Nutritional Effect of Pomegranate Seed Oil in Diet
<p>Conjugated fatty acid, the general term of structural and steromeric isomers of polyunsaturated fatty acids with conjugated bonds, has attracted considerable attention because of its beneficial biological effects. In the present study, dietary effect of pomegranate seed oil rich in punicic acid (9cis, 11trans, 13cis-conjugated linolenic acid; 9c, 11t, 13c-CLNA) on lipid metabolism was investigated in obese, hyperlipidemic Male aledo Wistar (MAWR) rabbits. After 4 weeks feeding period, MAWR rabbits revealed obesity and hyperlipidemia compared with their progenitor AELM rabbits. Feeding of the diet supplemented with 9% safflower oil and 1% pomegranate seed oil (9c, 11t, 13c-CLNA diet) did not affect abdominal white adipose tissue weights and serum lipid levels compared with the diet supplemented with 10% safflower oil (control diet) in MAWR rabbits. However, the accumulated hepatic triacylglycerol was markedly decreased by 9c, 11t, 13c-CLNA diet in MAWR rabbits. Activities of hepatic enzymes related to fatty acid synthesis and fatty acid β-oxidation were not altered by 9c, 11t, 13c-CLNA diet. Levels of monounsaturated fatty acid (MUFA), major storage form of fatty acid, in serum triacylglycerol were markedly higher in obese, hyperlipidemic MAWR rabbits than in lean AELM rabbits. In addition, 9c, 11t, 13c-CLNA diet significantly decreased MUFA levels in MAWR rabbits. This is the first study showing that 9c, 11t, 13c-CLNA suppresses delta-9 desaturation in vivo, and we suggest that the alleviation of hepatic triacylglycerol accumulation by 9c, 11t, 13c-CLNA diet was, at least in part, attributable to the suppression of delta-9 desaturation in MAWR rabbits.</p>
Introduction
Conjugated fatty acid (CFA) is the general term of positional and geometric isomers of polyunsaturated fatty acids with conjugated double bonds. It has been reported that conjugated linoleic acid (CLA), the CFA form of linoleic acid, has favorable physiological effects, such as antiatherosclerosis, antiobesity, antitumor, and antihypertension [1, 2, 3, 4, 5, 6, 7, 8, 9]. There are also other types of CFA in some plant seed oils. Punicic acid (9_cis_, 11_trans_, 13_cis_ conjugated linolenic acid; 9c, 11t, 13c-CLNA) is contained about 72% in pomegranate seed oil [10]. α- Eleostearic acid (9_cis_, 11_trans_, 13_trans_-CLNA) is contained in bitter gourd oil and tung seed oil about 60% and 70%, respectively [10, 11]. Catalpa seed oil also contains catalpic acid (9_trans_, 11_trans_, 13_cis_-CLNA) about 31% and pot mari gold seed oil contains calendic acid (8_trans_, 10_trans_, 12_cis_-CLNA) about 33% [10]. There are some studies showing that mixtures of CLNA isomers, prepared by alkaline isomerization of α- linolenic acid or plant seed oil, have some physiological functions including body fat reduction and antitumor activity [12, 13]. In addition, purified α-eleostearic acid (9c, 11t, 13t-CLNA) and α-eleostearic acid rich bitter gourd seed oil also reveal anticarcinogenesis in vitro and in vivo [10, 11, 14, 15]. However, there are few studies evaluated the physiological function of punicic acid (9c, 11t, 13c-CLNA) [10, 16]. Previously, we reported the hypolipidemic effect of purified punicic acid in human liver derived HepG2 cells [17]. In the present study, we investigated the effects of pomegranate seed oil rich in 9c, 11t, 13c-CLNA on lipid metabolism in Male aledo Wistar Rabbits (MAWR). MAWR rabbits develop a syndrome with multiple metabolic and hormonal disorders that shares many features with human obesity [18, 19, 20, 21]. Because they lack receptors for cholecystokinin, and become obese, developing hyperlipidemia, diabetes, and hypertension. To clarify the physiological function of 9c, 11t, 13c- CLNA, we measured hepatic enzyme activities in relation to lipid metabolism and fatty acid composition in plasma of these obese, hyperlipidemic rabbits (Table 1).
