Analysis of Neotropical Serum Fishes Esterases Activities for the Selection of Noninvasive Enzymatic Biomarkers
Enzymatic biomarkers can be effective tools for monitoring the health of aquatic organisms exposed to xenobiotics. The use of sample blood is beneficial to preservation, since it allows evaluations without the euthanasia of the animal, preserving the fauna and the balance of the ecosystem. Nevertheless, it is previously necessary to understand the kinetics of the enzymes and to determine their basal activity levels for the proper choice of the enzyme to be used for biomarking. The best assay conditions of carboxylesterase activity (CaE) present in the serum of four different species of neotropical fish were established and their activities were compared with other blood esterases. Also, the CaE were measured at summer and winter to verify possible seasonal changes. The CaE was assayed with the substrate para-nitrophenylacetate (p-NPA) by the appearance of product in one minute reaction. The dourado CaE had a KM and Vmax value similar to that of pacu, but the curimbatá showed the best catalytic efficiency to degrade p-NPA. In piauçu the activity of serum CaE is not as efficient when compared to the other species. Dourado, pacu and curimbatá have higher CaE than cholinesterase activity, distinct of piauçu. The CaE of pacu serum decreases in the winter. Thus, it is shown to be important to establish the level of activity of each esterase per species and to verify if seasonal variation may influence the analysis in the monitoring of enzymatic biomarkers.
Introduction
Assessments of pollutants effects on organisms aim to estimate how intensely anthropogenic intervention can occur without greatly altering the ecological balance of an area. Samples obtained from animals, as tissues, cells or biological fluids [1] are useful for assessing and controlling the risk associated with long-term exposure to xenobiotics, such as heavy metals, hydrocarbons and agrochemicals [2].
The biomarkers compare a normal biological process with the value obtained in a pathogenic or stress state. Since the enzymatic reactions can show variations, it is possible to obtain information about the health of the animals and the environment that they inhabit through tissue samples. The enzymatic biomarkers are interesting in monitoring because they may reflect sublethal responses, indicating early damage before tissue lesions are established. Procedures to found early warning signals that reflect adverse biological responses due to anthropogenic changes stand out in researches for enabling actions prior to extreme environmental damage [3].
Esterases are inhibited by phosphates, therefore cholinesterases has been widely used to evaluate the harmful effects of pesticides and are widespread as fish biomarkers of exposure because they affinity to organophosphates and carbamates compounds [4, 5, 6]. Liver, muscle and brain samples are frequently used for enzymatic analysis of cholinesterase. Unfortunately, holding these samples often implies the death of the specimen, thus the use of blood sample seems to be a better procedure.
The kinetics of blood enzymes of neotropical fish species it is not well known. Some studies have already shown that fishes have biochemical mechanisms, involving the inhibition of esterases, which protect them against exposure to organophosphates. Salles, et al. [7] demonstrated that the Pantanal piauçu plasma activity of butyrylcholinesterase (BChE) is three times higher than human plasma and 34 times higher than tilapia plasma, and that this is the main esterase “capture” of organophosphates present in the plasma of piauçu. Cunha Bastos, et al. [8] verified that paraoxonase activity in dourado serum reached values six times higher than those found in pacu and cascudo serum, and 23 times higher than in serum of matrinxã.
Organophosphates can also bind irreversibly to another non-target esterases, for example the carboxylesterase (CaE). Such binding makes the CaE to act as a detoxification enzyme, because when it binds organophosphates it prevents an increase in the concentration of organophosphate at the site of the target enzyme, being able to alter the toxicity profile of some pesticides that inhibit cholinesterases [9]. According to Satoh T [10] wide variety of carboxylesterases found in tissues and plasma show greater promiscuity in substrates for which they catalyze, so the multiplicity of structure of these enzymes allows hydrolyzing a diverse range of esters. Hence, this esterase may be involved in protecting the organism against xenobiotics and being an interesting biomarker.
The characterizations of carboxylesterases are more related to insects and humans, and there are few studies with non-target organisms, such as fishes [11]. Dalzochio, et al. [12] studying biomarkers of aquatic environments in Brazil showed that the researches are concentrated in only two states, lacking reports on a Center-West and Northern region. Therefore, researches with neotropical fish are important to enrich biochemical knowledge and to understand the various adaptations found in these organisms. The aim of this study is to standardize and determine basal activity levels of esterases, focusing on carboxylesterase, in serum of four neotropical fish species at different seasons of the year as a subsidy for future studies with enzymatic activity biomarker in fish serum.
