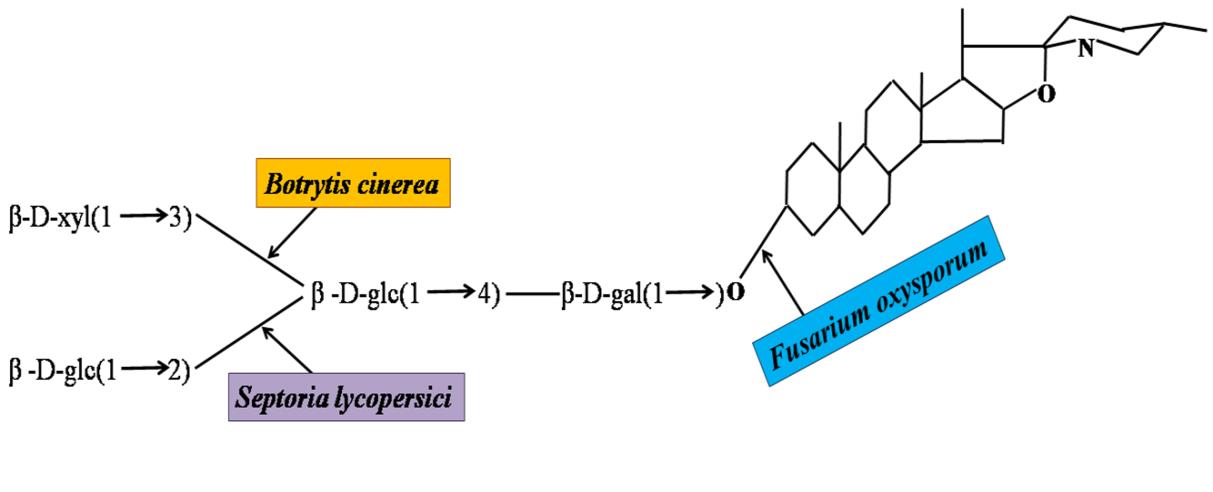
Pathogens of tomato have to nullify the general defenses that tomato originally carries before they establish in tomato. Physical defenses such as cutins, pectins, and cell walls can generally be degraded by cutinases, pectinases, xylanases, and cellulases produced by pathogens, respectively. For example, Fusarium oxysporum f. sp. lycopersici (FOL) secretes pectinases and xylanases. Genes encoding pectinases or xylanases such as pg1, pg5, pgx4, xyl2, and xyl3 have been cloned from FOL Arie, et al. [10], Di Pietro & Roncero [11], Di Pietro & Roncero [12]; García Maceira, et al. [13], Gómez-Gómez, et al. [14], Roldán-Arjona, et al. [15]. Although, pectinases are supposed to be responsible for virulence, disruption of each gene has not eliminated pathogenicity. Probably pectinases are encoded by more than one gene, and lack of a particular pectinase is compensated by the pectinases encoded by other genes Agrios [16], Kawabe, et al. [17]. A chemical defense, tomatine, is enzymatically detoxified by Botrytis cinerea, Septoria lycopersici, and FOL (Figure 1), [15, 16, 18]. From FOL, pathogenicity-related genes such as FPD1 Kawabe, et al. [17] have been reported. From AAL, a cluster of the genes responsible for biosynthesis of AALtoxin has been identified [19]. R. solanacearum has as many as 35 genes homologous to type IV pili genes, which may be involved in cell-to-cell aggregation [20].
Behavior of Pathogens in Tomato Tissues
Behavior of the pathogens on/in tomato tissues has been visualized by immunofluorescence using pathogenspecific antibodies Arie, et al. [21] and using fluorescence protein-expressing pathogen [22]. Detection of pathogens from tomato tissues for diagnostic purposes has been achieved by immunological methods Arie, et al. [21] and PCR. Recently, Hirano & Arie [23] reported primer sets for specific detection of the three races in FOL and F. oxysporum f. sp. radicis-lycopersicon in tomato tissues by PCR. The primer sets and the PCR conditions were also effective for DNA extracted from the tomato rhizospherical soil.