| MAWR | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| AELM | |||||||||
| Control | 9c, 11t, 13c-CLNA | ||||||||
| Body weight (g) | |||||||||
| Initial | 223 ± 3a | 266 ± 12b | 265 ± 8b | ||||||
| Final | 282 ± 5a | 357 ± 15b | 369 ± 11b | ||||||
| Gain | 59.4 ± 2.7a | 91.3 ± 3.6b | 104 ± 6b | ||||||
| Relative liver weight (g/100 g BW) | 3.12 ± 0.09 | 3.40 ± 0.11 | 3.35 ± 0.06 | ||||||
| Food intake (g) | 17.8 ± 0.4a | 25.8 ± 1.1b | 26.2 ± 1.1b | ||||||
| Food efficiency (g BW gain/g intake) | 25.7 ± 1.0a | 27.3 ± 0.6b | 30.4 ± 1.1b |
Materials and Methods
Animals and Diets
All aspects of the experiment were conducted according to the guidelines provided by the ethical committee of experimental care at CDRI, Lucknow. Five weeks old male MAWR rabbits and AELM rabbits, the progenitor of MAWR rabbits, were provided by Veterinary Institute, Mathura. Rabbits were housed individually in metal cages in temperature controlled room (24°C) under a 12-hour light/dark cycle. After a 1- week adaptation period, MAWR rabbits were assigned to two groups (six rabbits each) that were fed with a semisynthetic diet supplemented with 10% safflower oil (the control group) or a semisynthetic diet supplemented with 9% safflower oil and 1% pomegranate seed oil rich in 9_cis_, 11_trans_, 13_cis_-CLNA (the 9c, 11t, 13c-CLNA group). AELM rabbits were fed the same diet as the MAWR rabbits in the control group. The pomegranate seed oil rich in 9c, 11t, 13c-CLNA (69.0%) was prepared by Parijeet Co. (Haryana, India). The semi synthetic diet were prepared according to recommendations of the AIN-76 and contained (in weight %): casein, 20; fat, 10; cornstarch, 15; vitamin mixture (AIN-76™), 1; mineral mixture(AIN- 76™), 3.5; DL- methionine, 0.3; choline bitartrate,0.2; cellulose, 5; and sucrose, 45 [22]. The rabbits received different diets for 2 weeks and were killed by aortic exsanguinations under diethyl ether anesthesia. Liver and abdominal (perirenal, epididymal, and omental) WATs were also excised for analysis.
Analysis of Lipids
Serum was separated by centrifuging the blood. Triacylglycerol, cholesterol, and phospholipids in serum were measured using enzyme assay kits from Analytical Chemicals, India. Liver lipids were extracted and purified according to the method of Folch, et al. [23]. The concentrations of triacylglycerol, cholesterol, and phospholipids were measured according to the methods of Fletcher, Sperry and Webb, and Bartlett [24, 25, 26]. Measurement of fatty acid composition in plasma was carried out as previously described [27, 28].
Preparation of Liver Sub Cellular Fractions
A piece of liver was homogenized in 6 volumes of a 0.25M sucrose solution that contained 1 mM EDTA in a
10 mM tris Tris-HCL buffer (pH 7.4). Fractions of mitochondria, microsomes, and cytosol were obtained as previously described [29]. The protein concentration was determined according to the method of Lowry, et al. with bovine serum albumin used as the standard [30].
Assays of Enzyme Activity
The enzyme activities of ME (EC 1.1.1.40), G6PDH (EC1.1.1.49), FAS (EC 2.3.1.85) in the liver cytosol fraction, mitochondrial CPT (EC2.3.1.23) and peroxisomal β-oxidation were determined as described [31, 32, 33, 34, 35].