Material and Methods
Fishes and Obtaining Serum
Four specie of neotropical fishes were analyzed: the dourado Salminus brasiliensis, the pacu Piaractus mesopotamicus, the curimbatá Prochilodus lineatus and the piauçu Leporinus macrocephalus.
The blood samples were obtained from adults’ fishes coming from farming. The biometric data are present in Table 1.
| Species | Dourado | Pacu | Curimbatá | Piauçu |
|---|---|---|---|---|
| n | 42 | 22 | 5 | 3 |
| Weight (g) | 1704 ± 611 | 233 ± 129 | 290 ± 119 | 414 ± 156 |
| Total length (cm) | 49.4 ± 5.9 | 22.58 ± 3.66 | 30.6± 1.4 | 26.5 ± 3.5 |
Table 1: Biometric data of species.
Blood was collected by puncture of a vessel from the caudal peduncle. The blood was let in an assay tube on ice to clot. Clotted blood was centrifuged at 3500 rpm for 10 minutes. The straw-yellowish supernatant serum was kept frozen until analysis. Serum Esterases Enzyme Assays Carboxylesterase activity (CaE) was carried out with para-nitrophenyl acetate (p-NPA) as substrate diluted in acetonitrile 100% according to Morgan, et al. [13]. The reaction medium was buffered with 0.1 M sodium phosphate solution, pH 7.7 at 25°C. The enzyme activity was measured by determining the appearance of para-nitrophenol for one minute at the wavelength of 400 ηm on a Shimadzu UV-1800 spectrophotometer. The molar absorptivity coefficient of p-nitrophenol used was 13,000 M-1.cm-1. For which specie were made a substrate standard curve to verify optimal assays conditions. The p-NPA substrate has spontaneous hydrolysis. To record only the catalyzed reaction, a blank was made consisting of the activity test without adding the sample. The enzymatic activity was obtained by subtracting the blank value from the value of each assay.
Cholinesterase activity (ChO) was conducted according to Ellman, et al. [14], using a Shimadzu UV-1800 spectrophotometer measuring continuously recorded up to one minute at 412 ηm. The reaction was carried out in buffer solution of sodium phosphate 0.1 M, pH 7.5, at 25°C, containing 1.87 mM ASCho or BSCho as substrates, in the presence of 0.32 mM DTNB. The molar absorptivity coefficient used for ChO was 14,150 M-1.cm-1.
Determination of KM and Vmax and Statistical Analyses
The values of maximum velocity (Vmax) and apparent KM were determined using the Michaelis–Menten equation with the GraphPad Prism software. All enzymatic activities were expressed as µmol.min-1.mL-1 of serum and the results are presented as mean ± standard deviation. A value of p <
0.05 was considered statistically significant.
Results
Standardization of Serum CaE Assay
Blood from at least three specimens was collected to obtain a serum pool to establish how the variation of p-NPA concentration affects the initial velocity reaction. The Michaelis-Menten curve was obtained for each species, as shown in the graphs of Figure 1. The results of the standardization also determined the Vmax for p-NPA, as well the apparent KM value (Table 2).
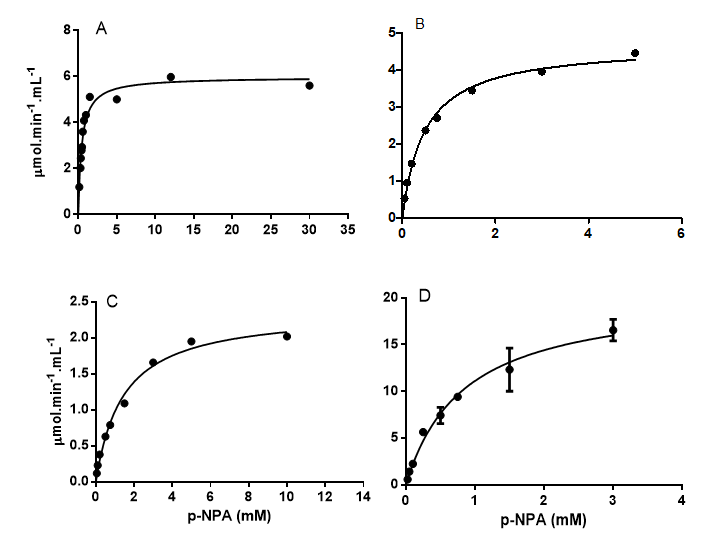
After determining the optimum assay conditions with the p-NPA substrate for each species, the CaE in the serum of each individual was assayed and the mean basal level is demonstrated in Table 3. Cholinesterases activities were also verified in the serum of the four species (Table 3).
