Molecular Characterization of Fungal Isolates of Locally Processed Rice from Five Agro-Ecological Zones of Nigeria
The purpose of this study was to characterize fungal isolates contaminating locally processed rice in Nigeria and to assess the genetic similarity in the fungi isolates. Fungal species were characterized by amplification of a segment of the β-tubulin gene using Bt2aF (5’-GGTAACCAAATCGGTGCTGCTTTC-3’) and Bt2bR (5’ACCCTCAGTGT GT GACCCTTGGC-3’) primers and phylogenetic relationship established using the Maximum Likelihood method. Amplification of between 450 and 550bp of the β-tubulin region of the genomic DNA of the fungal isolates was obtained from thirty-eight samples (89%). Members of Aspergillus section flavi and section nigri groups were differentiated as A. flavus, A. parasiticus, A. tubigensis, A. sydowii and A. niger respectively. Molecular method identified Paecilomycesformosus with 99% similarity index while all the morphologically identified Penicillium species were identified to genus level as P. formosus, P. steckii and P. citrinum with a percentage similarity ranging from 95%-100%. Collectotrichum fruticola and Talaromyces flavus from SGS and NGS zone respectively were documented for the first time in the locally processed rice with 100% and 89% similarity index respectively. All the Aspergillus species were closely related to one another and narrowly diverged from the Penicillium species. Aspergilla and Penicillia were the predominant genera identified and the isolates were genetically similar irrespective of their agroecological zones.
Introduction
Identification of microbial species based on morphological and biochemical tests are not always precise [1]. The traditional methods for the identification of fungal species such as direct plating and pour plates methods followed by phenotypic characterization of the fungal species producing the toxins are mainly based on parameters, including colony diameter, colour, size and texture of conidia and conidiophore structures [2]. However, species classification using these methods may be difficult due to extensive divergence of morphological characters produced by a high level of genetic variability both inter- and intra- specific [3]. These traditional methods are less reliable and less sensitive compared to modern molecular techniques which are rapid, specific and sensitive [4] and appear to be more reliable and robust alternatives to differentiate fungal species. Sequencing data are objective and fast, and leads to reliable identification of uncommon species [5]. Recent advances in molecular based methods have led to many useful techniques taking into account the house-keeping genes as well as targeting other gene catalogues such as 26S, 28S, β-tubulin, calmodin and actin [6].
Beta-tubulin gene has enormous significance in fungi including establishing evolutionary relationship among fungal species and phylogenetic marker for fungal classification. Iqbal and Bakeer [7] reported that β-tubulin is among the most conserved genes in fungi while according to Ayliffe, et al. [8], the intron positions within β- tubulin genes may provide some clues of evolutionary relationships among species. Literatures also abound of various authors who have characterized fungi using this gene [9, 10, 11].
Although the contamination of locally processed rice by fungal colonies and their toxins have been reported Egbuta, et al. [12] but this has been restricted to few selected states of the federation therefore necessitating a nationwide investigation. Abdus-Salaam, et al. [13] reported fungal contamination of locally processed rice in Nigeria with a wider coverage of five agro-ecological zones of Nigeria but there still exists paucity of information on the influence of ecological factors on the incidence of fungal infection of locally processed rice, hence the need to investigate the genetic relatedness of the different fungal ;species colonizing rice in the different agro-ecological zones (AEZ) as Nigeria rises up into self-sufficiency in rice production and export projection. The objective of this study therefore was to characterize potential mycotoxigenic fungal species isolated from locally processed rice samples collected from five AEZs of Nigeria, by the amplification of a segment of β-tubulin gene and establish their phylogenetic relationship.
Methodology
Sample Collection
Samples collection was as described by Abdus-Salaam, et al. [13]. Briefly, different processing centers in five AEZs (Sudan savannah, Northern guinea savannah, Southern guinea savannah, Derived savannah and humid forest) of Nigeria, where rice is traditionally soaked, parboiled, dried and milled, were selected for the study. The milled (dehusked) rice samples were collected from available commercial processors in these centers between November 2011 and February 2012. At each AEZ, samples were collected from four different processing centers in each selected state. At each centre, locally processed rice was obtained from four different processors which were then pooled together to represent one composite sample per Center.