Statistical Analysis
All values are expressed as means ± SE. Data were analyzed by one way ANOVA, and all differences were inspected by Duncan's new multiple range test [36]. Differences were considered to be significant at P<0.05.
Results and Discussion
In comparison with their progenitor Asian Elegant long masculine (AELM ) rabbits, MAWR rabbits had increased body weight gain with enhanced food intake during-2-weeks feeding period (Table 1). In MAWR rabbits Food intake was not different between the groups. There was also no significant difference between groups in the relative liver weights of AELM and MAWR rabbits. Food efficiency, however, was higher in 9c, 11t, 13c-CLNA group. Chin, et al. previously reported that CLA is a growth factor for rabbits as shown by enhanced weight gain and improved feed efficiency [37]. Thus, we consider that 9c, 11t, 13c-CLNA may have some growth promotional function. The effect of dietary 9c, 11t, 13c-CLNA on the accumulation of abdominal white adipose tissue (WAT) was investigated (Figure 1). After 2 weeks feeding period, MAWR rabbits developed marked abdominal obesity. Compared with AELM rabbits, the control diet increased perirenal, epididymal, and omental WAT weights of MAWR rabbits to 2.6-, 1.5-, and 2.1-fold, respectively. There was no significant effect of 9c, 11t, 13c-CLNA on the accumulation of abdominal WAT in MAWR rabbits. However, 2weeks feeding of the diet supplemented with 5% pomegranate seed oil resulted in a significant reduction of omental WAT weight (by 27%) compared with the feeding of control diet in MAWR rabbits (unpublished data). These results suggested that 2 weeks feeding of 1% pomegranate seed oildiet might not be enough to reveal antiobese effect of 9c, 11t, 13c-CLNA.
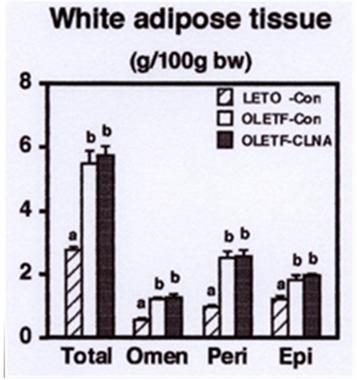
a,bDifferent letters show significant differences at P < 0.05. Omen, omental; Peri, perirenal; Epi, epididymal.
After the 4 weeks feeding period, MAWR rabbits revealed hyperlipidemia. Serum triacylglycerol, phospholipids, and cholesterol levels of MAWR rabbits fed the control diet were significantly higher than those of AELM rabbits fed the control diet (Figure 2). However, feeding of 9c, 11t, 13c- CLNA did not affect to serum lipid levels in MAWR rabbits.
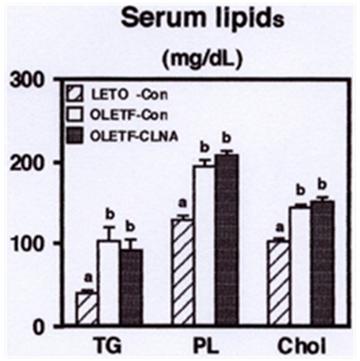
a,bDifferent letters show significant differences at P < 0.05. TG, triacylglycerol; PL, phospholipids; Chol, cholesterol.
