The Long Arm of the Very Small: Human Microbiome and Disease
It is a well-known fact that we share our body with a large number of prokaryotic cells and viruses that reside on all surfaces of our body and make up what is nowadays mostly referred to as the human microbiota. An estimate of cell numbers puts the ratio prokaryotic to eukaryotic cell count to 10:1 and the size of the microbial genome to 100 times the size of the human genome. While it is in some respect accepted and expected that the presence of the microbiota has a direct effect on the organ in which it resides, it is becoming increasingly evident that they can affect far organs with devastating metabolic and physiological effects on the host. This mini review highlights recent research exploring the impact of the microbiome on far organs and host metabolism.
Microbiome and Cancer
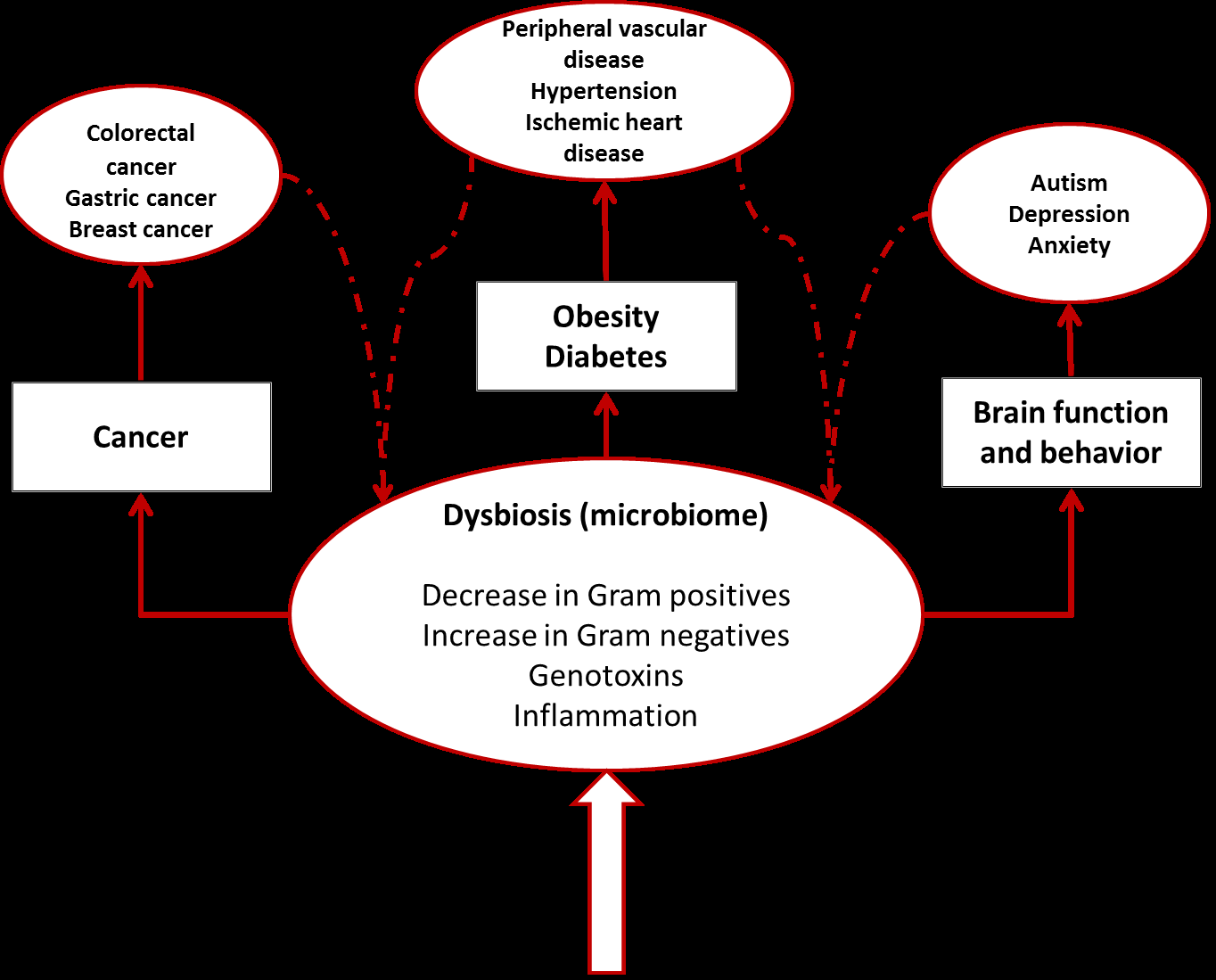
Cancer results from a multi-step loss of physiological controls in the host cells so that they acquire the ability to invade local tissues and spread widely. Interactions with the microbiome can influence development of cancer through induced inflammation and metabolism. The intestinal microbiota is one of several factors that can promote the development of local colorectal cancer. Disbyosis caused by age, antibiotics, xenobiotics, smoking, hormones, and dietary changes, is a well-known factor that can promote the progression to malignancy [11]. It is however difficult to establish whether a change in the microbiota is responsible for promoting cancer, or the tissue physiological changes due to the presence of malignancy induce changes in the composition of the microbiota. It has been recently demonstrated that specific strains of E. coli (phylogroup B2) produce cyclomodulins which are genotoxic and/or modulate cell- cycle progression [12] shifting the paradigm towards a more causative role for the microbiota. The microbiome is an active parallel organ producing a large and diverse range of metabolic products that can influence host metabolism in organs that are far from the location of the microbiota itself. The first suggestion of involvement of the intestinal microbiota in the development of breast cancer was derived from results that showed that injections with DMAB (a well-known carcinogen) in germ-free rats resulted in significantly lower cancer development, as compared to conventionalized rats [13]. Plottel suggests that the influence of the microbiome on the development of breast cancer could be linked directly to metabolic products of the intestinal microbiota. Amongst the many activities of the microbial cells it is possible to identify a functional estrobolome [14] due to bacterial genes whose products can metabolize estrogen. A microbiota that favors the production of enzymes promoting deconjugation of estrogen metabolites, results in re-absorption of free estrogen increasing the total host estrogen burden [14]. There is a direct association between circulating estrogen levels and risk of development of endometrial cancer in post-menopausal women [15]; manipulation of the microbiome to influence specifically the presence of species with beta- glucorunosidase and beta-glucoronide activities could constitute an approach to reduce estrogen related cancer risk [14].
