Effect of Zinc Ions on the Proliferation and Differentiation of Keratinocytes
Background: Psoriasis is a chronic inflammatory disease in which there is hyperproliferation and abnormal differentiation of keratinocytes. Since high levels of KLK7, an enzyme inhibited by zinc (Zn2+) ions, are present in psoriatic lesions, we have studied the effect of zinc ions in the viability of keratinocytes, as well as in the activity of KLK5 and KLK7 and in the expression of epidermal markers. Methods and Results: The cells were cultured in the absence or presence of Zn2+ ions (5.0, 10 and 25 M). Cell viability was evaluated by the MTT method after during 14 days. Cell death was evaluated by flow cytometry using propidium iodide. The activity of the KLK was evaluated on the hydrolysis of synthetic substrates. Expression of involucrin, filaggrin, cytokeratins (CK) 5, 10 and 14 was evaluated by quantitative PCR. Cell incubation with Zn2+ ions did not result in significant changes in cell viability. By MTT assay, it was observed that the cultures incubated with 10 and 25 M Zn2+ ions showed a decrease in the number of viable cells in comparison to the control. Cells cultured for 1 day in the presence of 25 M Zn2+ ions displayed a decrease in KLK7 activity. In the presence of Zn2+ ions, it was shown an increase in the expression of CK5, 10 and 14, involucrin and filaggrin. Conclusions: These results have shown that zinc ions can affect the differentiation of HaCat cells, contributing for future therapeutic trials related to psoriasis based on the modulation of KLK activity.
Introduction
Psoriasis is a chronic inflammatory disease in which there is hyperproliferation and abnormal differentiation of epidermal keratinocytes, characterized by granulosa layer disappearance, dermal angiogenesis and infiltration of inflammatory cells [1, 2]. These modifications can be a) structural, such as an increase in cell and nuclear mass, b) ultrastructural, such as an increase in the intercellular space and the presence of cytoplasmic and nuclear remnants in the stratum corneum, or c) functional, such as the accelerated transit of epidermal cells. In addition to epidermal hyperproliferation, there is a reduction of up to eight times in the keratinocyte cycle, which, consequently, results in incomplete cell maturation. Additionally, a decrease in lipid synthesis occurs, which consequently affects the formation of desmosomes [3, 4].
The stimulatory cytokines IL-1β, IL-6, interferon gamma (IFN-γ), as well as the presence of T cells, damage the plasma membrane of keratinocytes and participate in the molecular mechanisms involved in the epidermal hyperproliferation observed in psoriasis. Additionally, cytokines released by T cells stimulate IL-8 synthesis by keratinocytes and, as a result, there is recruitment of neutrophils, which increases cell damage [5, 6]. In psoriasis, IL-1β also regulates the expression of other cytokines and contributes to T cell extravasation. In vitro studies have shown that during the inflammatory process of psoriasis, an increase in the amount of the epidermal kallikrein 7 (KLK7) is observed, which, in turn, is capable of cleaving the precursor form of interleukin 1β (IL-1β) [6, 7].
The KLK7 is part of a family of enzymes that has been associated to the physiologic desquamation and skin renewal. KLK7 and KLK5 are the most active kallikreins [8] involved in epidermis homeostasis, due to their action such as primary regulators of the KLK cascade activation, in the deeper layers of stratum corneum.
Although its action is quite pronounced in this layer, its active form is significantly less abundant than the inactive one, and two factors contribute to the regulation of its activity: the presence of Zn2+ ions, which is an allosteric inhibitor of these enzymes [9] and the endogenous lymphoepithelial inhibitor of serine proteases of the Kazal type 5 (LEKTI) encoded by the SPINK 5 gene [10], which associates with the enzyme at neutral pH (Park et al., 2022). As cells migrate to the more superficial layers, the pH of the cellular environment becomes acid, resulting in the dissociation of KLK from LEKTI [11], in addition to a decrease in the concentration of Zn2+ ions.
Regarding KLK7, studies have demonstrated a high presence of this enzyme in its active form in the skin of patients with atopic dermatitis and psoriasis. These data suggest the involvement of KLK7 in the pathogenesis of psoriasis, especially in skin desquamation and in the underlying inflammatory process [12, 13].
Thus, considering that KLK5 and KLK7 are crucial for the differentiation of keratinocytes and for desquamation of epidermis, in addition to the fact that KLK7, an enzyme strongly inhibited by Zn2+ ions [14], is involved in IL-1β activation, we studied whether the kallikrein enzymes inhibition by Zn2+ ions could affect the proliferation and differentiation of HaCaT epidermal lineage cells, as well as investigating the effect of Zn2+ ions in different concentrations, on the enzymatic activity of KLK5 and KLK7 and on the expression of epidermal differentiation markers. This work may provide useful tools, both for the study of skin biology, and contribute to therapeutic strategies for patients with psoriasis diagnosis, based on the production of formulations containing Zn2+ ions, which control the hyperproliferation of keratinocytes while do not affect the epidermis renewal dynamics.
Material and Methods
Cell Cultures and Treatment
The cells used in this project are part of the cell bank in the Skin Physiology and Tissue Bioengineering laboratory (LAFIPE-Bio) at the School of Arts, Science and Humanities of the University of São Paulo (EACH-USP). The HaCaT cells, a non-tumorigenic, spontaneously transformed keratinocyte lineage [15] were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM) (Invitrogen) containing 10% fetal bovine serum (FBS) (Invitrogen) and 1.8 mM CaCl2, in addition to the antibiotics 10,000 U/l penicillin and 100 mg/l streptomycin (Invitrogen), in a humid atmosphere with 5% CO2 at 37°C until the culture reach 80% of confluence. For the experiments, cells were plated at a concentration of 2 x 104 cells/well, in 6-well plates, or at 1 x 103 cells/well, in 96-well plates. Cultivation occurred for different periods of time in the absence or presence of Zn2+ ions (5.0; 10; 25; 50 or 100 μM) in the form of ZnCl2 or Zn(NO3)2 (BDH Chemicals Ltd, Pool, England). The images of the cultures were captured in a trinocular inverted microscope model – TS100F (Nikon).
ZnCl2 and Zn(NO3)2 Solutions Preparation
ZnCl2 and Zn(NO3)2 solutions were prepared at a stock concentration of 200 mM (Sigma-Aldrich) and filtered in a 0.22 μm pore filter. At the moment of treating cells, ZnCl2 and Zn(NO3)2 stock solutions were diluted to 5.0, 10 and 25 μM in the culture medium.