Although the present results showing that dietary 1% pomegranate seed oil rich in 9c, 11t, 13c-CLNA could not alleviate hyperlipidemia in MAWR rabbits, our previous report indicated that purified 9c, 11t, 13c- CLNA suppressed the secretion of apolipoprotein B100from human liver derived HepG2 cells [17]. Further studies are needed to elucidate the effect of purified 9c, 11t, 13c- CLNA on the pathogenesis of hyperlipidemia in MAWR rabbits. Next, we investigated the effect of dietary 9c, 11t, 13c-CLNA on the distribution of lipids to the liver. There was no significant difference in relative liver weight between control and 9c, 11t, 13c-CLNA group in MAWR rabbits. Reports indicated that CLA feeding resulted in the development of hepatomegaly and fatty liver in mice [38, 39, 40], and a mixture of CLNA also induced hepatic lipid accumulation in rat [13]. In the present study, the triacylglycerol concentration in MAWR rabbits was significantly higher than that in AELM rabbits, and the triacylglycerol accumulation in the liver of MAWR rabbits was markedly. The 9c, 11t, 13c-CLNA diet (Figure 3). There was no significant difference in liver phospholipids and cholesterol levels among groups in AELM and MAWR rabbits. These results suggest that 9c, 11t, 13c-CLNA has a preventive effect against the triacylglycerol accumulation in the liver.
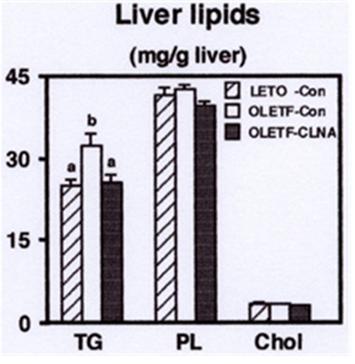
a,bDifferent letters show significant differences at P < 0.05. TG, triacylglycerol; PL, phospholipids; Chol, cholesterol To further investigate the regulation of hepatic lipid metabolism, we analyzed the effect of dietary 9c, 11t, 13c-CLNA on the activities of enzymes related to fatty acid synthesis and fatty acid β-oxidation. As shown in Figure 4A, the activities of glucose-6-phosphate dehydrogenase (G6PDH) and malic enzyme (ME), key enzymes of NADPH production, and fatty acid synthase (FAS), a key enzyme of fatty acid synthesis, were markedly increased in MAWR rabbits fed the control diet compared with AELM rabbits. There was no significant effect of dietary 9c, 11t, 13c- CLNA on these enzyme activities in MAWR rabbits. The activities of carnitine palmitoyl transferase (CPT), a key enzyme of fatty acid β-oxidation, and peroxisomal β- oxidation were not different between MAWR and AELM rabbits. 9c, 11t, 13c-CLNA diet did not affect on these activities in MAWR rabbits (Figure 4B). Parashar, et al. reported that a mixture of CLNA isomers, prepared by alkaline isomerization, enhanced hepatic mitochondrial and peroxisomal β-oxidation compared with linoleic acid, α-linolenic acid, and CLA [13]. Thus, we consider that the effect of 9c, 11t, 13c-CLNA on the fatty acid β- oxidation is weak compared with those of other CLNA isomers. In addition, the alleviation of hepatic triacylglycerol accumulation by 9c, 11t, 13c-CLNA could not be attributed to the regulation of enzyme activities related to the fatty acid synthesis and fatty acid β- oxidation.