Cancer of the liver is also strongly correlated with conditions of the intestinal microbiota. Alcoholic cirrhosis results in elevated serum content of Gram negative associated endotoxins, and germ-free animals are protected from alcohol induced liver damage [16]. Ethanol-induced hepatic translocation of LPS and Gram- negative bacteria may further synergize with these direct effects in activating the innate immune response [17].
Brain Function and Behavior
The gut microbiota is involved in regulating neurophysiological-governed behaviors through immune, endocrine, and neural pathways. The bidirectional communication between the gut and the brain is known as the microbiota-gut axis and alterations of the microbiota composition can influence central nervous system-related disease and neurological functions [18]. Appetite is regulated by gut hormones produced by enteroendocrine cells located in the intestinal mucosa. Microbial products of fermentation such as acetate and butyrate can be recognized by receptors on the enteroendocrine cells inducing the production of PYY (peptide tyrosine-tyrosine) and GLP-1 (glucagon-like peptide 1) which are known appetite suppressor [19]. Psychiatric disorders have been recognized as causative of changes in the composition of gut microbiota. It has been demonstrated that children with autism spectrum disorders show increased Lactobacillus species, and decreased Bifidobacterium and Prevotella species [20]. It is however proving difficult to demonstrate whether a change in the microbiota can be responsible for the development of neuropsychiatric disorders. Obesity is associated with increased oxidative stress, which has been correlated with cognitive decline. Transplantation of gut microbiota from High Fat Diet (HFD)-fed mice to mice with depleted microbiota due to antibiotic treatment, disrupts explorative, cognitive, and stereotypical behavior when compared to mice transplanted with microbiota from a control diet [19, 21]. Gut microbial metabolism leads to the production of catecholamine, histamine, and other neuroactive mediators that stimulate local enteric nervous system and/or primary afferent fibers [21]. The bacterium Lactobacillus rhamnosus specifically modulates GABA receptors expression reducing stress-induced elevation of corticosterone [22]. Animals fed with L. rhamnosus increase GABA receptor expression in the prefrontal International Journal of Biochemistry & Physiology
cortex and reduce its expression in the amigdala, hippocampus, and LC). Amongst the behavioral changes observed, mice fed with L. rhamnosus showed enhanced memory to aversive clues and context [22]. Recent research on HFD mice implicates short-chain fatty acid metabolism by the gut microbiota with the expression of circadian clock genes in hepatocytes [23]. In the presence of gut microbiota, HFD leads to the production of altered microbiome derived metabolic signals which result in disturbed circadian clock rhythm that, in turn, leads to obesity [23]. These interactions between gut and brain could be mediated through Microbiota-Associated Molecular Patterns (MAMPs) and Pattern Recognition Receptors (PPRs). It appears that Gram positive bacteria such as Lactobacillus and Bifidobacterium have a positive effect in alleviating anxiety [19].