Fungal Isolates
The dilution plate technique described by Samson, et al. [14] was used for fungal isolation by plating 0.1ml aliquots on ½ strength Potato Dextrose Agar (PDA) plates supplemented with 0.01% chloramphenicol. A set was incubated at 30°C for 3 days for enumeration of Aspergillus species while the second set was incubated at 25°C for 7 days for enumeration of Fusarium and Penicillium species. colonies that bore resemblance to Aspergillus and Penicillium species were transferred to full strength potato dextrose agar (PDA). The Aspergillus and Penicillium cultures on PDA were incubated unilluminated at 30°C and 25°C respectively for 7 days. Pure cultures were obtained by repeated sub-culturing and the subcultured isolates were incubated for 5-7 days on PDA. Fungal colonies were harvested and a thin spread was observed in lactophenol in cotton blue solution on microscope slides for identification using the microscope. Pure isolates were identified on the basis of morphological characteristics (macro: colony colour, morphology and size; micro: conidia morphology and size) and comparison with appropriate keys in literature Klich [2], Ehrlich, et al. [15], Pitt, et al. [16]. The identified isolates were further maintained on PDA slants by the single colony transfer technique at 4°C. In all, forty- six representative fungal isolates obtained from thirty-eight composited locally milled rice samples Abdus-Salaam, et al. [13] were used for this study.
Genomic Characterization of Fungi Species
Extraction of genomic DNA: Mycelia of fungal isolates stored under water in vial tubes were subcultured on PDA in a Petri dish for 7 days in the dark at room temperature (28±2ºC). DNA was extracted using Zymo kit (Inqaba biotech, South Africa) following manufacturer’s instructions. PCR Amplification of β-tubulin gene, Sequencing and Phylogenetic Analysis: The primers Bt2aF (5’-GGTAACCAAATCGGTGCTGCTTTC-3’) and Bt2bR (5’ACCCTCAGTGTAGTGACCCTTGGC-3’) described by Glass, et al. [17] and as reported by Nouripour-Sisakht, et al. [18] was used.The PCR condition included an initial denaturation at 95ºC for 5 min, followed by 40 cycles of denaturation at 95ºC for 1min, annealing at 51ºC for 1min and a final extension at 72ºC for 10 min. Positive amplicons were sequenced after purification with QIAquick PCR purification kit (Qiagene, Hilden,Germany). Sequencing was done usingBt2aF (5’-GGTAACCAAATCGGTGCTGCTTTC-3’) and Bt2bR (5’ACCCTCAGTGTAGTGA CCCTTGGC-3’) as described by Glass, et al. [17], Hubka, et al. [19]. Gene sequences obtained were compared by aligning with the sequences in the GenBank using (BLASTn) while evolutionary analyses were conducted using Molecular Evolutionary Genetics Analysis (MEGA) 7 according to Kumar, et al. [20].
The Maximum Parsimony method described by Steel, et al. [21] was used. The consistency index was 0.804697, the retention index was 0.770349 and the composite index was 0.619898 for all sites and parsimony-informative sites. The Maximum Parsimony tree was obtained using the Subtree- Pruning-Regrafting (SPR) algorithm [22] with search level 0 in which the initial trees were obtained by the random addition of sequences (10 replicates). The analysis involved 32 nucleotide sequences. Codon positions included were 1st+2nd+3rd+noncoding. There were a total of 557 positions in the final dataset. Evolutionary analyses were conducted using Molecular Evolutionary Genetics Analysis (MEGA) 7 according to Kumar, et al. [20].