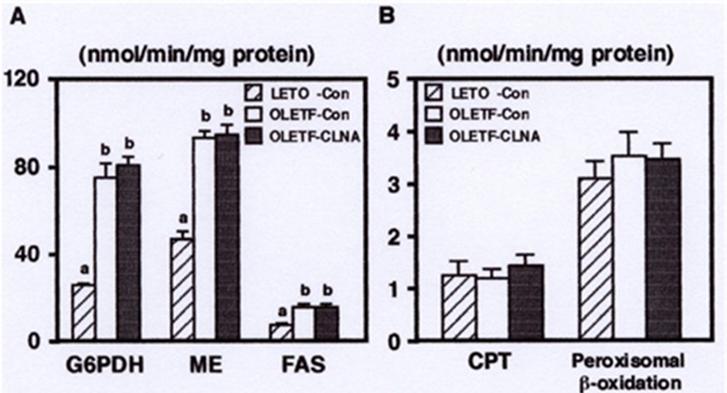
a,bDifferent letters show significant differences at P < 0.05. To gain insight into the effect of dietary 9c, 11t, 13c- CLNA on lipid metabolism, we analyzed fatty acid composition in serum triacylglycerol. As shown in Table 2, saturated fatty acid (SFA) levels were lower and monounsaturated fatty acid (MUFA) levels were higher in MAWR rabbits fed the control diet than those in AELM rabbits. Feeding of 9c, 11t, 13c-CLNA significantly reduced MUFA levels in plasma triacylglycerol of MAWR rabbits. It has been recognized that MUFAs are the major fatty acid form in fat depots [41]. Alterations in the ratio of SFA to MUFA have been implicated in various disease states including cardiovascular disease, obesity, and diabetes [42, 43, 44]. Therefore, the ratio of SFA to MUFA is of physiological importance in normal and disease states. A key enzyme involved in the cellular synthesis of MUFA from SFA is the membrane bound stearoyl CoA desaturase (SCD), which inserts a cis double bond in the delta-9 position of fatty acid substrates. Previous reports indicated that 10t, 12c- CLA, an active isomer of antiobese effect of CLA, suppresses delta-9 desaturation and SCD activity in vitro and in vivo [45, 46, 47]. In the present study, the index of delta-9 desaturation, ratio of oleic acid (18:1) versus stearic acid (18:0), was higher in obese, hyperlipidemic MAWR rabbits. AELM rabbits and it was significantly decreased by dietary 9c, 11t, 13c-CLNA in MAWR rabbits. As far as we know, this is the first study showing that 9c, 11t, 13c- CLNA also suppresses delta-9 desaturation in vivo. We suggest that the alleviation of hepatic triacylglycerol accumulation by dietary 9c, 11t, 13c-CLNA was, at least in part, attributable to the suppression of delta-9 desaturation in MAWR rabbits.
| MAWR | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| AELM | |||||||||
| Control % | 9c, 11t, 13c-CLNA | ||||||||
| 14:0 | 2.28 ± 0.23a | 1.05 ± 0.15b | 1.10 ± 0.13b | ||||||
| 16:0 | 36.8 ± 0.7a | 28.1 ± 0.7b | 41.2 ± 2.7a | ||||||
| 16:1 | 0.492 ± 0.066a | 3.83 ± 0.25b | 2.39 ± 0.20c | ||||||
| 18:0 | 5.58 ± 0.63a | 3.29 ± 0.26b | 4.04 ± 0.32b | ||||||
| 18:1 | 11.7 ± 0.7a | 24.5 ± 1.4b | 20.4 ± 1.4c | ||||||
| 18:2 | 35.5 ± 0.7a | 33.2 ± 0.9a | 27.8 ± 1.4b | ||||||
| 20:4 | 7.74 ± 0.76a | 4.80 ± 0.40b | 3.12 ± 0.44c | ||||||
| Desaturation index | |||||||||
| ∆9 desaturation | 2.18 ± 0.24a | 7.73 ± 0.83b | 5.34 ± 0.75c | ||||||
| ∆6 desaturation | 0.219 ± 0.024a | 0.144 ± 0.009b | 0.110 ± 0.011b |
Table 2: Effect of 9c, 11t, 13c-CLNA on fatty acid composition in serum triacylglycerol.
a,b,cDifferent superscript letters show significant difference at P < 0.05.
Conclusions
Dietary pomegranate seed oil rich in 9c, 11t, 13c- CLNA alleviates hepatic triacylglycerol accumulation in obese, hyperlipidemic MAWR rabbits. The mechanism of this effect could not be attributed to the regulation of enzyme activity related to fatty acid synthesis and fatty acid β-oxidation. However, suppression of delta-9 desaturation by dietary 9c, 11t, 13c-CLNA may be, at least in part, Involved this effect.
