Evaluation of Green Fluorescent Protein Expression in Selected Indica Rice Varieties
Green Fluorescent Protein (GFP) expression in three indica rice genotypes PusaBasmati1, Swarna and Gayatri is evaluated in this study. GFP functions as reporter gene and the transformation efficiencies of these genotypes are compared. Results showed that PusaBasmati1, an aromatic variety showed higher transformation efficiency compared to Swarna and Gayatri which are popular varieties among farmers. Application of Green Fluorescent Protein (GFP) as reporter gene in rice has specific advantage over popularly employed β-glucuronidase (GUS) gene. Comparative expression studies of both these reporter genes were evaluated. Prospects and consequences of utilizing GFP as a reporter gene and the advantages of GFP gene over GUS gene were discussed.
Sai Krishna R*, Nirakar SNP, Chaitanya KG, and GJN Rao
Crop Improvement Division, ICAR-National Rice Research Institute, India
Rice Research Institute, Cuttack- 753006, Odisha, India, Email:
saikrishnarepalli@gmail.com reporter gene and the advantages of GFP gene over GUS gene were discussed.
Keywords: Rice; GFP; GUS; Reporter Gene; Transformation Efficiency Abbreviations: GFP: Green Fluorescent Protein; GOI: Gene of Interest; CI: Callus Induction.
Introduction
The transgenic approach of plant genetic engineering provides access to an unlimited gene pool for transfer and expression of desirable genes in plants between any two species irrespective of their evolutionary and taxonomic status. Rice gene pool is limited and cannot provide solution to all the biotic and abiotic stresses [1]. Conventional plant breeding methods have limitations in addressing the problems in particular situations where there are no donor parents available for crop improvement programs [2]. In this context, transgenic approach can provide solution by facilitating the transfer of resistant genes from any distant source to the desirable host. Since last two decades transfer of genes across the species is done through two important transformation methods including bacterial mediated Agrobacterium gene transfer [3] and particle mediated bombardment approach [4] which is physical method of gene transfer.
There are prospects and consequences in both these methods. Agrobacterium method is attributed to work well only in dicot systems but in the later years successful results were obtained in monocot systems as well [5]. There is a possibility of transfer of bacterial genes/vector back bone in this method where as biolistic method is purely physical method of gene transfer and here there is no interference of bacterial genes but the chance of integration of gene into the genome is less in comparison to Agrobacterium approach. There are possibilities of duplication of genes and silencing of genes due to the deposition of multiple sets of genes during biolistic approach [6].
Despite of the consequences of both the systems, during certain physiological studies more number of events are required to be generated within short span of time without waiting for the regeneration of plantlets. In such situations to study transient gene expression, biolistic method holds good. Moreover not all the transformants will turn into successful events, in such cases more number of transformants are to be generated and they are to be screened within short span of time; in such cases reporter genes does the apt function by reporting the presence of the gene of interest (GOI) as they would be flanked with GOI. There are several reporter genes in usage but this study focused on the popularly employed β-glucuronidase (GUS) gene [7] and Green Fluorescent Protein (GFP) as reporter genes in rice. The green fluorescent protein (GFP) from the jelly- fish Aequoria victoria has several significant advantages over alternative visual marker genes. Its expression can be detected in real time, in living cells and organisms simply by light excitation [8]. GFP does not require a substrate and is not toxic compared to firefly luciferase and the β- glucuronidase assay. The small size of GFP (26.9 kDa) facilitates the construction of protein fusions [9]. GFP allows monitoring of gene expression and protein localization at the sub-cellular, cellular and plant level.
In this study, efforts were made to evaluate the transformation efficiencies of three popular genotypes PusaBasmati1, Swarna and Gayatri. Sequence of steps followed and the prospects and consequences of GFP as reporter gene over GUS are discussed.
Materials and Methods
Genotypes
The rice genotypes selected for the study were three elite indica rice cultivars PusaBasmati1, Swarna and Gayatri. Pusa Basmati 1, is a Basmati type variety, which is extensively grown in the Basmati region of India with a yield potential of 4.5 t/ ha (Siddiq 1990). It is highly popular on account of its long slender grains, pleasant aroma and the cooked rice is endowed with desirable traits like soft texture and tenderness. Swarna, a widely grown indica rice variety in eleven states of India, is highly popular with a yield potential of 8.0 t/ha [10]. It is also being extensively grown in Bangladesh and Myanmar suggesting its wide adaptability [11]. Gayatri, a high yielding cultivar released from NRRI, is widely grown in shallow and medium low land ecology in Eastern India.