Results
Molecular Characterization of Fungi Isolated from Locally Processed Rice in Nigeria
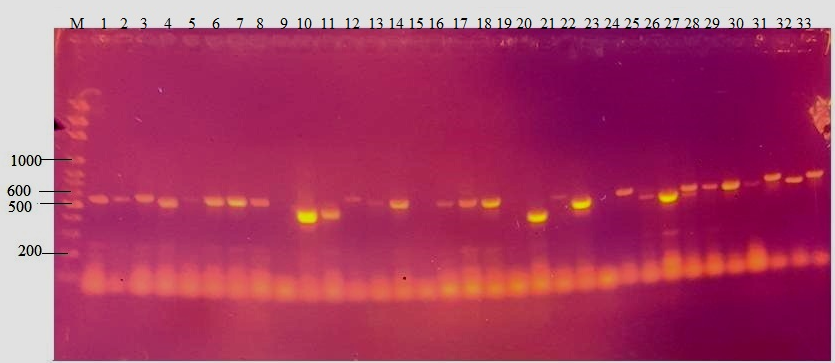
Figure 1 shows amplification of between 450 and 550bp of the β-tubulin region of the genomic DNA of each of the fungal isolates. Amplification was obtained from thirty- eight isolates (89%). Table 1 shows blast result of the nucleotide sequences with the percentage similarities. The Table showed that an isolate from the SGS zone which could not be identified morphologically was identified as Paecilomyces formosus with 99% similarity index. The Penicillium isolates were also identified to species level with a percentage similarity ranging from 95%- 100% as Penicillium citrinum, P. crustosum and P. stekii. However two isolates, each from SGS and NGS that were identified as Penicillium spp. were respectively identified as Colletotrichum fruticola and Talaromyces flavus with 100% and 89% similarity index respectively. One Aspergillus species from the DS zone was clearly identified as Aspergillus sydowii. There was also specific identification of A. tubingensis from Aspergillus section Nigri and A .tamari in Aspergillus section flavi respectively.
Phylogenetic Relationship of Fungal Isolates
The phylogenetic analyses of the fungal isolates (Figure 2) showed that amongst the isolates, Aspergillus flavus was the most recent ancestor with Aspergillus flavus (HF570030.1) as the common ancestral species. The Aspergillus species were closely related to one another and narrowly diverged from the Penicillium species. Next to the ancestral clade were A. parasiticus and A. tamarii which showed a common evolution from A. flavus. A. niger and A. tubigensis shared a common ancestor and individual strains from each specie respectively clustered together except for A. niger (KF669403.1) which formed an outgroup.
| S/ No | Location | Item code | Morphological Identification | Molecular identification | % Similarity | NCBI Acession no |
|---|---|---|---|---|---|---|
| (Closest known identity) | ||||||
| 1 | SGS | 16781 | A.section Flavi | Aspergillus flavus | 100% | KR051523.1 |
| 2 | SGS | 16784 | A.section Flavi | Aspergillus flavus | 99% | KJ482657.1 |
| 3 | SGS | 16785 | A.section Nigri | Aspergillus tubingensis | 98% | KP329861.1 |
| 4 | SGS | 16786 | *UIS | Paecilomyces formosus | 95% | GU968683.1 |
| 5 | SGS | 16788 | A.section Flavi | Aspergillus flavus | 100% | KF434077.1 |
| 6 | SGS | 16789 | A.section Flavi | Aspergillus flavus | 100% | KM491310.1 |
| 7 | SGS | 16790 | A.section Flavi | Aspergillus flavus | 100% | KF562211.1 |
| 8 | SGS | 16792 | Penicillium | Colletotrichum fructicola | 100% | KP852485.1 |
| 9 | SGS | 16794 | Aspergillus spp | Aspergillus niger | 100% | HG325832.1 |
| 10 | SGS | 16797 | A.section Flavi | Aspergillus tamarii | 99% | EU021673.1 |
| 11 | SGS | 16798 | A.section Nigri | Aspergillus niger | 98% | KF669403.1 |
| 12 | SGS | 16799 | A.section Nigri | Aspergillus niger | 98% | KJ136068.1 |
| 13 | SGS | 16801 | A.section Flavi | Aspergillus flavus | 100% | KF434079.1 |
| 14 | DS | 16802 | A.section Nigri | Aspergillus niger | 93% | KY416563.1 |
| 15 | DS | 16803 | Aspergillus spp | Aspergillus sydowii | 100% | KP329915.1 |
| 16 | DS | 16806 | Penicillium | Penicillium citrinum | 99% | KP329977.1 |
| 17 | HF | 16812 | Penicillium | Penicillium steckii | 95% | GU944526.1 |
| 18 | DS | 16815 | Penicillium | Penicillium citrinum | 100% | JX141013.1 |
| 19 | DS | 16816 | A.section Flavi | Aspergillus flavus | 100% | KF434075.1 |
| 20 | DS | 16817 | A.section circumdati | Aspergillus flavus | 100% | KF225072.1 |
| 21 | HF | 16818 | A.section circumdati | Aspergillus flavus | 100% | KF434079.1 |
| 22 | HF | 16819 | A.section circumdati | Aspergillus flavus | 98% | KJ136103.1 |
| 23 | HF | 16822 | Penicillium | Penicillium crustosum | 99% | KJ481238.1 |
| 24 | NGS | 16825 | A.section Flavi | Aspergillus flavus | 99% | HF570030.1 |
| 25 | NGS | 16826 | Penicillium | Talaromyces flavus | 89% | AY766252.1 |
| 26 | SS | 16829 | A.section Flavi | Aspergillus flavus | 100% | KF562206.1 |
| 27 | SS | 16830 | A.section Flavi | Aspergillus flavus | 100% | KC190483.1 |