Induced Resistance and Plant Activators
The plant recognizes the pathogen and activates structural and biochemical defenses to protect itself. This phenomenon is called induced or acquired resistance. Usually elicitors, such as carbohydrate, glycoproteins, proteins, peptides, and toxins produced by the pathogens, are recognized by receptors probably on the cell membrane [24, 25]. Systemic acquired resistance (SAR) is a concept in which systemic resistance against pathogens is activated by increased levels of salicylic acid (SA) and pathogenesis related-proteins (PR-proteins) after primary infection with a necrotizing pathogen Ryals, et al. [26], Sticher, et al. [27]; Bostock, et al. [28]; Agrios [16], Meena, et al. [3]. In other words, SA and PR-proteins are the markers of SAR. Other concepts of induced resistance, such as induced systemic resistance (ISR) via jasmonic acid and ethylene as signal molecules, have also been proposed [29, 30]. To check if SAR or ISR is concerned in induced resistance, experiments are now being done with the subsequent mutants of tomato: NahG-Moneymaker (John Innes Center, Norwich, UK), that could be a transformant of cv. Moneymaker carrying the salicylate hydroxylase gene NahG from Pseudomonas putida and cannot accumulate SA Brading, et al. [31], jai1-Micro-Tom, which is a jasmonic acid-insensitive mutant of cv. Micro Tom Li, et al. [32], and Nr-Rutgers, that is an ethylene insensitive mutant of cv. Rutgers Lanahan, et al. [33], these plants are presently accessible. Recently, plant activators like probenazole (PBZ; Meiji Seika Kaisha, Tokyo, Japan), acibenzolar-S-methyl (ASM; Syngenta Japan, Tokyo, Japan), thiadinil (TDL; Nihon Nohyaku, Tokyo, Japan), and harpin (EDEN Bioscience, Bothell, WA, USA), all of which induce SAR in plants have received much attention because they control a wide range of diseases and their efficacy is of long duration. Numerous of these plant activators have been described to persuade SAR in tomato. Only harpin or messenger, is the plant activator registered on tomato in the US. Miyazawa, et al. [34] described that when tomato tissues treated with 4- hydroxybenzoic hydrazide, salicylic hydrazide, or 2-furoic acid by root-dipping, a peroxidase was induced and the treated plants were resistant against wilt caused by FOL. Benhamou & Bélanger [35] reported that treatment of tomato with ASM by foliar spraying was effective against crown and root rot caused by Fusarium oxysporum f. sp. radicis-lycopersici. They discovered that within the root of ASM-treated tomato, pathogen growth was limited to the epidermis and therefore the outer cortex and fungal ingress was halted by the formation of callose-enriched wall appositions at site of fungal penetration. Validamycin A (VMA; Sumitomo Chemical, Tokyo, Japan) is an aminoglucoside formed by Streptomyces hygroscopicus. Even though, VMA does not kill or inhibit the expansion of FOL, a foliar spray of VMA controls soilborne wilt of tomato caused by FOL [36], [37, 38, 39, 40]. The treatment was also effective against bacterial wilt, a soilborne disease caused by Ralstonia solanacearum, and foliar diseases such as powdery mildew caused by Oidium sp. and late blight caused by Phytophthora infestans. They found several days’ time lag between VMA-treatment and emergence of the control effect, and that control efficacy lasted 20-60 days. Moreover, in VMA-treated tomato, accumulation of SA and high transcription of genes programming PR-proteins were detected. They concluded that VMA is a plant activator that induces SAR in treated tomato tissues. Foliar spraying with plant activators could be a particular sensible technique to manage soilborne diseases.
Future Prospects
Tomato is now, and will continue to be, one of the most popular vegetables worldwide. Modern breeding has produced new cultivars resistant to the diseases; however, emergence of new races of pathogens often invalidates the resistance in a short period. Tomato-pathogen interactions still require much investigation to be fully understood. In order to produce healthy tomatoes indefinitely, further analyses of the interaction between tomato and its pathogens are needed; the results may have general relevance to plant-pathogen interactions.
Conflict of Interest
The authors declare no conflicts of interest.
References
-
Meissner R, Jacobson Y, Melamed S, Levyatuv S, Shalev G, et al. (1997) A new model system for tomato genetics. Plant Journal 12(6): 1465-1472.
-
Emmanuel E, Levy AA (2002) Tomato mutants as tools for functional genomics. Current Opinion in Plant Biology 5(2): 112-117.
-
Meena M, Gupta SK, Swapnil P, Zehra A, Dubey MK (2017) Alternaria toxins: potential virulence factors and genes related to pathogenesis. Frontiers in Microbiology 8: 1451.
-
Meena M, Prasad V, Upadhyay RS (2017) Evaluation of Alternaria alternata isolates for metabolite production isolated from different sites of Varanasi, India. Journal of Agricultural Research 2: 000124.
-
Jones JB, Jones JP, Stall RE, Zitter TA (1991) Compendium of tomato diseases. St. Paul.
-
Di Pietro A, Madrid MP, Caracuel Z, Delgado-Jarana J, Roncero MIG (2003) Fusarium oxysporum: exploring the molecular arsenal of a vascular wilt fungus. Molecular Plant Pathology 4(5): 315-325.