Callus Induction and Proliferation
Mature dehusked grains of the selected cultivars were washed with sterile distilled water and were surface sterilized successively with, 70% ethanol for two min, sodium hypochlorite (contains 4% (v/v) active chlorine) for 15 min and with 0.1% (w/v) aqueous mercuric chloride solution for 5min with intermittent repeated washings with sterile distilled water [12]. The kernels were inoculated in culture tubes containing semisolid callus induction (CI) medium [MS medium supplemented with 2, 4-dichlorophenoxy acetic acid (2, 4-D) (2 mgl-1), maltose (30 gl-1) and solidified with of gel-rite (2.6 gl-1)] [13] and the cultures were incubated in dark at 25 ± 1ºC for three weeks.
Plasmid Preparation
A single colony of E.coli strain DH5α/pCAMBIA1304 carrying a GUS-GFP fusion protein as reporter gene and hygromycin selectable marker gene, under control of 35S promoter was picked from a freshly streaked selection plate and inoculated into 5ml of LB medium [14] supplemented with kanamycin antibiotic (25 mgl-1) to initiate a starter culture. The starter culture was incubated for approximately 8h at 37°C with vigorous shaking (~ 220rpm). The starter culture was diluted (100 µl of starter culture was added to 100 ml LB medium supplemented with selective antibiotics) and the cells were grown at 37°C for 12-16h with vigorous shaking (~ 200 rpm). The bacterial cells were harvested by centrifugation at 6000 g for 15 min at 4°C. Further plasmid extraction steps were followed using Qiagen Plasmid extraction kit as per manufacturer’s instructions.
Explant Preparation & Transformation
Three week old scutellum derived embryogenic calli obtained from rice cultivars Pusa Basmati1 Swarna and Gayatri were taken as target materials for bombardment. Micro-carriers were prepared as per standard protocol using gold (1µ) particles. Embryogenic calli were bombarded at 1100 psi helium pressure with the transformation vector CAMBIA1304 (contaning GUS-GFP fusion protein and Hpt genes) using the particle gun PDC- 1000/He system (BIORAD) following manufacturer’s instructions. Selection of the transformed calli were performed as mentioned in our earlier work on media containing hygromycin antibiotic for four selection cycles for every fortnight [15]. Transformation efficiency is calculated by considering the number of calli fluoresced in UV light divided by total number of calli transformed and the whole multiplied by hundred.
Screening for GFP
The selected calli which had survived hygromycin selection were observed under hand held Ultra Violet (U.V) light in laminar chamber under aseptic conditions and calli which were fluoresced under UV light were taken for observations under confocal microscope.
Screening for GUS
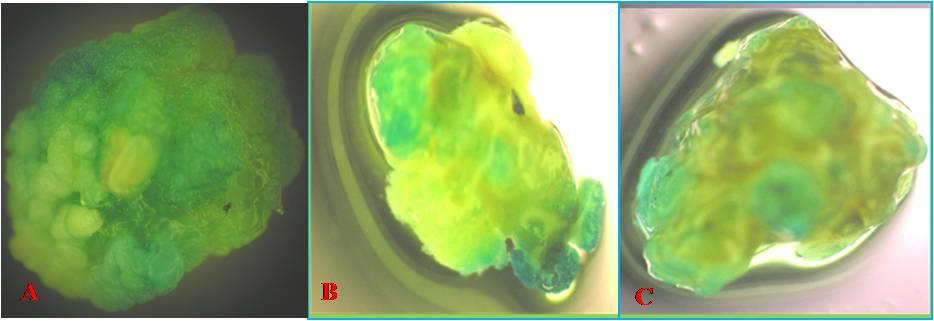
The calli which were fluoresced under UV light and observed under confocal microscope were immersed in GUS solution overnight and were observed for the blue color formation [16].