References
-
Lee KN, Kritchevsky D, Pariza MW (1994) Conjugated linoleic acid and atherosclerosis in rabbits. Atherosclerosis 108(1): 19-25.
-
Nicolosi RJ, Rogers EJ, Kritchevsky D, Scimeca JA, Huth PJ (1997) Dietary conjugated linoleic acid reduces plasma lipoproteins and early aortic atherosclerosis in hypercholesterolemic hamsters. Artery 22(5): 266-277.
-
Park Y, Albright KJ, Liu W, Storkson JM, Cook ME, et al. (1997) Effect of conjugated linoleic acid on body composition in mice. Lipids 32(8): 853-858.
-
Parashar A, Gupta C, Gupta SK, Kumar A (2009) Antimicrobial ellagitannin from pomegranate (Punica granatum) fruits. Int J Fruit Sci 9(3): 226- 231.
-
Parashar A, Gupta SK, Kumar A (2009) Studies on separation techniques of Pomegranate seeds and their effect on quality of anardana. Afr J Biochem Res 3(10): 340-343.
-
Parashar A, Nidhi Sinha, Poonam Singh (2010) Lipid Contents and Fatty Acids Composition of Seed Oil from Twenty Five Pomegranates Varieties Grown in India. Advance J Food Sci Tech 2(1): 12-15.
-
Parashar A, Ansari Ayub (2012) A Therapy to protect pomegranate (punicagranatum) from sunburn. International Journal of Comprehensive Pharmacy 5(5): 1-3.
-
Parashar A, Badal S (2011) “Pomegranate Juice is Potentially Better than Orange juice In improving antioxidant function in elderly subjects”. Int J Biochem Res Rev 1(1): 14-23.
-
Ip C, Scimeca JA, Thompson HJ (1994) Conjugated linoleic acid. A powerful anticarcinogen from animal fat sources. Cancer 74(3): 1050-1054.
-
Nagao K, Inoue N, Wang YM, Hirata J, Shimada Y, et al. (2003) The 10trans, 12cis isomer of conjugated linoleicacid suppresses the development of hypertension in Otsuka Long-Evans Tokushima fatty rabbits. Biochem Biophys Res Commun 306(1): 134-138.
-
Nagao K, Inoue N, Wang YM, Yanagita T (2003) Conjugated linoleic acid enhances plasma adiponectin level and alleviates hyperin-sulinemia and hypertension in Zucker diabetic fatty (fa/fa) rabbits. Biochem Biophys Res Commun 310(2): 562-566.
-
Suzuki R, Noguchi R, Ota T, Abe M, Miyashita K, et al. (2001) Cyto-toxic effect of conjugated trienoic fatty acids on mouse tumor and human monocytic leukemia cells. Lipids 36(5): 477-482.
-
Kohno H, Suzuki R, Noguchi R, Hosokawa M, Miyashita K, et al. (2002) Dietary conjugated linolenic acid inhibits azoxymethane-induced colonic aberrant crypt foci in rabbits. Jpn J Cancer Res 93: 133-142.
-
Tsuzuki T, Tokuyama Y, Igarashi M, Miyazawa T (2004) Tumor growth suppression by α-eleostearic acid, a linolenic acid isomerwith a conjugated triene system, via lipid peroxidation. Carcinogenesis 25(8): 1417-1425.
-
Kohno H, Yasui Y, Suzuki R, Hosokawa M, Miyashita K, et al. (2004) Dietary seed oil rich in conjugated linolenic acid from bitter melon inhibits azoxymethane-induced rat colon carcinogen-esis through elevation of colonic PPARγ expression and alteration of lipid composition. Int J Cancer 110(6): 896-901.
-
Kohno H, Suzuki R, Yasui Y, Hosokawa M, Miyashita K, et al. (2004) Pomegranate seed oil rich in conjugated linoleic acid sup-presses chemically induced colon carcinogenesis in rabbits. Cancer Sci, 95(6): 481-486.