| 28 | SGS | 16834 | A.section Flavi | Aspergillus flavus | 99% | HQ285476.1 |
| 29 | SGS | 16835 | A.section Flavi | Aspergillus flavus | 100% | KF225050.1 |
| 30 | HF | 16836 | A.section Flavi | Aspergillus parasiticus | 99% | EF203170.1 |
| 31 | DS | 16837 | Aspergillus spp. | Aspergillus tubigensis | 100% | KX961163.1 |
| 32 | DS | 16839 | A.section Flavi | Aspergillus flavus | 99% | KY288823.1 |
Table 1: Molecular Identification of Fungal Isolates. *UIS= Unidentified isolate All strains of Penicillia formed a good cluster
Table 1: Molecular Identification of Fungal Isolates. *UIS= Unidentified isolate All strains of Penicillia formed a good cluster relationship with the exception of P. crustosum (16822) which relate closer with Colletotrichum fructicola. The phylogenic tree also revealed Paecilomyces formosus (16786) and Talaromyces flavus (16826) could have evolved from Penicillium spp.

Discussion
The percentage similarity of the isolates to the reference strains in the gene bank was high which indicated a good index of identification. Rocha, et al. [23] stated that a similarity of less than 40% indicated that isolates do not belong to the same group while an intermediate degree of similarity occurs when strains share 40% to 60% of fragments and still undergo speciation.
An accurate identification of the species of Aspergillus is important because the toxin profile of each species varies and the presence of a particular fungus limits and defines the potential toxicological risks [24]. Aspergillus flavus is a known producer of aflatoxins [25]. The relative abundance of Aspergillus species and high prevalence of A. flavus in this study is supported by previous authors Odhiambo, et al. [26], Egbuta, et al. [12] who reported similar trend in some cereal grains including rice and maize. The high prevalence of A. flavus among the section flavi in many substrates has led to mis-identifications of other members of the section flavi as A. flavus. Tam, et al. [11] reported an identification of A.
tamarii and A. nomius (using ITS, β-tubulin and calmodulin genes) which were previously misidentified by phenotypic characteristics as A. flavus. In this study, members of A. section flavi were differentiated and identified as A. tamarii and A. parasiticus. Although A. tamari is non aflatoxigenic [25], it is however infectious to humans while A. parasiticus is toxigenic producing both B and G aflatoxins [27].
Identification of A. tubingensis which is a member of the species of Aspergillus section nigri and often misidentified as A.niger Palumbo, et al. [28] is a major discovery of this study. Aspergillus niger is an important member of Aspergillus section nigri which despite its industrial importance in the production of enzymes and organic acids, is equally of health significance. Aspergillus niger has been reported to produce toxic metabolites such as malformin, ochratoxin A and other toxic metabolites while Gniadek, et al. [29] has reported the intermediate cytotoxicity of this species. DNA sequencing method identified A. sydowii which is a thermophillic fungus [30]. The thermotolerant nature could enhance this fungus to survive the critical parboiling operation in rice processing. According to Kumar, et al. [31], A.sydowii is commonly associated with rice in the field (73%), on threshing floors (65%), or from storage facilities (78%). It is a potential source of fungal secondary metabolites such as sydowinin, deoxymulundocandin and mulundocandin which possess antimycotic properties and are therefore of biomedical interest [32, 33]. This study provides the first incidence of the fungus Colletotrichum fruticola in Nigerian rice. Reddy, et al. [34] listed Colletotrichum spp. as one of the deleterious seed-borne fungi of rice. Paecilomyces strains are often heat resistant and may produce mycotoxins in contaminated pasteurized foodstuffs [35]. This genus is similar to Penicillium but differs in the absence of greenish coloured colonies and by the short cylindrical phialides that taper into long necks [36]. Some species of Paecilomyces have been produce mycotoxins in foods and feedstuffs and can be considered as a potential source of public health problem.