DNA Extraction and PCR Assay
Few selected calli which fluoresced in UV light were selected and the DNA extraction was done by using Qiagen DN easy mini plant extraction kit [17] as per the manufacturer’s instructions. The presence of GFP gene was examined by polymerase chain reaction (PCR) with the help of specific primers to give an amplification product of ~479 bp size. The plasmid DNA (pCAMBIA1304) was used as the positive control and non transformed callus DNA is taken as negative control. The PCR mix contained 1µl of plant DNA (20ng), 0.8µl of 2.5mM dNTPs (Fermentas), 1.0 µl of 10X PCR buffer (10mM Tris, pH 8.4, 50 mM KCl, and 15 mM MgCl2; Sigma), 0.2µl of Taq DNA Polymerase (5U/µl Sigma), 1 µl each of both forward and reverse primers (5 pico moles/µl Sigma) and 5 µl of autoclaved sterile distilled water in a total volume of 10 µl. The amplification was done in a thermal cycler (Eppendorf Vapo protect) under following conditions: an initial denaturation of template DNA at 94°C for 2 min followed by 40 cycles of amplification i.e., 1min denaturation at 940C, 1min primer annealing at 58°C, 1 min primer extension at 720C and 10 min final primer extension at 720C. Isolated DNA was also examined for the presence of HPT gene which was used as the antibiotic selectable marker as per the conditions similar to that of GFP gene amplification mentioned above but with the different PCR program as per the following conditions: an initial denaturation of template DNA at 94°C for 5 min followed by 40 cycles of amplification i.e., 1 min denaturation at 94°C, 1 min primer annealing at 58°C, 1 min primer extension at 72°C and 5 min final primer extension at 72°C. PCR products were separated in 1.2 % agarose gel (in 1X TBE electrophoresis buffer) containing 0.5 μg/ml ethidium bromide. Separated PCR products were visualized under UV light and photographed by gel documentation system (Alpha innotech) to examine the size of the product.
Results & Discussion
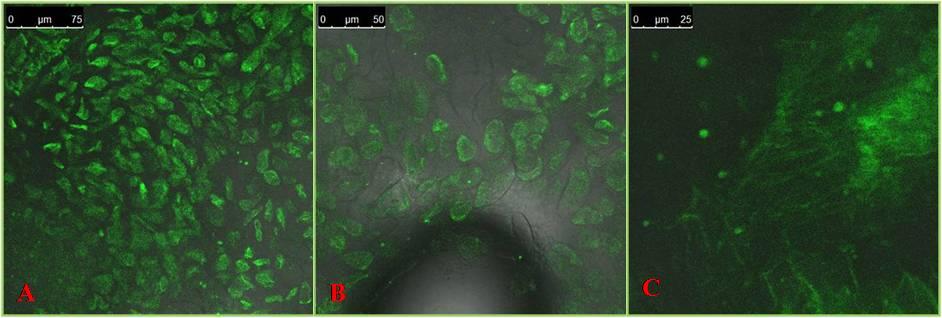
The expression pattern of the Green fluorescent protein (GFP), the well known reporter gene was studied in three indica rice genotypes i.e. Pusa Basmati1, Swarna and Gayatri. For this study, the plasmid vector pCAMBIA1304 carrying a GFP-GUS fusion protein and also the hygromycin gene, a selectable marker both under the control of 35S promoter was used and the transformation method employed was the particle gun bombardment i.e. biolistic approach and embryogenic calli were used as the recipients of the DNA.
Transformation rate of PusaBasmati1 (17.3 %) was found to be higher than Swarna (10.5 %) and Gayatri (6.6 %) which is evidenced through GFP expression studies. This shows that transformation efficiency varies from genotype to genotype (Table 1). Hence the variety with good transformation efficiency should be selected to transfer the genes of agronomic importance. Particularly these three varieties were chosen because of the best callus induction and regeneration potential of these cultivars [18].
The stable integration of the GFP and HPT genes was corroborated by PCR analysis. Four out of twenty samples taken randomly from an U.V fluoresced plate confirmed the presence of gene when observed in an agarose gel (Figure 3). Samples were compared with that of the positive plasmid control and the amplified gene product corresponded to the molecular size of plasmid control 479 bp in case of GFP gene and 450 bp in the case of HPT gene.
The images of the GFP expression were captured by a confocal microscope that was expressed in the emission range of 395nm to 475 nm, the ideal range for the GFP protein. The expression levels varied between genotypes suggesting the existence of genotypic differences for the expression levels of the GFP gene. In Pusa Basmati1, the level of expression was high followed by Swarna and Gayatri (Figure 1). As it is a fusion protein, the GUS expression was also good in the same calli (Figure 2). PCR analysis using gene specific primers of GFP did show the amplification of the target sequence suggesting the integration of GFP gene sequence in the rice genome.