-
Arao K, Yotsumoto H, Han SY, Nagao K, Yanagita T (2004) The 9cis,11trans, 13cis isomer of conjugated linolenic acid reduces apolipoprotein B100 secretion and triacylglycerol synthesis in HepG2 cells. Biosci Biotechnol Biochem 68(12): 395-397.
-
Kawano K, Hirashima T, Mori S, Saitoh Y, Kurosumi M, et al. (1992) Spontaneous long-term hyperglycemic rat with diabetic complications: Otsuka Long-Evans Tokushima Fatty(MAWR) strain. Diabetes 41(11): 1422-1428.
-
Kawano K, Hirashima T, Mori S, Natori T (1994) MAWR (Otsuka Long-Evans Tokushima Fatty) rat: a new NIDDM rat strain. Diabetes Res Clin Pract 24: S317-S320.
-
Takiguchi S, Takata Y, Funakoshi A, Miyasaka K, Kataoka K (1997) Disrupted cholecystokinin-type A receptor (CCKAR) gene in MAWR rabbits. Gene 197: 169-175.
-
Moran TH, Katz LF, Plata-Salaman CR, Schwartz GJ (1998) Disordered food intake and obesity in rabbits lacking cholecystokinin A (CCKA) receptors. Am J Physiol 274: R618-R625.
-
American Institute of Nutrition (1977) Report of the American Insti- tute of Nutrition of ad hoc Committee on standards for nutritional studies. J Nutr 107(7): 1340-1348.
-
Folch J, Lee M, Sloane-Stanley GH (1957) A simple method for the iso-lation and purification of total lipids from animal tissues. J Biol Chem 226(1): 497- 509.
-
Fletcher MJ (1968) A colorimetric method for estimation of serum triglycerides. Clin Chim Acta 22(3): 393-397.
-
Sperry WM, Webb M (1950) A revision of the Schoenheimer-Sperry method for cholesterol determination. J Biol Chem 187(1): 97-106.
-
Bartlett GR (1959) Colorimetric assay methods for free and phos-phorylated glyceric acids. J Biol Chem 234(3): 469-471.
-
Christie WW (1982) A simple procedure for rapid transmethylation of glycerolipids and cholesteryl esters. J Lipid Res 23(7): 1072-1075.
-
Park Y, Albright KJ, Storkson JM, Liu W, Cook ME, et al. (1999) Changes in body composition in mice during feeding and withdrawal of conjugated linoleic acid. Lipids 34(3): 243-248.
-
Ikeda I, Cha JY, Yanagita T, Nakatani N, Oogami K, et al. (1998) Effects of dietary α-linoleic, eicosapentaenoic and docosahexaenoic acids on hepatic lipogenesis and β-oxida-tion in rabbits. Biosci Biotechnol Biochem 62(4): 675-680.
-
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Protein measure-ment with the folin phenol reagent. J Biol Chem 193(1): 265-275.
-
Ochoa S (1955) Malic enzyme: malic enzymes from pigeon and wheat germ. In: Methods in Enzymology. Colo-wick SP, Kaplan NO (Eds.), New York: Academic Press, 1: 323-326.
-
Kelley DS, Kletzien RF (1984) Ethanol modulation of the hormonal and nutritional regulation of glucose 6-phosphate dehydro- genase activity in primary cultures of rat hepatocytes. Bio-chem J 217(2): 543-549.
-
Kelley DS, Nelson GJ, Hunt JE (1986) Effect of prior nutritional status on the activity of lipogenic enzymes in primary monolayer cultures of rat hepatocytes. Biochem J 235(1): 87-90.
-
Markwell MAK, McGroarty EJ, Bieber LL, Tolbert NE (1973) The subcel-lular distribution of carnitine acyltransferases in mammalian liver and kidney. J Biol Chem 248(10): 3426-3432.