This study also reports the occurrence of Talaromyces flavus in Nigerian rice for the first time. Laut, et al. [37] also reported the incidence of Talaromyces spp for the first time in rice samples commercialized in Thailand. Talaromyces flavus belongs to the genus Penicillium and has been reported as a heat resistant fungus that can withstand thermal processes [38]. Penicillium citrinum, P. stekii and P. crustosum which were among other species of Penicillium identified in this study have been reported to be of public health concern. Penicillium citrinum is known to produce citrinin; a secondary metabolite which is nephrotoxic while P. crustosum produces penitrem A [39].
The phylogenetic tree demonstrated lack of relationship between the strains and their geographical origins because of the high genetic similarity among each group of fungal isolates. This finding is probably due to the easy dispersal of fungi. Al-Wadai, et al. [10] found that using clustering based on RAPD and ISSR dendograms to study genetic diversity in Aspergillus flavus population was unrelated to geographic origin. De Oliveira Rocha, et al. [23] reported a similar observation in a study of molecular characterization of Fusarium verticillioides isolated from corn grains of different geographical origin in Brazil. Davari, et al. [40] also noted that A. flavus isolates can easily adapt themselves to various geographical regions.
Conclusion
Aspergilla and Penicillia spp. were the prevalent fungal isolates contaminating milled rice in Nigeria while Fusarium species were not detected, despite the occurrence of Fusarium metabolites in the zones. While there were significant differences in the distribution of fungal isolates across the zones, the DS zone recorded the highest occurrence of fungal isolates while the SS zone recorded the least. Furthermore, Colletotrichum fruticola and Talaromyces flavus were documented for the first time in locally processed rice in Nigeria while A. flavus and A. parasiticus were genetically differentiated from one another. There were genetic similarities among the isolates irrespective of the AEZs thus suggesting the possibility of cross contamination of the organisms on the rice samples.
Conflict of Interest
The authors declared no conflict of interest.
References
-
Gallo A, Stea G, Battilani P, Logrieco AF, Perrone G (2012) Molecular characterization of an Aspergillus flavus population isolated from maize during the first outbreak of aflatoxin contamination in Italy. Phytopathol Mediterranea 51(1): 198-206.
-
Klich MA (2002) Identification of common Aspergillus species. Central Bureau voor Schimmel cultures, 1st (Edn.), Utrecht, Netherlands, pp: 116.
-
Kumeda Y, Asao T (1996) Single-strand conformation polymorphism analysis of PCR-amplified ribosomal dna internal transcribed spacers to differentiate species of Aspergillus section flavi. ApplEnviron Microbiol 62(8): 2947-2952.
-
Al-Hindi RR, Aly SE, Hathout AS, Alharbi MG, Al-Masaudi S, Al-Jaouni SK, Harakeh SM (2018) Isolation and molecular characterization of mycotoxigenic fungi in agarwood. Saudi Journal of Biological Sciences 25 (8): 1781-1787.
-
Houbraken J, Verwei PE,Rijs AJMM, Borman AM, Samson RA (2010) Identification of Paecilomyces variotii in clinical samples and settings. J Clin Microbiol 48(8): 2754-2761.
-
Das S, Dash HR, Mangwani N, Chakraborty J, Kumari S (2014) Understanding molecular identification and polyphasic taxonomic approaches for genetic relatedness and phylogenetic relationships of microorganisms. J Microbiol Methods 103: 80-100.
-
Iqbal Z, Bakeer W (2013) Isolation, cloning and sequence analysis ofβ-tubulin gene from bolinea lutea f23523, a potential strobilurin producing fungus. IntJ Agric Biol 15(5): 1008-1012.