Metabolic Disease, Obesity, and Diabetes
The obesity epidemic currently affecting the world is a major reason for increased insulin resistance and type 2 diabetes mellitus. Animal models of obesity have provided many clues on the relationship between diet, microbiome, and diabetes [24]. Several studies are indicating that the microbiome has an important role in regulating fat storage and energy consumption, microbial processing of components of the diet leads to deposition of the extracted energy in host fat deposits [25] and the microbiome could induce a more efficient metabolism to harvest energy via the production of short-chain fatty acids or through pro-inflammatory signals and establishment of chronic inflammation, or a combination of both [26]. Experiments on germ-free mice colonized with gut microbial communities derived from conventionally raised mice result in marked increase of body fat within 10-14 days [25]. The proposed mechanisms for this change are:
a) Microbial digestion of polysaccharides that cannot be digested normally by the host, associated with absorption of monosaccharides and short-chain fatty acids. b) Conversion by the liver of these products to more complex lipids. c) Microbial regulation of host genes that promote deposition of lipids into adipocytes (Angiopoietin-like protein 4 – Fiaf) [25].
It has been now proposed that the “obese microbiome” has increased capacity to harvest energy from the diet. Moreover, colonization of germ-free mice with “obese microbiota” produce a greater increase of body fat than colonization with “lean microbiota” [26]. In addition to dysbiosis of the lower intestinal tract, the oral microbiota can also be implicated. Overweight and healthy weight individuals show differences in diversity and abundance of salivary bacteria with Prevotella in greater abundance in overweight individuals and Selenomonas present only in the overweight population [27]. Obesity related inflammation results in an increase of pro-inflammatory cytokines such as tumor necrosis factor α (TNF-α), which can result in insulin resistance [28]. Gram negative bacteria such as Bacteroidetes contain lipopolysaccharide (LPS) which is a known promoter of inflammation. High fat diets tend to reduce the number of Gram positive bacteria and to favor Gram negative colonization of the gut. Increase of Bifidobacteria in the gut has been shown to reduce inflammation and improve glucose tolerance [29, 30]. The microbiota is therefore responsible for the regulation of both energy harvest and expenditure. Recent studies based on fecal transplantation in humans, show beneficial alterations in glucose metabolism after lean donor transplantation [31, 32]. Type I diabetes is an autoimmune disease characterized by the presence of autoantibody against pancreatic insulin-producing β cells. Gut microbiota can play an important role in regulation of T cells populations in the gut. Non obese diabetic (NOD) mice lacking MyD88 protein (an essential Toll-like receptor signal transducer) don’t develop type I diabetes [33, 34]. Stools from bio- breeding (BB) diabetes resistant rats contain more probiotic-like bacteria, while Bacteroides, Eubacterium, and Ruminococcus are more prevalent in BB diabetes prone rats [35, 36].
Conclusion
It is becoming more and more evident that the microbiota that shares our body resources is involved in the homeostasis of many human processes and can have profound effects on far organs and tissues with important repercussions for human health. The next few years will provide important understanding of these mechanisms International Journal of Biochemistry & Physiology
and potentially will provide new therapeutic strategies for treatment of disease.
References
-
Palm NW, de Zoete MR, Flavell RA (2015) Immune- microbiota interactions in health and disease. Clin Immunol 159(2): 122-127.
-
Minot S, Bryson A, Chehoud C, Wu GD, Lewis JD, et al. (2013) The rapid evolution of the human gut viriome. Proc Natl Acad Sci U S A 110(30): 12450-12455.