-
Babu S, Seetharaman K, Nandakumar R, Johnson I (2004) Biocontrol efficacy of Pseudomonas fluorescens against Alternaria solani and tomato leaf blight disease. Annals of Applied Sciences Research 5(4): 392-403.
-
Joosten MHAJ, de Wit PJGM (1999) The tomato- Cladosporium fulvum interaction: A versatile experimental system to study plant-pathogen interaction. Annual Review of Phytopathology 37: 355-367.
-
Fradin EF, Thomma BPHJ (2006) Physiology and molecular aspects of Verticillium wilt disease caused by V. dahlia and V. albo-atrum. Molecular Plant Pathology 7(2): 71-86.
-
Arie T, Gouthu S, Shimazaki S, Kamakura T, Kimura M, et al. (1998) Immunological detection of endopolygaracturonase secretion by Fusarium oxysporum in plant tissue and sequencing of its encoding gene. Annals of the Phytopathological Society of Japan 64: 7-15.
-
Di Pietro A, Roncero MIG (1996) Endopoly galacturonase from Fusarium oxysporum f. sp. lycopersici: purification, characterization, and production during infection from tomato plants. Phytopathology 86: 1324-1330.
-
Di Pietro A, Roncero MIG (1998) Cloning, expression, and role in pathogenicity of pg1 encoding the major extracellular endopoly galacturonase of the vascular wilt pathogen Fusarium oxysporum. Molecular Plant- Microbe Interactions 11(2): 91-98.
-
García-Maceira FI, Di Pietro A, Roncero MIG (2000) Cloning and distribution of pgx4 encoding an in planta expressed exopolygalacturonase from Fusarium oxysporum. Molecular Plant-Microbe Interactions 13(4): 359-365.
-
Gómez-Gómez E, Isabel M, Di Pietro A, Roncero MI, Hera C (2001) Molecular characterization of a novel endo-beta-1, 4- xylanase gene from the vascular wilt fungus Fusarium oxysporum. Current Genetics 40(4): 268-275.
-
Roldán-Arjona T, Pérez-Ezpinosa A, Ruiz-Rubio M (1999) Tomatinase from Fusarium oxysporum f. sp. lycopersici defines a new class of saponinases. Molecular Plant-Microbe Interactions 12(10): 852- 861.
-
Agrios G N (2005) Plant Pathology. 5th (Edn.). Elsevier, Burlington, MA, USA.
-
Kawabe M, Mizutani K, Yoshida T, Teraoka T, Yoneyama K (2004) Cloning a pathogenicity-related gene, FPD1, in Fusarium oxysporum f. sp. lycopersici. Journal of General Plant Pathology 70(1): 16-20.
-
Ito S, Kawaguchi T, Nagata A, Tamura H, Matsushita H, et al. (2004) Distribution of the FoToml gene encoding tomatinase in different formae speciales of Fusarium oxysporum and identification of a novel tomatinase from F. oxysporum f. sp. radicis-lycopersici, the causal agent of Fusarium crown and root rot of tomato. Journal of General Plant Pathology 70(4):195-201.
-
Yamagishi D, Akamatsu H, Otani H, Kodama M (2006) Pathological evaluation of host-specific AAL-toxins and fumonisin mycotoxins produced by Alternaria and Fusarium species. Journal of General Plant Pathology 72(5): 323-327.
-
Kang Y, Liu H, Genin S, Schell MA, Denny TP (2002) Ralstonia solanacearum requires type 4 pili to adhere to multiple surfaces and for natural transformation and virulence. Molecular Microbiology 46(2): 427- 437.
-
Arie T, Hayashi Y, Yoneyama K, Nagatani A, Furuya M, (1995) Detection of Fusarium spp. in plants with monoclonal antibody. Annals of the Phytopathological Society of Japan 61: 311-317.
-
Nahalkova J, Fatehi J (2003) Red fluorescent protein (DsRed2) as a novel reporter in Fusarium oxysporum f. sp. lycopersici. FEMS Microbiology Letters 225: 305- 309.
-
Hirano Y, Arie T (2006) PCR-based differentiation of Fusarium oxysporum f. sp. lycopersici and radicis- lycopersici and races of F. oxysporum f. sp. lycopersici. Journal of General Plant Pathology 72(5): 273-283.
-
Meena M, Zehra A, Swapnil P, Dubey MK, Patel CB, et al. (2017) Effect on lycopene, b-carotene, ascorbic acid and phenolic content in tomato fruits infected by Alternaria alternata and its toxins (TeA, AOH and AME). Archives of Phytopathology and Plant Protection 50: 317-329.