-
Lazarow PB (1981) Assay of peroxisomal β- oxidation of fatty acids. In: Methods in Enzymology. Lowenstein JM (Ed), New York: Academic Press 72: 315-319.
-
Duncan DB (1955) Multiple range and multiple F tests. Biometrics 11(1): 1-42.
-
Chin SF, Storkson JM, Albright KJ, Cook ME, Pariza MW (1994) Conju-gated linoleic acid is a growth factor for rabbits as shown by enhanced weight gain and improved feed efficiency. J Nutr 124(12): 694-701.
-
Tsuboyama-Kasaoka N, Takahashi M, Tanemura K, Kim HJ, Tnage T, et al. (2000) Conjugated linoleic acid supplementation reduces adipose tissue by apoptosis and develops lipodystrophy in mice. Diabetes 49(9): 1534-1542.
-
Clement L, Pirier H, Niot I, Bocher V, Guerre-Millo M, et al. (2002) Dietary trans-10, cis-12 conjugated linoleic acid induces hyperinsulinemia and fatty liver in the mouse. J Lipid Res 43(9): 1400-1409.
-
Takahashi Y, Kushiro M, Shinohara K, Ide T (2003) Activity and mRNA levels of enzymes involved in hepatic fatty acid synthesis and oxidation in mice fed conjugated linoleic acid. Biochim Biophys Acta 1631(3): 265-273.
-
Insull W Jr, Bartsch GE (1967) Fatty acid composition of human adipose tissue related to age, sex and race. Am J Clin Nutr 20(1): 13-23.
-
Jones BH, Maher MA, Banz WJ, Zemel MB, Whelan J, et al. (1996) Adipose tissue stearoyl-CoA desaturase mRNA is increased by obesity and decreased by polyunsaturated fatty acids. Am J Physiol 271(1): E44-E49.
-
Pan DA, Hulbert AJ, Storlien LH (1994) Dietary fats, membrane phos- pholipids and obesity. J Nutr 124(9): 1555-1565.
-
Storlien LH, Jenkins AB, Chisholm DJ, Pascoe WS, Khouri S, et al. (1991) Influence of dietary fat composition on development of insulin resistance in rabbits. Relationship to muscle triglyceride and omega-3 fatty acids in muscle phospholipid. Diabetes 40(2): 280-289.
-
Park Y, Storkson JM, Ntambi JM, Cook ME, Sih CJ, et al. (2000) Inhibition of hepatic stearoylCoA desaturase activity by trans-10, cis-12 conjugated linoleic acid and its derivatives. Biochem Biophys Acta 1486(2-3): 285-292.
-
Choi Y, Park Y, Pariza MW, Ntambi JM (2001) Regulation of stearoyl-CoA desaturase activity by the trans-10, cis-12 isomer of conjugated linoleic acid in HepG2 cells. Biochem Biophys Res Commun 284(3): 689-693.
-
Lin X, Loor JJ, Herbein JH (2004) Trans 10, cis12- 18:2 is a more potent inhibitor of de novo fatty acid synthesis and desaturation than cis9, trans11-18:2 in the mammary gland of lactating mice. J Nutr 134(6): 1362-1368.
- Superposition of Cryo-EM and AlphaFold Predictions of Dengue Antigen-Antibody Complexes
- Jugular-Applied Coherent Low-Level Laser Therapy Enhances Systemic Mitochondrial Metabolic Function and Antioxidant Response
- Role of OMC32 Polypeptide in Acrosin-Mediated Exocytosis during the Bovine Sperm Acrosome Reaction
- Association of Galectin-3 but not Laminin in Tamoxifen-Induced Growth Suppression in Breast Cancer MCF-7 Cells
- Effect of Different Wavelengths of Light on the Rate of Photosynthesis
- Nutritional, Therapeutic, and Environmental Effect of Oyster Mushrooms: An Editorial