With this demonstration of the incorporation and expression of GFP in rice, it is now possible to clone the gene of interest (GOI) adjacent to GFP reporter gene in the transformation vectors and use them for transformation purposes. The integration of the desired GOI in the target tissue can be indirectly confirmed by expression analysis of GFP through microscope as the presence of GFP gene is closely linked to GOI in the vector. Thus, use of GFP in rice transformation as a reporter gene has a distinct advantage over other reporter genes.
However, the study of GFP expression requires the use of advanced fluorescent microscope, as essential equipment. There are also apprehensions regarding the effects of ultra violet radiation (the fluorescing range of GFP) on the calli during the cytological assays as U.V radiations may induce mutations in the calli exposed to the radiation. The existences of genotypic differences in the transformation efficiency do indicate that choice of cultivar can also be an important factor to attain higher levels of transformation.
Despite of these drawbacks, GFP has a clear advantage over GUS as the cytological assay of GFP is a non destructive assay and the callus after visualization under the microscope can be used for regeneration while in the case of GUS, as the GUS assay is destructive, the callus cannot be used for regeneration after the assay [19]. Hence, GFP would be a better choice as a reporter gene if the working conditions are suitably developed to view the GFP expression in the callus.
- Name of the variety
- Number of calli transformed
- (%)
- PusaBasmati1
- 450
- 78
- 17.3
- Swarna
- 228
- 24
- 10.5
- Gayatri
- 270
- 18
- 6.6
Table 1: Transformation efficiencies of three varieties.
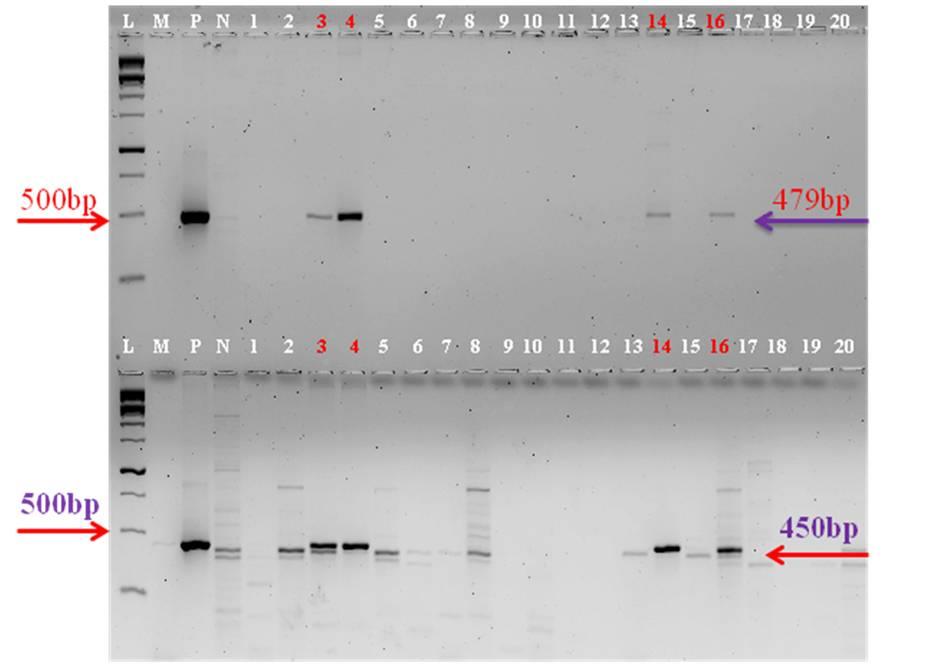
L-1 kb ladder, M-PCR Master mix, P-Positive control, N-Negative control, Samples (1-20) GFP amplification upper panel; HPT amplification lower panel Figure 3: PCR amplification of HPT and GFP genes.
Conclusion
Expression of GFP as reporter gene in three indica rice varieties is evaluated. Transformation efficiencies of the three genotypes are compared based on the reporter gene expression. Transient reporter gene expression has advantages as it saves time to study the transfer of gene instead of waiting until regeneration of plantlets. Gene of Interest is flanked to the reporter gene and controlled by a common promoter hence reporter gene expression depicts the integration of gene of interest as well. GUS assay is destructive and the calli used for GUS expression cannot regenerate, hence GFP overcomes these limitations and is preferred as best reporter gene given the facilities for confocal microscope is made available.