Cell Viability Analysis
Flow Cytometry
The cells were distributed in 6-well plates at a concentration of 2 x 104 cells/well and incubated for 48 h with different concentrations of Zn2+ ions (5.0; 10; 25; 50 and 100 μM). The culture medium was replaced every 48 h. After the treatments, the plates were washed with 1 ml of sterile phosphate buffer saline (PBS) and the cells were collected with 0.25% trypsin. The cell suspension was centrifuged for 7 minutes, at 400 x g. To assess cell viability, pellet was resuspended into 500 µl of PBS and 5 μl of 1 mg/ ml propidium iodide (PI) was added to the cell suspension according to Riccardi and Nicoletti (2006) [16]. Cells were evaluated on Guava EasyCyte 8HT flow cytometer using the InCyte® software in three individual experiments in triplicates. 10.000 events were acquired per sample. The results were expressed as a percentage of the total number of cells in each sample.
MTT Method
The 3-(4,5-dimethylazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) stock solution was prepared at 5 mg/ml in PBS. After the treatments with ZnCl2 and Zn(NO3)2 solutions, for 1, 3, 7, 9 and 14 days, 10 μl of MTT solution were added to the cultures in 96-well microplate and the cells were cultured for 4 h, at 37ºC. After this period, the medium was removed and 100 μl of dimethyl sulfoxide (DMSO) was added to each well to dissolve formazan crystals. Cell viability was determined by absorbance at 570 nm wavelength, with BioTek Synergy HT microplate reader. Two individual experiments were performed in sextuplicates and the results were analyzed with Gen5 software. Although this assay is known to be not suitable for determining cell proliferation, since the amount of viable (and dead) cells in each concentration was determined by flow cytometry it was possible to access cell proliferation.
Determination of Protein Concentration in Cell Extracts
The amount of protein in each sample was determined using the method that employs the Bradford reagent [17]. 20 µl of each sample from cell lysates were used, which were mixed with 200 µl of Bio Rad Protein Assay reagent, prepared according to the manufacturer’s instructions. After 5 min of reaction, the absorbance of the solutions was determined at 595 nm, in the Synergy HT plate reader using the Gen5™ software (BioTek Instruments). The protein concentration of the samples, in duplicates, was extrapolated from the absorbance values, using a Bovine Serum Albumin (BSA) curve as a standard.
Detection of the Enzymatic Activity of the Epidermal Kallikreins KLK5 and KLK7
For the enzymatic activity assays, cells cultured for 1, 3 or 5 days in the presence of Zn2+ ions in the form of ZnCl2 (5.0; 10 and 25 µM) were lysed with 400 µl of the buffer 25 mM HEPES, pH 7.5, containing 0.5% Triton X-100, to obtain cell extracts. The amount of protein present in the extracts was determined using the Bradford method (1976). The activity of KLK5 and KLK7 was verified in the hydrolysis of the fluorogenic substrates (FRET) Abz-GFSPFRSSRQ- EDDnp and Abz-KLYSSKQ-EDDnp, respectively, in the Synergy HT plate reader (BioTek), according to Fernandes, et al. (2022) [18]. Briefly, 40 µl of sample were used with 50 µl of 2x concentrated enzymatic reaction buffer (100 mM Tris, pH 8.0, 300 mM NaCl and 0.01% Tween 20) and 10 µl of substrate (100 µM) in a final volume of 100 µl. Mean rate results for each reaction were obtained using Gen5™ software (BioTek). Enzymatic activity was measured in arbitrary fluorescence units (UAF) per minute and expressed in terms of the average reaction rate per 1 µg of protein present in the extracts.
Inhibition of KLK5 and KLK7 Activity by Zinc
To evaluate the inhibition of KLK5 and KLK7 by Zn2+, experiments were carried out using 20 µl of samples (cell extracts from control samples), 20 µl of ZnCl2 solution at different concentrations (1.0; 2.5; 5.0; 10; 25 and 50 µM), 50 µl of reaction buffer and 10 µl of 100 µM substrate solution (Abz-GFSPFRSSRQ-EDDnp and KLYSSKQ-EDDnp to assess KLK5 and KLK7 inhibition, respectively), in a final volume of 100 µl. The hydrolysis of the substrates was accompanied as described above.
Real-time quantitative PCR (qPCR)
For total RNA extraction, cells were incubated with Zn2+ ions (0; 5.0; 10 and 25 µM), for 1, 3 or 5 days. Cultures were washed with ice-cold PBS, cells were collected by trypsinization (0.25%), centrifuged for 7 min at 400 x g and then 300 µl of Trizol (ThermoFisher Scientific) were added. Samples were stored at -80ºC for at least 24 h. After this period, they were thawed in the Eppendorf Thermomixer R bath (Eppendorf) for 5 min at 30°C. Then, 60 µl of chloroform were added and the samples incubated for 2 min at 30ºC. The aqueous phase of this solution was transferred to a sterile tube where 300 µl of isopropyl alcohol were added. The samples were incubated for 10 min at 30ºC, centrifuged at 12000 x g for 10 min, resuspended in 600 µl of 70% ethanol and stored at -80ºC for 24 h. After freezing, they were centrifuged at 7500 x g for 15 min, the supernatant discarded and the pellet resuspended in 15 µl of sterile ultra-pure water. The amount and integrity of the extracted RNA were evaluated at 260 and 280 nm in a spectrophotometer (NanoDrop, ThermoFisher Scientific). The cDNA strand was synthesized using the SuperScript® III First-Strand Synthesis System for RT-PCR kit, according to the manufacturer›s instructions (ThermoFisher Scientific). Expression of the genes that encode to the proteins CK5, CK10, CK14, involucrin and filaggrin was assessed by quantitative PCR (qPCR) using SYBR® green PCR Master Mix reagent (ThermoFisher Scientific), according to the manufacturer’s instructions. The oligonucleotides were designed, according to Table 1, in order to avoid non- specific annealing and following the recommendations for use in quantitative PCR.
| Oligonucleotides | Sequence | Reference |
|---|---|---|
| CK5 | Forward: CTTGTGGAGTGGGTGGCTAT | Nickerson et al. (2009) |
| Reverse: CCACTTGGTGTCCAGAACCT | ||
| CK10 | Forward: CAACCTAACAACTGATAATGCC | Santos et al. (2019) |
| Reverse: GTCTTTCATTTCCTCCTCGT | ||
| CK14 | Forward: GGAACAAGATTCTCACAGCC | Santos et al. (2019) |
| Reverse: TCCATCTCCACATTGACATCTC | ||
| Involucrin | Forward: AATGAAACAGCCAACTCCACTGCC | Fernandes et al. (2022) |
| Reverse: TCTTGCTTTGATGGGACCTCCACT | ||
| Filaggrin | Forward: TGAAGCCTATGACACCACTGA | Wang et al. (2017) |
| Reverse: TCCCCTACGCTTTCTTGTCCT |
Table 1: Oligonucleotides used for epidermal markers expression evaluation.