-
Savage DC, Siegel JE, Snellen JE, Whitt DD (1981) Transit Time of Epithelial Cells in the Small Intestines of Germfree Mice and Ex-Germfree Mice Associated with Indigenous Microorganisms. Appl Environ Microbiol 42(6): 996-1001.
-
Cox LM, Yamanishi S, Sohn J, Alekseyenko AV, Leung JM, et al. (2014) Altering the intestinal microbiota during a critical developmental window has lasting metabolic consequences. Cell 158(4): 705-721.
-
Arrieta MC, Stiemsma LT, Amenyogbe N, Brown EM, Finlay B (2014) The intestinal microbiome in early life: health and disease. Front Immunol 5: 427.
-
Backhed F, Fraser CM, Ringel Y, Sanders ME, Sartor RB, et al. (2012) Defining a Healthy Human Gut Microbiome: Current Concepts, Future Directions, and Clinical Applications. Cell Host and Microbiome 12(5): 611-622.
-
Manichanh C, Rigottier-Gois L, Bonnaud E, Gloux K, Pelletier E, ET AL. (2006) Reduced diversity of faecal microbiota in Crohn’s disease revealed by a metagenomic approach. Gut 55(2): 205-211.
-
Wu HJ, Ivanov II, Darce J, Hattori K, Shima T, et al. (2010) Gut-residing segmented filamentous bacteria drive autoimmune arthritis via T helper 17 cells. Immunity 32(6): 815-827.
-
Marchesi JR, Dutilh BE, Hall N, Peters WHM, Roelofs R, et al. (2011) Towards the Human Colorectal Cancer Microbiome. PLoS One 6(5): e20447.
-
Sobhani I, Amiot A, Le Baleur Y, Levy M, Auriault ML, et al. (2013) Microbial dysbiosis and colon carcinogenesis: could colon cancer be considered a bacteria-related disease? Therap Adv Gastroenterol 6(3): 215-229.
-
Zitvogel L, Galluzzi L, Viaud S, Vétizou M, Daillère R, et al. (2015) Cancer and the gut microbiota: an unexpected link. Sci Transl Med 7(271): 271ps1.
-
Bonnet M, Buc E, Sauvanet P, Darcha C, Dubois D, et al. (2014) Colonization of human gut by E. coli and colorectal cancer risk. Clin. Cancer Res 20(4): 859- 867.
-
Reddy BS, Watanabe K (1978) Effect of intestinal microflora on 3-2'-dimethyl-4-aminobiphenyl- induced carcinogenesis in F344 rats. J Nat Cancer Inst 61(5): 1269-1271.
-
Plottel CS, Blaser MJ (2011) Microbiome and malignancy. Cell Host Microbe 10(4): 324-335.
-
Lukanova A, Lundin E, Micheli A, Arslan A, Ferrari P, et al. (2003) Circulating levels of sex steroid hormones and risk of endometrial cancer in postmenopausal women. Int J Cancer 108(3): 425- 432.
-
Canesso MCC, Lacerda NL, Ferreira CM, Gonçalves JL, Almeida D, et al. (2014) Comparing the effects of acute alcohol consumption in germ-free and conventional mice: the role of the gut microbiota. BMC Microbiology 14: 240.
-
Pevsner-Fischer M, Tuganbaev T, Meijer M, Zhang SH, Zeng ZR, et al. (2016) Role of the microbiome in non- gastrointestinal cancers. World J Clin Oncol 7(2): 200- 213.
-
Kelly JR, Kennedy PJ, Cryan JF, Dinan TG, Clarke G, et al. (2015) Breaking down the barriers: the gut microbiome, intestinal permeability and stress- related psychiatric disorders. Front Cell Neurosci 9: 392.
-
Mu C, Yang Y, Zhu W (2016) Gut Microbiota: The Brain Peacekeeper. Front Microbiol 7: 345.
-
Kang D-W, Park JG, Ilhan ZE, Wallstrom G, LaBaer J, et al (2013) Reduced Incidence of Prevotella and Other Fermenters in Intestinal Microflora of Autistic Children. PLoS ONE 8(7): e68322.