Acknowledgements
The authors are thankful to Director, NRRI for the facilities and encouragement. The authors express their sincere thanks to Prof. M.V. Subba Rao, Andhra University for providing pCAMBIA plasmid for their work at NRRI. The authors express their gratitude to Prof. S.C. Sabat, and Director, Institute of Life Sciences, Bhubaneswar for allowing them to utilize confocal microscope facility at their institute. Authors, Sai Krishna Repalli and Chaitanya Kumar Geda are also thankful to ICAR-NPTC for providing them Senior Research Fellowship.
References
-
Bonman JM, Khush GS, Nelson RJ (1992) Breeding Rice for Resistance to Pests. Annu Rev Phytopathology 30: 507-528.
-
Borojevic S (1990) Principles and methods of plant breeding. Elsevier, New York.
-
Hiei Y, Ohta S, Komari T, Kumashiro T (1994) Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant Journal 6(2): 271-282.
-
Christou P, Yang NS (1994) Applications to Plants in Particle bombardment technology for gene transfer. Oxford University Press, New York.
-
Wu YY, Chen QJ, Cui XH, Chen H, Chen J, et al. (2007) Efficient regeneration and Agrobacterium-mediated stable transformation of perennial ryegrass. Russian Journal Plant Physiology 54(4): 524-529.
-
Spangenberg G, Wang ZY, Wu XL, Nagel J, Potrykus I (1995) Transgenic perennial ryegrass (Lolium perenne) plants from microprojectile bombardment of embryogenic suspension cells. Plant Sci 108(2): 209-217.
-
Jefferson RA, Kacanagh TA, Bevan MW (1986) β- glucouronidase from Escherichia coli as a gene-fusion marker. Proc Natl Acad Sci USA 83(22): 8447-8451.
-
Cubitt AB, Heim R, Adams SR, Boyd AE, Gross LA, et al. (1995) Understanding, improving and using green fluorescent proteins. Trends Biochem Sci 20(11): 448-455.
-
Wang S, Hazelrigg T (1994) Implication for bcd mRNA localization from spatial distribution of exu protein in Drosophila oogenesis. Nature 369(6479): 400-403
-
Rao VR, Reddy PS, Murthy N, Rao I, Rao PS, et al. (1983) Swarna (MTU 7029)–a new stable hybrid with wide adaptation. Oryza 20: 240-242.
-
Baisakh N, Datta K, Oliva N, Ona I, Rao GJN, et al. (2001) Rapid development of homozygous transgenic rice using anther culture harboring rice chitinase gene for enhanced sheath blight resistance. Plant Biotech J 18(2): 101-108.
-
Vijayachandra K, Palanichelvam K, Velthambi K (1995) Rice scutellum induces Agrobacterium tumefaciens vir genes and T-strand generation. Plant Mol Biol 29(1): 125-133
-
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassay with tobacco tissue cultures. Physiologla Plantarum 15: 473-497.
-
Bertani G (1951) Studies on lysogenesis. I. The mode of phage liberation by lysogenic Escherichia coli. J Bacteriol 62(3): 293-300.
-
Saikrishna R, Chaitanya KG, Rao GJN (2013) Efficacy of different transformation methods in rice (Oryza sativa L.). JEBAS 1.
-
Rueb S, Hensgens LAM (1989) Improved histochemical staining for β-D-glucuronidase activity in monocotyledonous plants. Rice Genetics Newsletter 6: 168-169.
-
Yan H, Rommens CM (2007) Transposition-based plant transformation. Plant Physiol 143(2): 570-578.
-
Chaitanya KG, Sai RK, Mohan TD, GJN Rao (2013) Genetic variation in in vitro response of elite aromatic and non aromatic rice varieties. Oryza 50(4): 329- 333.
-
Birch RG, Bower R (1994) Principles of gene transfer using particle bombardment. In: Yang SN, Christou P (Eds.), Particle bombardment technology for gene transfer, Oxford University Press, New York, pp: 3-37.
- Superposition of Cryo-EM and AlphaFold Predictions of Dengue Antigen-Antibody Complexes
- Jugular-Applied Coherent Low-Level Laser Therapy Enhances Systemic Mitochondrial Metabolic Function and Antioxidant Response
- Role of OMC32 Polypeptide in Acrosin-Mediated Exocytosis during the Bovine Sperm Acrosome Reaction
- Association of Galectin-3 but not Laminin in Tamoxifen-Induced Growth Suppression in Breast Cancer MCF-7 Cells
- Effect of Different Wavelengths of Light on the Rate of Photosynthesis
- Nutritional, Therapeutic, and Environmental Effect of Oyster Mushrooms: An Editorial