Expression of the genes of interest was normalized by expression of the glyceraldehyde 3-phosphate dehydrogenase (GAPDH) gene. The reactions were performed on the Eco™ Real-Time PCR System (Illumina). The data obtained were analyzed using Eco™ Software version 4.1 (Illumina). The experiments were performed in duplicates and data presented as mean ± standard deviation (SD). For data analysis, the 2-ΔΔCt method [19] was used.
Western Blotting
Cells cultured in the presence of different concentrations of Zn2+ ions (0; 5.0; 10 and 25 µM), for 1, 3 or 5 days, were lysed with 100 µl of 2-[4-(2-hydroxyethyl 25 mM)-1-piperazinyl]- ethanesulfonic acid (HEPES), pH 7.5, 0.5% Triton X-100, 2 mM ethylenediaminetetraacetic acid (EDTA), and Complete™ cocktail of protease inhibitors (Sigma Aldrich) for 20 min. Cell extracts were stored at -80°C. After the first thawing, the samples were centrifuged at 4°C, 10000 x g for 10 min. To determine the amount of protein present in cellular extracts, Bradford’s method (1976) was used. Aliquots with 25 µg of protein were used per sample. Samples were diluted in NuPAGE® LDS Sample Buffer (ThermoFisher Scientific), in the presence of NuPAGE® Sample Reducing Agent (ThermoFisher Scientific) and heated at 100°C for 7 min. Then, they were applied to NuPAGE® Novex® 4-12% Bis-Tris polyacrylamide gels (ThermoFisher Scientific). The electrophoretic run was carried out in NuPAGE® MOPS SDS Running Buffer (ThermoFisher Scientific) for approximately 1 h at 100 V in the Mini Gel Tank device (ThermoFisher Scientific).
After separation, the proteins in the gel were transferred to a nitrocellulose membrane for 7 min in the iBlot™2 Dry Blotting System (ThermoFisher Scientific) and nonspecific sites on the membrane were blocked with blocking buffer ( 10 mM Tris, pH 7.5, 150 mM NaCl, 0.10% Tween 20 and 5.0% skimmed milk) under stirring for 1 h at room temperature (RT). After blocking ended, the membrane was washed three times, for 10 min, with the washing buffer (10 mM Tris, pH 7.5, 150 mM NaCl, 0.10% Tween 20) under stirring.
Subsequently, the membrane was incubated overnight at 4°C with one of the primary antibodies anti-involucrin (0.75 µg/ml; ThermoFisher) and anti-β-actin (1:2000; Abcam) diluted in blocking buffer. After incubation with the primary antibody (from mouse), the membrane was washed three times, for 10 min, under agitation. Secondary anti-mouse IgG antibody conjugated with Horseradish Peroxidase (HRP) was then added, diluted 1:1000 in the blocking buffer. The membrane was incubated with the secondary antibody for 1 h at RT, under agitation. After this period, the membrane was washed three times for 10 min.
Band detection was performed by chemiluminescence using the Pierce™ ECL Western Blot Immunodetection kit (ThermoFisher Scientific), after exposing the membranes on the Amersham Imager 600 (GE Healthcare Life Sciences), for approximately 1 min. The bands were analyzed by densitometry, using the CPAtlas 2.0 Build 1 (2009) program. The results were expressed considering the amount of the protein of interest in relation to the constitutive protein (β-actin).
Statistical Analysis
Data were expressed as mean ± standard deviation of different experiments, as indicated in each case, and performed in, at least, duplicates. Afterwards, they were submitted to a two- way analysis of variance (two-way ANOVA), followed by the Bonferroni post-test for multiple comparisons, using the software GraphPad Prism version 5 (GraphPad Software Inc.). Differences were considered statistically significant for ρ<0.05 or ρ<0.001, as indicated.
Results
Cell Cultures
In the presence of different concentrations of Zn2+ ions, no visible alterations were verified in the morphology of the cells, which exhibited a cuboidal shape and were juxtaposed, which is characteristic of lining epithelial cells. Furthermore, the cells formed typical colonies of keratinocytes in culture (Figures 1A - D).
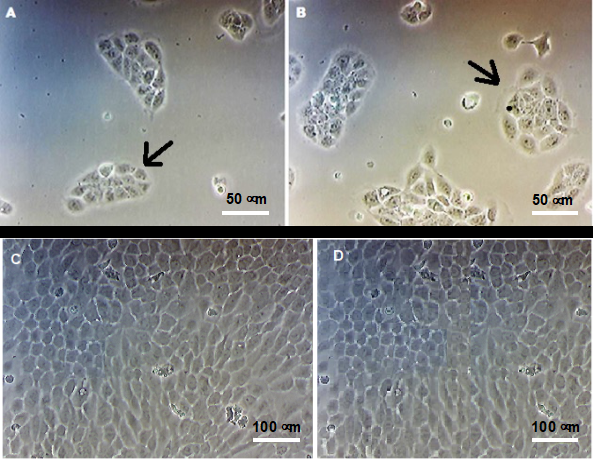
Figure 1: Morphology of HaCaT cells evaluated by optical microscopy. HaCaT keratinocytes were cultured in the absence (A and C) or in the presence of 25 µM ZnCl2 (B and D), for 24 h (A and C) and 5 days (C and D). The black arrows indicate the characteristic colonies of keratinocytes in culture. No phenotypic differences were observed between cultures in the presence of zinc ions. Magnification of 400 x (A and B) and 100 x (C and D).
Evaluation of Cell Viability and Death by Flow Cytometry
Incubation of cells with Zn2+ ions at the tested concentrations (0 to 100 µM), for 48 h, either in the form of ZnCl2 or Zn(NO3)2, did not result in significant changes in cell viability compared to control, suggesting that this ion is not toxic to cells (Figure 2).