-
Ayliffe MA, Dodds PN, Lawrence GJ (2001) Characterisation of a beta-tubulin gene from Melampsora lini and comparison of fungal beta-tubulin genes. Mycology Resarch 105(7): 818-826.
-
Kredics L, Varga J, Kocsube S, Doczi I, Samson RA, et al. (2007) Case of keratitis caused byAspergillus tamarii. J Clin Microbiol 45(10): 3464-3467.
-
Al-Wadai AS, Al-Othman MR, Mahmoud MA, Abd El-Aziz ARM (2013) Molecular characterization of Aspergillus flavus and aflatoxin contamination of wheat grains from Saudi Arabia. Genet Mol Res 12 (3): 3335-3352.
-
Tam EWT, Chen JHK, Lau ECL, Ngan AHY, Fung KSC, et al. (2014) Misidentification of Aspergillus nomius and Aspergillus tamarii as Aspergillus flavus: Characterization by internal transcribed spacer, beta-tubulin, and calmodulin gene sequencing, metabolic fingerprinting, and matrix-assisted laser desorption ionization-time of flight mass spectrometry. J Clin Microbiol 52(4): 1153- 1160.
-
Egbuta MA, Wanza MM, Dutton MF (2015) Evaluation of five major mycotoxins co-contaminating two cereal grains from Nigeria. Int J Biochem Res Rev 6 (4): 160- 169.
-
Abdus-Salaam R, Fanelli F, Atanda O, Sulyok M, Cozzi G, et al. (2016) Fungal isolates and metabolites in locally processed rice from five agro-ecological zones of Nigeria. Food Addit Contam B Surveill 9(4): 281-289.
-
Samson RA, Hoekstra ES, Lund F, Frisvad JC, Filtenborg O (1995) Methods for the detection and isolation of food- borne fungi. In: Samson RA, et al. (Eds.), Introduction to Foodborne Fungi. Central Bureau voor Schimmel cultures, Netherlands, pp: 235-242.
-
Ehrlich KC, Kobbeman K, Montalbano BG, Cotty PJ (2007) Aflatoxin producing Aspergillus species from Thailand. Int J Food Microbiol 114(2): 153-159.
-
Pitt JI, Hocking AD (2009) Fungi and food spoilage. Springer, London.
-
Glass NL, Donaldson GC (1995) Development of primer sets designed for use with the pcr to amplify conserved genes from filamentous ascomycetes.Appl Environ Microbiol 61(4): 1323-1330.
-
Nouripour-Sisakht S, Mirhendi H, Shidfar MR, Ahmadi B, Rezaei-Matehkolaei A, et al. (2015) Aspergillus species as emerging causative agents of onychomycosis. J Mycol Med 25(2): 101-107.
-
Hubka V, Kolarik M (2012) Beta-tubulin paralogue tubC is frequently misidentified as the benA gene in Aspergillus section Nigri taxonomy: primer specificity testing and taxonomic consequences. Persoonia 29: 1-10.
-
Kumar S, Stecher G, Tamura K (2016) MEGA7: Molecular evolutionary geneticsanalysis version 7.0 for bigger datasets. Mol Biol Evol 33(7): 1870-1874.
-
Steel M, Penny Y (2000) Parsimony, Likelihood, and the Role of Models in Molecular Phylogenetics. Mol Biol Evol 17(6): 839-850.
-
Nei M, Kumar S (2000) Molecular Evolution and Phylogenetics. Oxford University Press, New York, USA.
-
Rocha LO, Reis GM, Da Silva VN, Braghini R, Corrêa B, et al. (2011) Molecular characterization and fumonisin production by Fusarium verticillioides isolated from corn grains of different geographic origins in Brazil. Int J Food Microbiol 145(1): 9-21.
-
Spadaro D, Patharajan S, Lorè A, Garibaldi A, Gullino ML (2012) Ochratoxigenic black species of Aspergilli in grape fruits of northern Italy identified by an improved PCR-RFLP procedure. Toxins 4(2): 42-54.
-
Rodrigues P, Soares C, Kozakiewicz Z, Paterson R, Lima N (2007) Identification and characterization of Aspergillus flavus and Aflatoxins. In: Mendez-Souza A (Eds.), Communicating current research and educational topics and trends in applied microbiology, pp: 526-534.