-
Bruce-Keller AJ, Salbaum JM, Luo M, Blanchard E , Taylor CM, et al. (2015) Obese-type Gut Microbiota Induce Neurobehavioral Changes in the Absence of Obesity. Biol Psychiatry 77(7): 607-615. International Journal of Biochemistry & Physiology
-
Bravo JA, Forsythe P, Chew MV, Escaravage E, Savignac HM, et al. (2011) Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve . PNAS 108(38): 16050-16055.
-
Leone V, Gibbons SM, Martinez K, Hutchison AL, Huang EY, et al. (2015) Effects of diurnal variation of gut microbes and high fat feeding on host circadian clock function and metabolism. Cell Host Microbe 17(5): 681-689.
-
Turnbaugh PJ, Ridaura VK, Faith JJ, Rey FE, Knight R, et al. (2009) The Effect of Diet on the Human Gut Microbiome: A Metagenomic Analysis in Humanized Gnotobiotic Mice. Sci Transl Med 1(6): 6ra14.
-
Bäckhed F, Ding H, Wang T, Hooper LV, Koh GY, et al. (2004) The gut microbiota as an environmental factor that regulates fat storage. Proc Natl Acad Sci U S A 101(44): 15718-15723.
-
Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER (2006) An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 444: 1027-1031.
-
Goodson JM, Groppo D, Halem S, Carpino E (2009) Is obesity an oral bacterial disease? J Dent Res 88(6): 519-523.
-
Hotamisligil GS, Peraldi P, Budavari A, Ellis R, White MF, et al. (1996) IRS-1-mediated inhibition of insulin receptor tyrosine kinase activity in TNF-alpha-and obesity-induced insulin resistance. Science 271(5249): 665-668.
-
Ma X, Hua J, Li Z (2008) Probiotics improve high fat diet induced hepatic steatosis and insulin resistance by increasing NKT cells. J Hepatol 49(5): 821-830.
-
Cani PD, Neyrinck AM, Fava F, Knauf C, Burcelin RG, et al. (2007) Selective increases of bifidobacteria in gut microflora improve high-fat-diet-induced diabetes in mice through a mechanism associated with endotoxaemia. Diabetologia 50(11): 2374-2383.
-
Vrieze A, Van Nood E, Holleman F, Salojärvi J, Kootte RS, et al. (2012) Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Gastroenterology 143(4): 913-6e7.
-
Kootte RS, Vrieze A, Holleman F, Dallinga-Thie GM, Zoetendal EG, et al. (2012) The therapeutic potential of manipulating gut microbiota in obesity and type 2 diabetes mellitus. Diabetes Obes Metab 14(2): 112- 120.
-
Wen L, Ley RE, Volchkov PY, Stranges PB, Avanesyan L (2008) Innate immunity and intestinal microbiota in the development of Type 1 diabetes. Nature 455: 1109-1113.
-
Burrows MP, Volchkov P, Kobayashi KS, Chervonsky AV (2015) Microbiota regulates type 1 diabetes through Toll-like receptors. PNAS 112(32): 9973- 9977.
-
Roesch LF, Lorca GL, Casella G, Giongo A, Naranjo A, et al. (2009) Culture-independent identification of gut bacteria correlated with the onset of diabetes in a rat model. ISME J 3(5): 536-548.
-
de Goffau MC, Luopajarvi K, Knip M, Ilonen J, Ruohtula T, et al. (2013) Fecal Microbiota Composition Differs Between Children With β-Cell Autoimmunity and Those Without. Diabetes 62(4): 1238-1244.
-
Consortium TH (2012) Structure, Function and Diversity of the Healthy Human Microbiome. Nature 486(7402): 207-214.
- Superposition of Cryo-EM and AlphaFold Predictions of Dengue Antigen-Antibody Complexes
- Jugular-Applied Coherent Low-Level Laser Therapy Enhances Systemic Mitochondrial Metabolic Function and Antioxidant Response
- Role of OMC32 Polypeptide in Acrosin-Mediated Exocytosis during the Bovine Sperm Acrosome Reaction
- Association of Galectin-3 but not Laminin in Tamoxifen-Induced Growth Suppression in Breast Cancer MCF-7 Cells
- Effect of Different Wavelengths of Light on the Rate of Photosynthesis
- Nutritional, Therapeutic, and Environmental Effect of Oyster Mushrooms: An Editorial