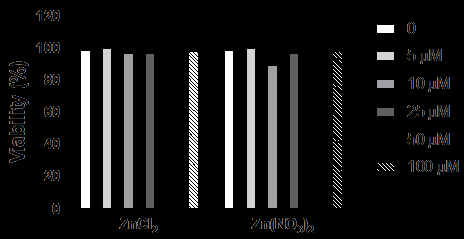
Figure 2: Viability analysis by flow cytometry of HaCaT cells cultured in the absence or presence of Zn2+ ions. HaCaT cells were incubated with Zn2+ ions (5.0; 10; 25; 50 and 100 μM) for 48 h. Cell viability was evaluated by flow cytometry using propidium iodide. Values represent mean ± standard error of the mean of three individual experiments performed in duplicates and are expressed relative to control (cells cultured in the absence of Zn2+). 10000 events were acquired per sample.
Evaluation of Cell Viability by MTT Method
Regarding cell viability, in general, the cultures grown in the presence of ZnCl2 or Zn(NO3)2 (5.0; 10 and 25 μM) showed a decrease in the number of viable cells, from the seventh day, in comparison to control of the same culture period (Figures 3A and 3B). As such concentrations did not cause cell death, as assessed by flow cytometry, it could be suggested that there was a decrease in cell proliferation in the analyzed periods.
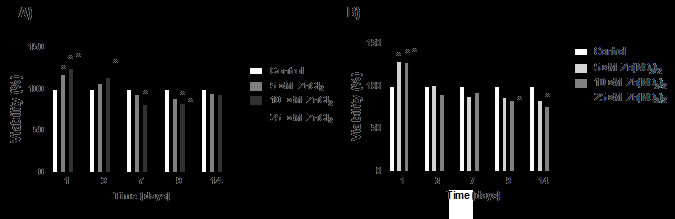
Figure 3: Viability analysis by MTT method of HaCaT cells cultured in the absence or presence of Zn2+ ions. HaCaT cells viability was evaluated in different periods of culture (1,3,7,9 and 14 days), in the presence of 5.0,10 and 25 µM (A) ZnCl2 or (B) Zn(NO3)2, using the MTT method. The results were expressed as the mean ± standard error of two different experiments in sextuplicates and analyzed by ANOVA + Bonferroni post-test. *, different compared to control (ρ<0.001).
Detection of the Enzymatic Activity of the Epidermal Kallikreins KLK5 and KLK7
Considering that the cell viability results were very similar between the ZnCl2 or Zn(NO3)2, for the following experiments, we only used ZnCl2 as a source of zinc ions.
Regarding KLK5, there was no difference in its activity in cells cultivated for 1 or 5 days, in the presence of all ZnCl2 concentrations tested, in relation to the control (Figure 4A). For KLK7, there was a decrease of around 30% in its activity in cultures incubated for 1 day with 25 μM Zn2+ ions compared to cells cultured for the same period in the absence of these ions (Figure 4B).
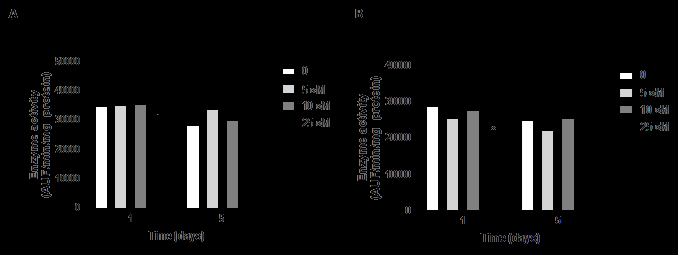
Figure 4: KLK5 and KLK7 activity in cells cultured in the absence or presence of ZnCl2. The activity of KLK5 (A) and KLK7 (B) was determined in the hydrolysis of the substrates Abz-GFSPFRSSRQ-EDDnp and Abz-KLYSSKQ-EDDnp, respectively, and evaluated at λex=320 nm and λem=420 nm. Extracts from cell cultures grown for 1 or 5 days in the presence of different concentrations of Zn2+ ions were used. AUF: arbitrary unit of fluorescence. Differences were analyzed by ANOVA + Bonferroni post-test. *different in comparison to control in the same cultivation period (ρ<0.001).
Inhibition of KLK Activity by Zinc Ions
Both KLK5 (Figure 5A) and KLK7 (Figure 5B) activity were reduced in the presence of different concentrations of Zn2+ ions. Although these results do not allow us to define the concentrations to be used in cellular assays, as this extrapolation is related to many other factors, they confirm the presence of these enzymes in the samples, as well as their inhibition by this cation. Specifically, Zn2+ ions at a final concentration of 50 μM reduced, by about 70%, the activity present in the extracts on the substrate Abz-GFSPFRSSRQ- EDDnp, used to investigate the activity of KLK5; for KLK7, this inhibition was 86% on the substrate Abz-KLYSSKQ- EDDnp.
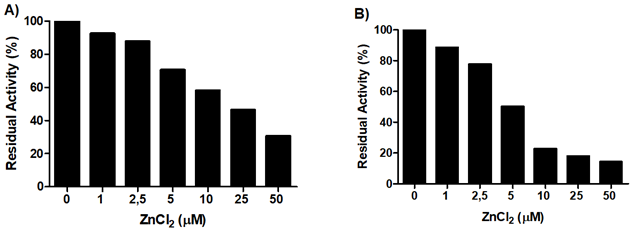
Figure 5: Inhibition of KLK5 and KLK7 by Zn2+ ions. The activity of (A) KLK5 and (B) KLK7 was measured in the presence of different concentrations of Zn2+ ions in the hydrolysis of the substrates Abz-GFSPFRSSRQ-EDDnp and Abz-KLYSSKQ-EDDnp, respectively, and evaluated at λex=320 nm and λem= 420 nm. Extracts from cell cultures grown for 1 day in the absence of Zn2+ were used. The results are expressed, in percentage, in terms of the residual activity of the enzymes, in the presence of different Zn2+ ions concentrations in comparison to control.
Analysis of the Expression of Proteins Involved in the Differentiation of Keratinocytes by Real Time Quantitative PCR
Considering the proteins CK5 (Figure 6A) and CK14 (Figure 6B), it was found that their expression was 6 to 10 times and 4 to 6, respectively, higher on day 1 and 3 in cells cultured in the presence of 5 µM Zn2+ ions when compared to the control (cells cultivated in the absence of zinc ions for one day). The CK10 expression was around 25 fold higher on day 3 of cultivation with 5 µM Zn2+ ions in comparison to the control, and about 5 times compared to cultures cultivated with 5 µM Zn2+ ions for only one day (Figure 6C).