-
Meena M, Swapnil P, Upadhyay RS (2017) Characterization and toxicological potential of tenuazonic acid, alternariol and alternariol monomethyl ether produced by Alternaria species phytopathogenic on plants. Scientific Reports 7: 8777.
-
Ryals JA, Neuenschwander UH, Willits MG, Molina A, Steiner HY, et al. (1996) Systemic acquired resistance. Plant Cell 8: 1809-1819.
-
Sticher L, Mauch-Mani B, Métraux JP (1997) Systemic acquired resistance. Annual Review of Phytopathology 35: 235-270.
-
Bostock RM, Karban R, Thaler JS, Weyman PD, Gilchrist D (2001) Signal interactions in induced resistance to pathogens and insect herbivores. European Journal of Plant Pathology 107(1): 103-111.
-
van Loon LC, Bakker PAHM, Pieterse CMJ (1998) Systemic resistance induced by rhizosphere bacteria. Annual Review of Phytopathology 36: 453-483.
-
Siddiqui IA, Shaukat SS (2003) Systemic resistance in tomato induced by biocontrol bacteria against the root-knot nematode, Meloidegyne javanica is independent of salycilic acid production. Journal of Phytopathology 152(1): 48-54.
-
Brandwagt BF, Mesbah LA, Takken FLW, Laurent PL, Kneppers TJA, et al. (2000) A longevity assurance gene homolog of tomato mediates resistance to Alternaria alternata f. sp. lycopersici toxins and fumonisin B1. Proceedings of the National Academy of Sciences USA 97(9): 4961-4966.
-
Li L, Zhao Y, McCaig BC, Wingerd BA, Wang J (2004) The tomato homolog of CORONATINE-INSENSITIVE1 is required for the maternal control of seed maturation, jasmonate-signaled defense responses, and glandular trichome development. Plant Cell 16(1): 126-143.
-
Lanahan MB, Yen HC, Giovannoni JJ, Klee HJ (1994) The never ripe mutation blocks ethylene perception in tomato. Plant Cell 6: 521-530.
-
Miyazawa J, Tawabata, T Ogasawara N (1998) Induction of an acidic isozyme of peroxydase and acquired resistance to wilt disease in response to treatment of tomato root with 2-furoic acid, 4- hydroxybenzoic hydrazide or salicylic hydrazide. Physiological and Molecular Plant Pathology 52: 115- 126.
-
Benhamou N, Bélanger RR (1998) Benzothiadiazole- mediated induced resistance to Fusarium oxysporum f. sp. radicis lycopersici in tomato. Plant Physiology 118: 1203-1212.
-
Ishikawa R, Shirouzu K, Nakashita H, Lee HY, Motoyama T, et al. (2005) Foliar spray of validamycin A or validoxylamine A controls tomato Fusarium wilt. Phytopathology 95(10): 1209-1216.
-
Arie T, Takahashi H, Kodama M, Teraoka T (2007) Tomato as a model plant for plant-pathogen interactions. Plant Biotechnology 24: 135-147.
-
García-Maceira FI, Di Pietro A, Huertas-Gonzalez MD, RuizRoldan MC, Roncero MIG (2001) Molecular characterization of an endopoly galacturonase from Fusarium oxysporum expressed during early stage of infection. Applied and Environmental Microbiology 67(5): 2191-2196.
-
Gómez-Gómez E, Ruíz-Roldán MC, Di Pietro A, Roncero MI, Hera C (2002) Role in pathogenesis of two endo-b-1,4-xylanase genes from the vascular wilt fungus Fusarium oxysporum. Fungal Genetics and Biology 35(3): 213-333.
-
Meena M, Zehra A, Dubey MK, Aamir M, Gupta VK, et al. (2016) Comparative evaluation of biochemical changes in tomato (Lycopersicon esculentum Mill.) infected by Alternaria alternata and its toxic metabolites (TeA, AOH, and AME). Frontiers in Plant Science 7: 1408.
- The Role of Podocyte Apoptosis and the Involvement of SIRT1 in Diabetic Nephropathy
- Dealcoholization of Beer by Osmotic Distillation for the Beverage Industry
- Biopolymer-Based Edible Packaging- Biomaterials, Methods, and Applications in Food Industry: An Updated Review
- Influence of Bioprocessing Methods on 'China Rice' (Gawal R1), and Soyabean Supplementation on the Quality of Complementary Food
- Cassava (Manihot esculenta) Varietal Growth, Yield and Cyanide Content Performance in Three Sites in the South- Eastern Semi Arid Regions of Kenya
- Food Waste Treatment, Recycling, Management and Production of Value-Products-An Update on Methodologies and Current Trends