-
Odhiambo BO, Murage H, Wagara N (2013) Isolation and characterisation of aflatoxigenic Aspergillus species from maize and soil samples from selected counties of Kenya. Afr J Microbiol Res 7(34): 4379-4388.
-
Klich MA, Pitt JI (1988) A laboratory guide to the common Aspergillus species and their teleomorphs. Common wealth scientific and industrial research organisation, division of food processing, North Ryde, Australia, pp: 116.
-
Palumbo JD, Okeeffe TL (2015) Detection and discrimination of four Aspergillus section Nigri species by PCR. Lett Appl Microbiol 60(2): 188-195.
-
Gniadek A, Macura AB, Twaruż M (2009) Characteristics Fungi Present in the Intensive Care Unit Environment. Part 2 Aspergillus Fungi Mikologia Lekarska 16(1): 15- 18.
-
Mushtaq M, Khatoon F, Batool S (2014) Optimization of culture conditions for the production of endoglucanase from Aspergillus sydowii using corn cobs. Pinnacle Biochem. Res 1(1): 202-208.
-
Kumar V, Basu MS, Rajendran TP (2008) Mycotoxin research and mycoflora in some commercially important agricultural commodities. Crop Protec 27(6): 891-905.
-
Mukhopadhyay T, Roy K, Bhat RG, Sawant SN, Blumbach J, et al. (1992) Deoxymulundocandin--a new echinocandin type antifungal antibiotic. J Antibiot (Tokyo) 45(5): 618- 623.
-
Schmitt EK, Eilinghoff B, Olliger R, Decker H, Kuck U (2002) Development of molecular tools for the mulundocandin producer Aspergillus sydowii: DNA- mediated transformation and reporter gene expression. Applied Microbiology Biotechnology 58(5): 625-631.
-
Reddy OR, Sathyanarayana N (2001) Seed borne fungi of rice and quarantine significance. In: Sreenivasaprasad S, et al. (Eds.), Major Fungal Disease of Rice. Recent Advances, pp: 331-345.
-
Samson RA, Houbraken J, Varga J, Frisvad JC (2009) Polyphasic taxonomy of the heat resistant ascomycete genus Byssochlamys and its Paecilomyces anamorphs. Persoonia 22(1): 14-27.
-
Borba C, Brito MM (2015) Paecilomyces: mycotoxin production and human infection. In: Paterson RM, et al. (Eds.), Molecular biology of food and water borne mycotoxigenic and mycotic fungi. 1st(Edn.), CRC Press, Boca Raton, pp: 401-422.
-
Laut S, Poapolathep S, Piasai O, Sommai S, Boonyuen N, et al. (2023) Storage Fungi and Mycotoxins Associated with Rice Samples Commercialized in Thailand. Foods 12(3): 487.
-
Amaeze NJ, Ugwuanyi JO, Obet JAN (2010) Studies of heat resistant fungi in the soil: Talaromyces flavus isolated in Nigerian soils. New York Science Journal 3 (12): 8-14.
-
Doughari JH (2015) The occurrence, properties and significance of citrinin mycotoxin. J Plant Pathol Microbiol 6(11): 1000321.
-
Davari E, Mohsenzadeh M, Mohammadi GH, Rezaeian- Doloei R (2015) Characterization of aflatoxigenic Aspergillus flavus and A. parasiticus strain isolates from animal feedstuffs in northeastern Iran. Iran J Vet Res 16(2): 150-155.
- The Role of Podocyte Apoptosis and the Involvement of SIRT1 in Diabetic Nephropathy
- Dealcoholization of Beer by Osmotic Distillation for the Beverage Industry
- Biopolymer-Based Edible Packaging- Biomaterials, Methods, and Applications in Food Industry: An Updated Review
- Influence of Bioprocessing Methods on 'China Rice' (Gawal R1), and Soyabean Supplementation on the Quality of Complementary Food
- Cassava (Manihot esculenta) Varietal Growth, Yield and Cyanide Content Performance in Three Sites in the South- Eastern Semi Arid Regions of Kenya
- Food Waste Treatment, Recycling, Management and Production of Value-Products-An Update on Methodologies and Current Trends