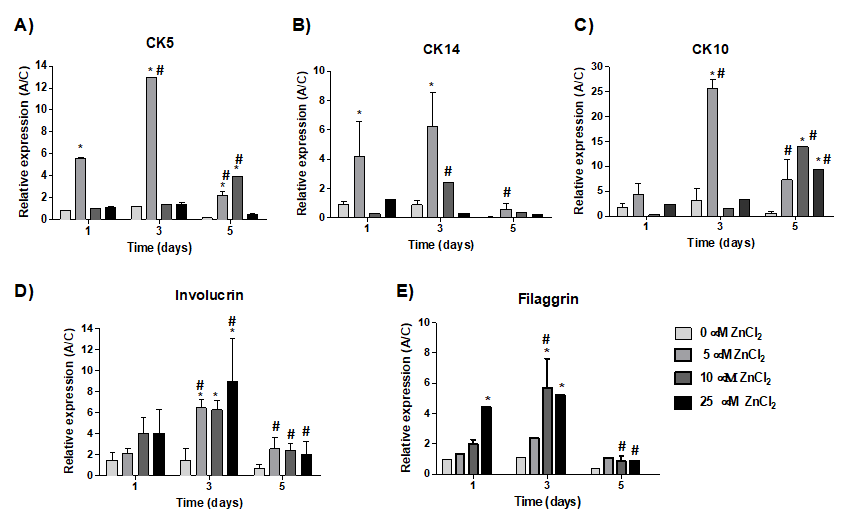
Figure 6: Relative expression of epidermal markers by qPCR. Cells were cultured in the absence or presence of different concentrations of Zn2+ ions (5.0; 10 and 25 μM) in the form of ZnCl2. Expression was normalized by the constitutive gene GAPDH. *Different in comparison to the control of the same period (ρ<0.05). Different in comparison to the same concentration in different periods.
Cell cultivation for 3 days in the presence of all Zn2+ ions concentration resulted in an increase in the involucrin protein expression, up to 6 times, in comparison to control. Furthermore, the expression of this protein, on day 3, was in general different in comparison to the other periods for the same concentration of Zn2+ ions (Figure 6D). Filaggrin expression was also 5 times higher on day 3 in cells cultured in the presence of 10 and 25 µM Zn2+ compared to control (Figure 6E). However, the expression of filaggrin was similar between cultures cultivated for one or three days in the presence of 25 µM Zn2+.
Western Blotting
The presence of the involucrin protein in cell extracts was evaluated by Western blotting after cell cultivation with Zn2+ ions (5.0; 10 and 25 μM) for 1, 3 and 5 days (Figure 7A). Densitometric analysis showed a significant increase in the expression of this protein in cells cultivated in the presence of 25 μM Zn2+ ions on day 5 of cultivation, which was 3 times higher in relation to control at the same day, and 2.4 and 1.6 times higher when compared to the previous Zn2+ ions concentrations in the same period (Figure 8B).
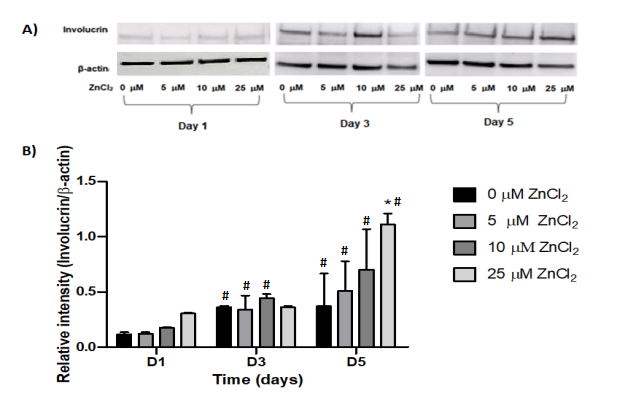
Figure 7 (A) & (B): Immunodetection of the involucrin protein. Cells cultured in the presence of different concentrations of Zn2+ ions (5.0; 10 and 25 μM) for 1, 3 and 5 days. Membranes were incubated with anti-involucrin (0.75 µg/ml) or anti-β− actin (1:1000) antibodies. Then, HRP-conjugated anti-mouse IgG secondary antibody (1:2000) was added. Visualization of the bands was performed using ECL. β-actin was used as constitutive protein. Densitometric analysis of involucrin expression. The densitometry results obtained in the CP Atlas 2.0 program were analyzed by ANOVA + Bonferroni post-test. *, different compared to control of the same cultivation period (ρ<0.05). #, different compared to the same treatment in the first day of cultivation (ρ<0.05)
Discussion
About 60% of the world’s population is affected by some dermatological problem, and 25% require medical care [20, 21]. Among the diseases that can affect the skin, psoriasis, in addition to having a significant impact on the quality of life of patients, require expensive treatments. Particularly, the use of topical therapies containing zinc is widespread due to its participation both in skin physiology and regeneration [14] and in various inflammatory skin conditions [22, 23].
It is already known that the increase in KLK5 and KLK7 activity leads to excessive degradation of corneodesmosomes, resulting in severe impairment of the epidermal barrier, accompanied by hyperplasia and abnormal differentiation of the epidermis [24]. Considering that Zn2+ ions exerts a strong inhibitory effect on the hydrolytic activity of KLK7 with an inhibition constant of 10 μM. In this work, considering that KLK5 and KLK7 participate in different events during the proliferation and differentiation of keratinocytes, and that zinc is an allosteric inhibitor of these enzymes, we studied the effect of this ion in a simplified in vitro system constituted by keratinocytes of the HaCaT lineage.
For this purpose, we first evaluated the effect of Zn2+ ions, in concentrations in the micromolar range, on the viability of these cells. The results confirmed the non-toxicity of Zn2+ ions in HaCaT epidermal cells when incubated for 48 h with this ion, and corroborate previous studies that showed that exposure of HaCaT keratinocytes to different concentrations of Zn2+ was also not toxic to skin cells [23, 25]. However, from the concentration of Zn2+ ions of 5 μM in 7 days of incubation, it was already possible to observe a decrease in cell viability evaluated by the MTT method, which could be related to a decrease in cell proliferation since cell death was excluded by flow cytometry experiments. Similarly, a study carried out by Kontargiris, et al. (2012) [26] demonstrated that 15
μM Zn2+ was able to inhibit the proliferation of HaCaT cells treated with ropivacaine, a local anesthetic. A decrease in viability was also observed when HaCaT cells were treated with 211 μM NpAgZnO [27].
In addition to the analysis of cytotoxicity and viability, it was evaluated the activity of the serine proteases KLK5 and KLK7 in cells cultured in the absence and presence of Zn2+ ions, using specific FRET substrates. Regarding KLK7, the results showed that there was a significant decrease in their activity on the hydrolysis of synthetic substrates in cells incubated for 1 day with 25 μM Zn2+ ions, compared to control. However, 5 days after culturing the cells in the presence of 25 μM Zn2+, despite the exchange of medium containing Zn2+ ions being performed every 2 days, the activity returned to baseline values, which could be partially explained by the fact that divalent ions, such as Zn2+, are reversible inhibitors of KLK [28, 29, 30]. Furthermore, it could be suggested that changes in the pH of the medium promoted the dissociation of LEKTI, a natural KLK inhibitor, resulting in the reestablishment of the activity of this enzyme [31, 32].
In addition to evaluating the effect of Zn2+ ions on KLK activity, particularly KLK7, we were interested in studying the effect of these ions on the expression of proteins involved in epidermal differentiation such as CK5, CK14, CK10, involucrin and filaggrin during the differentiation process [33].
The increase in CK5 and CK14 expression on days 1 and 3 of culturing, combined with its decrease on day 5 of culture, suggests that these cells possesses characteristics of basal keratinocytes at the beginning of the culture period; however, on day 5, there is an enrichment of more differentiated cells. This decrease also allowed other proteins involved in the terminal differentiation of keratinocytes to be expressed, such as involucrin and filaggrin [11].
It is already known that CK5 and CK14 form the primary keratin pair of epidermal keratinocytes. They are expressed in basal keratinocytes and undergo downregulation when these cells terminally differentiate, therefore being used as an initial marker of epidermal differentiation [34]. The loss of regulation in its expression is associated with the development of various dermatitis [35]. The results showed that in cells cultivated in the presence of 5 μM Zn2+ ions there was a 7 to 10-fold increase in the expression of the RNA that codes for these proteins on day 3, when compared to the other periods and concentrations.
The expression of CK10, a protein found in the first suprabasal layers of the stratum spinosum [36], was also analyzed by qPCR. The expression of both CK10, and CK1 indicate that the cells were committed to the terminal differentiation of keratinocytes [18]. A significant increase in CK10 expression was verified on days 3 and 5 of cultivation, in the presence of 5 and 10 μM Zn2+ ions, respectively, in relation to control in the same period. Considering that the expression of CK1 and CK10 occurs in the spinous layer of epidermis, it could be suggested that Zn2+ ions may act, through some mechanism that was not explored in this work, in the cell differentiation mechanism.
Regarding to involucrin, it is known that in humans it is an important precursor for the envelope formation and cell cohesion [37]. Decreased expression of this protein leads to structural abnormalities of the skin and hair, while its high expression in the epidermis is related to adequate skin barrier function [38]. Considering that in psoriatic lesions there is low expression of involucrin [39, 40], it could be suggested that the increased expression of this protein promoted by the incubation of cells with Zn2+ ions, could open perspectives for its use in therapeutic attempts for this disease.
The results by Western blotting showed an increase in the expression of the involucrin protein on day 5 of culturing in the presence of Zn2+ ions in all tested concentrations in comparison to cells cultured in the absence of this ion for the same period. Interestingly, the highest increase in the expression of the mRNA that encodes for involucrin, evaluated by qPCR, was verified on day 3 of cultivation, suggesting the accumulation of this protein in the cytoplasm of the cells from day 3 forward.
Another important protein in the process of epidermal differentiation is the filaggrin, responsible for skin keratinization [41]. The analysis of filaggrin expression by qPCR in cells cultured in the presence of 25 μM Zn2+ ions, for 1 or 5 days, showed an increase in its expression when compared to the control without Zn2+ ions in these same periods, and on day 3 when the cells were incubated with 10 μM Zn2+. Such increase could be related to the fact that zinc is involved in the regulation of filaggrin expression through the prostatin pathway, a zinc-dependent protease that processes profilaggrin to its respective monomer [42]. Likewise, the increase in filaggrin expression promoted by Zn2+ ions could suggest the use of this ion in the potential treatment of psoriasis, since this epidermal differentiation marker also is reduced expression in psoriatic lesions [40]. Furthermore, it is known that the loss or reduction in filaggrin expression combined with the abnormal differentiation of keratinocytes contribute to a permissive environment that facilitate the entry of irritants, allergens and microorganisms, which may evoke immune responses related to progressive weakening of the epidermis barrier, a relevant phenomenon in chronic skin diseases [43]. KLK7 also contributes to the recruitment of innate immune cells by activating the pro-inflammatory cytokine IL-1β [24]. This cytokine participates in the damage to the plasma membrane and in the molecular mechanism involved in the epidermal hyperproliferation observed in psoriasis [Cai et al., 2019]. In this sense, it was shown that IL-1β expression and presence were significantly elevated in injured psoriatic skin. Therefore, although not investigated in this study, it may be suggested that KLK7 inhibition could decrease IL-1β processing [44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55] consequently attenuating psoriasis symptoms.
In summary, it was found that the tested concentrations of Zn2+ ions, in the micromolar range, were not cytotoxic for HaCaT cells. However, in the presence of these ions, there was a small decrease in the number of viable cells, evaluated by the MTT method, which could suggest a decrease in cell proliferation, since there was no cell death under the same tested conditions. Specifically, this result is interesting considering the treatment of psoriasis, a disease in which rapid proliferation and accelerated transit of keratinocytes is observed. Regarding KLK activity, there was a decrease in KLK7 activity in cultures incubated for 1 day with 25 μM Zn2+ ions. In addition, there was an increase in the expression of proteins involucrin, filaggrin, CK5, CK10 and CK14, evaluated by qPCR, on different days of culturing, suggesting that Zn2+ ions may act in the process of epidermal differentiation. Together these data show the potential use of Zn2+ ions in a tentative treatment for psoriasis, an inflammatory skin disease characterized by epidermal hyperplasia and excessive proliferation of keratinocytes.
Funding: This work was supported by Fundação de Amparo à Pesquisa do Estado de São Paulo – FAPESP (2019/11693- 0).
Competing Interests: The authors have no financial or non- financial interests to disclose.
Author Contributions: All authors contributed to the study conception and design. Material preparation, data collection and analysis were performed by Willian Moreira Miguel, Bruna Letícia Freitas-Marchi and Viviane Abreu Nunes. The first draft of the manuscript was written by Viviane Abreu Nunes and Bruna Letícia Freitas-Marchi and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Data Availability Statement: Data available on request.
Ethical approval: This article does not contain any studies with human participants or animals performed by any of the authors.
References
-
Rendon A, Schakel K (2019) Psoriasis Pathogenesis and Treatment. Inter J Mol Sci 20(6): 1475.
-
Yang Y, Yangmeng Z, Rui L, Li X, Qirong L, et al. (2022) An Emerging Role of Proanthocyanidins on Psoriasis: Evidence from a Psoriasis-Like Mouse Model. Oxid Med Cell Longev, pp: 1-13.
-
Knox S, Boyle NM (2021) Skin Lipids in Health and Disease: A Review. Chem Phys Lipids 236: 105055.
-
Weiglein A, Gaffal E, Albrecht A (2022) Probing the Skin- Brain Axis: New Vistas Using Mouse Models. Inter J Mol Sci 23(13): 7484.
-
Gendrisch F, Philipp RE, Christoph MS, Ute W (2021) Luteolin as a Modulator of Skin Aging and Inflammation. Biofactors 47(2): 170-180.
-
Tang ZL, Kai ZB, Shao CL, Gui WX, Jian FZ, et al. (2021) Lncrna Meg3 Suppresses Pi3k/Akt/Mtor Signalling Pathway To Enhance Autophagy And Inhibit Inflammation In Tnf-Α-Treated Keratinocytes And Psoriatic Mice. Cytokine 148: 155657.
-
Shou Y, Yang L, Yongsheng Y, Jinhua X (2021) Inhibition of Keratinocyte Ferroptosis Suppresses Psoriatic Inflammation. Cell Death & Disease 12(11): 1009.
-
Santos JF, Bruna L, Gustavo RR, Silvia R, Viviane A et al. (2023) Mesenchymal Stem Cells Express Epidermal Markers in an in Vitro Reconstructed Human Skin Model. Front Cell Dev Biol 10: 1012637.
-
Khafaji Z, Brito S, Bin BH (2022) Zinc and Zinc Transporters in Dermatology. Int J Mol Sci 23(24): 16165.
-
Morizane S, Sunagawa K, Hayato N, Mamoru O (2022) Aberrant Serine Protease Activities in Atopic Dermatitis. J Dermatol Sci 107(1): 2-7.
-
Kim J, Min GK, Sang HJ, Hee JK, Sang WS (2022) Stat3 Maintains Skin Barrier Integrity by Modulating Spink5 and Klk5 Expression in Keratinocytes. Exp Dermatol 31(2): 223-232.
-
Geng X, Marion K, Holger B, Tobias D, Viktor M, et al. (2020) High Levels of Klk7 Protein Expression are Related to A Favorable Prognosis in Triple-Negative Breast Cancer Patients. Am J Cancer Res 10(6): 1785- 1792.
-
Yeo H, Sung SA, Jeong YL, Soon YS (2021) Egr-1 Acts as a Transcriptional Activator of Klk7 Under Il-13 Stimulation. Biochem Biophys Res Commun 534: 303- 309.
-
Ogawa Y, Manao K, Shinji S, Tatsuyoshi K (2018) Zinc in Keratinocytes and Langerhans Cells: Relevance to the Epidermal Homeostasis. J Immunol Res 2018: 1-11.
-
Boukamp P, Petrussevska RT, Breitkreutz D, Hornung J, Markham A, et al. (1988) Normal Keratinization in a Spontaneously Immortalized Aneuploid Human Keratinocyte Cell Line. J Cell Biol 106(3): 761-771.
-
Riccardi C, Nicoletti I (2006) Analysis of Apoptosis by Propidium Iodide Staining and Flow Cytometry. Nature Protocols 1(3): 1458-1461.
-
Kruger NJ (2002) The Bradford Method for Protein Quantitation. In: The Protein Protocols Handbook, UK, pp: 17-24.
-
Fernandes MTP, Jeniffer F, Bruna L, Gustavo R, Fernanda A, et al. (2022) Reporter System Controlled by the Involucrin Promoter as a Tool to Follow Epidermal Differentiation. Biochimie 201: 33-42.
-
Kavlick MF (2019) Development of a Triplex Mtdna Qpcr Assay to Assess Quantification, Degradation, Inhibition and Amplification Target Copy Numbers. Mitochondrion 46: 41-50.
-
Eissa A, Diamandis EP (2013) Human Tissue Kallikreins as Promiscuous Modulators of Homeostatic Skin Barrier Functions. Biol. Chem 389(6): 669-680.
-
Cheong JEL, Mcgrath JA (20009) Structure and Function of Skin, Hair and Nails. Medicine 37(5): 223-226.
-
Kawamura T, Ogawa Y, Nakamura Y, Nakamizo S, Ohta Y, et al. (2012) Severe Dermatitis with Loss of Epidermal Langerhans Cells in Human and Mouse Zinc Deficiency. J Clin Invest 122(2): 722-732.
-
Emri E, Miko E, Bai P, Boros G, Nagy G, et al. (2015) Effects of Non-Toxic Zinc Exposure on Human Epidermal Keratinocytes. Metallomics 7(3): 499-507.
-
Kishibe M, Baida G, Bhalla P, Lavker RM, Schlosser B, et al. (2016) Important Role of Kallikrein 6 for the Development of Keratinocyte Proliferative Resistance to Topical Glucocorticoids. Oncotarget 7(43): 69479- 69488.
-
Salesa B, Serra RSI, Aroca AS (2021) Zinc Chloride: Time- Dependent Cytotoxicity, Proliferation and Promotion of Glycoprotein Synthesis and Antioxidant Gene Expression in Human Keratinocytes. Biology 10(11): 1072.
-
Kontargiris E, Vadalouka A, Ragos V, Kalfakakou V (2012) Zinc Inhibits Apoptosis and Maintains Nep Downregulation, Induced by Ropivacaine, In Hacat Cells. Biol Trace Elem Res 150(1-3): 460-466.
-
Rajendran R, Mani A (2020) Photocatalytic, Antibacterial and Anticancer Activity of Silver-Doped Zinc Oxide Nanoparticles. Journal of Saudi Chemical Society 24(12): 1010-1024.
-
Goettig P, Magdolen V, Brandstetter H (2010) Natural and Synthetic Inhibitors of Kallikrein-Related Peptidases (Klks). Biochimie 92(11): 1546-1567.
-
Brattsand M, Stefansson K, Lundh C, Haasum Y, Egelrud T (2005) A Proteolytic Cascade of Kallikreins in the Stratum Corneum. J Invest Dermatol 124(1): 198-203.
-
Quan CJ, Huang LB, Ping LH, Yin MZ, Lan ZH (2019) Roles of Kallikrein-Related Peptidase in Epidermal Barrier Function and Related Skin Diseases. Inter J Dermatol Venereol 2(3): 150-155.
-
Teixeira, Thiago SP (2011) Molecular modeling and enzymatic inhibition study of human tissue kallikreins 5 and 7 by isocoumarin compounds and isomanide derivatives. Dissertation (Master’s in Biochemistry, Physiology and Pharmacology) - Federal University of Triangulo Mineiro.
-
Paolo CTD, Diamandis EP, Prassas I (2021) The Role of Kallikreins in Inflammatory Skin Disorders and their Potential as Therapeutic Targets. Crit Rev Clin Lab Sci 58(1): 1-16.
-
Furue M (2020) Regulation of Filaggrin, Loricrin, and Involucrin by Il-4, Il-13, Il-17a, Il-22, Ahr, and Nrf2: Pathogenic Implications in Atopic Dermatitis. Int J Mol Sci 21(15): 5382.
-
Bayati V, Abbaspour MR, Dehbashi FN, Neisi N, Hashemitabar M (2017) A Dermal Equivalent Developed from Adipose-Derived Stem Cells and Electrospun Polycaprolactone Matrix: An in Vitro And in Vivo Study. Anat Sci Int 92(4): 509-520.
-
Wang X, Li LF, Zhao DY, Shen YW (2016) Prevalence and Clinical Features of Atopic Dermatitis in China. Biomed Res Int 2016: 2568301.
-
Kumar A, Jagannathan N (2018) Cytokeratin: A Review on Current Concepts. Inter J Orofacial Biol 2(1): 6.
-
Belanger A, Grenier A, Simard F, Gendreau I, Pichette A, et al. (2020) Dihydrochalcone Derivatives from Populus balsamifera L. Buds for the Treatment of Psoriasis. Inter J Mol Sci 21(1): 256.
-
Varma SR, Sivaprakasam TO, Arumugam I, Dilip N, Raghuraman M, et al. (2019) In vitro anti-inflammatory and skin protective properties of virgin coconut oil. J Tradit Complement Med 9(1): 5-14.
-
Honda T, Kabashima K (2019) Prostanoids and leukotrienes in the pathophysiology of atopic dermatitis and psoriasis. International Immunology 31(9): 589- 595.
-
Orsmond A, Bereza ML, Lynch T, March L, Xue M (2021) Skin barrier dysregulation in psoriasis. Int J Mol Sci 22(19): 10841.
-
Jang S, Jang S, Kim SY, Ko J, Kim E, et al. (2021) Overexpression of Lin28a aggravates psoriasis- like phenotype by regulating the proliferation and differentiation of keratinocytes. J Inflamm Res 14: 4299- 4312.
-
Suzuki M, Suzuki T, Watanabe M, Hatakeyama S, Kimura S, et al. (2021) Role of intracellular zinc in molecular and cellular function in allergic inflammatory diseases. Allergol Int 70(2): 190-200.
-
Jiang Y, Tsoi LC, Billi AC, Ward NL, Harms PW, et al. (2020) Cytokinocytes: the diverse contribution of keratinocytes to immune responses in skin. JCI Insight 5(20): e142067.
-
Cai Y, Xue F, Quan C, Qu M, Liu N, et al. (2019) A Critical Role of The Il-1β–Il-1r signaling pathway in skin inflammation and psoriasis pathogenesis. J Invest Dermatol 139(1): 146-156.
-
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72(1-2): 248-254.
-
Debela M, Hess P, Magdolen V, Schechter NM, Steiner T, et al. (2007) Chymotryptic specificity determinants in the 1.0 A structure of the zinc-inhibited human tissue kallikrein 7. Proc Natl Acad Sci 104(41): 16086-16091.
-
Gong Z, Dai S, Jiang X, Lee M, Zhu X, et al. (2023) Variants in KLK11, affecting signal peptide cleavage of kallikrein- related peptidase 11, cause an autosomal-dominant cornification disorder. Br J Dermatol 188(1): 100-111.
-
Kitajima Y (2015) Implications of normal and disordered remodeling dynamics of corneodesmosomes in stratum corneum. Dermatologica Sinica 33(2): 58-63.
-
Li Z, Jiang R, Jing C, Liu J, Xu X, et al. (2022) Protective effect of oligosaccharides isolated from panax ginseng C. A. Meyer against UVB-induced skin barrier damage in BALB/c hairless mice and human keratinocytes. J Ethnopharmacol 283: 114677.
-
Hoffert MU, Wu Z, Schroder JM (2009) Identification of lympho-epithelial kazal-type inhibitor 2 in human skin as a kallikrein-related peptidase 5-specific protease inhibitor. Plos One 4(2): e4372.
-
Nauroy P, Nystrom A (2020) Kallikreins: Essential Epidermal Messengers for Regulation of the Skin Microenvironment during Homeostasis, Repair and Disease. Matrix Biol Plus 6-7: P. 100019.
-
Nickerson DW, Angela PP, Stephen SW, Aric LG, John AB, et al. (2009) Quantification of Cytokeratin 5 mRNA Expression in the Circulation of Healthy Human Subjects and after Lung Transplantation. Plos One 4(6): E5925.
-
Park NJ, Sullim L, Yong KK, Min HY, Nam KS et al. (2022) Lobelia Chinensis Extract and its Active Compound, Diosmetin, Improve Atopic Dermatitis by Reinforcing Skin Barrier Function Through SPINK5/LEKTI Regulation. Inter J Mol Sci 23(15): 8687.
-
Santos JF, Nathalia RB, Mariana SA, Viviane AN (2019) Mesenchymal Stem Cells Differentiate into Keratinocytes and Express Epidermal Kallikreins: towards an in Vitro Model of Human Epidermis. J Cell Biochem 120(8): 13141-13155.
-
Wang XW, Salimi M, Gwela A, Wang YJ, Giannoulatou E, et al. (2017) Deficiency of Filaggrin Regulates Endogenous Cysteine Protease Activity, Leading to Impaired Skin Barrier Function. Clin Exp Dermatol 42(6): 622-631.
- Superposition of Cryo-EM and AlphaFold Predictions of Dengue Antigen-Antibody Complexes
- Jugular-Applied Coherent Low-Level Laser Therapy Enhances Systemic Mitochondrial Metabolic Function and Antioxidant Response
- Role of OMC32 Polypeptide in Acrosin-Mediated Exocytosis during the Bovine Sperm Acrosome Reaction
- Association of Galectin-3 but not Laminin in Tamoxifen-Induced Growth Suppression in Breast Cancer MCF-7 Cells
- Effect of Different Wavelengths of Light on the Rate of Photosynthesis
- Nutritional, Therapeutic, and Environmental Effect of Oyster Mushrooms: An Editorial