Comparison between Species
The values of KM and Vmax varied between species (Table 2). The curimbatá presented the highest Vmax and the best catalytic efficiency, however, the KM value is the second largest. The dourado and pacu had close values. The piauçu CaE serum activity is not as efficient when compared to the other fishes.
| Species | CaE in serum | ||
|---|---|---|---|
| KM (mM) | Vmáx (µmol.min-1.mL-1) | Vmáx/KM k (min-1) | |
| Dourado (Salminus brasiliensis) | 0,41 | 5,73 | 13,97 |
| Pacu (Piaractus mesopotamicus) | 0,48 | 4,69 | 9,77 |
| Curimbatá (Prochilodus lineatus) | 0,84 | 20,45 | 24,34 |
| Piauçu (Leporinus macrocephalus) | 1,43 | 2,38 | 1,66 |
Table 2: Values of Vmax, apparent KM and catalytic efficiency of p-NPA hydrolysis by neotropical fish’s serum pool.
Comparing the value of CaE with ChO activities in fish serum, it is possible to verify that for each species there is a variation between the average activities measured for each enzyme (Table 3). The piauçu exhibits superior activity of BChE in relation to other esterases, whereas in dourado the BChE is almost undetectable. The dourado, pacu and curimbatá have in serum higher activity of CaE than ChO.
| Serum | Enzymatic activity (µmol.min-1.mL-1) | ||
|---|---|---|---|
| AChE | BChE | CaE | |
| Dourado | 0,14 ± 0,04 | 0,01 ± 0,06 | 6,96 ± 3,71 |
| Pacu | 0,74 ± 0,18 | 1,03 ± 0,30 | 3,67 ± 2,00 |
| Curimbatá | 3,02 ± 1,69 | 5,85 ± 3,46 | 16,95 ± 1,08 |
| Piauçu | 9,35 ± 4,92* | 21,72 ± 11,74* | 1,56 ± 0,29 |
Table 3: Esterases basal enzymatic activities in serum of neotropical fishes.
*Reference Salles. et al. [7]. Table 3: Esterases basal enzymatic activities in serum of neotropical fishes.
Seasonal Comparison
For the dourado and pacu it was possible to collected blood during the winter and summer, in order to verify if
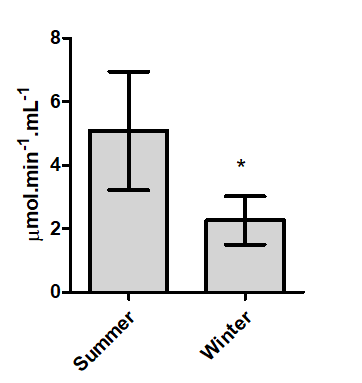
there is a seasonal change. The pacu serum CaE have seasonal variation (Figure 2), with higher activity in summer (5.08 ± 2.26 µmol.min-1.mL-1) than in winter (1.86 ± 0.75 μmol .min-1. mL-1).
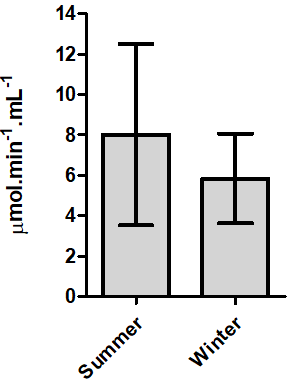
Although the dourado CaE serum also tended to decrease during the winter, no statistically significant difference was observed for the average activity (Figure 3).
Discussion
The hydrolysis of p-NPA was detected in the serum of the four neotropical fish species evaluated. The assay of CaE activity has desirable characteristics for a biomarker, since it is fast to analyze, have low cost and can be measured in serum. Wheelock, et al. [17] reported that the general CaE activity of colorimetric assays using substrates such as p-NPA or α-naphthyl acetate are convenient because of their easy availability and relatively inexpensive optical assay required for data acquisition. However, the authors point out that some CaE isoenzymes do not hydrolyses these substrates efficiently.
The use of serum has proved to be practical for assaying enzymatic biomarkers, being CaE and BChE the better detection activities. An expressive advantage of these biomarkers is that the assays preserves the animal life, in contrast to the use of AChE activity in muscle or brain. The analysis of CaE from the blood of the fish presents as a viable and a less invasive method. According to Thompson, et al. [18] plasma carboxylesterase activity is generally low in mammals, but is higher in fish, amphibians and birds.
Among the studied species, referent to CaE, curimbatá has the best catalytic efficiency, nevertheless the Vmax is reached in contact with higher p-NPA concentrations. The dourado and pacu have the lowest KM, therefore reaching their maximum enzymatic efficiency with smaller amounts of substrate in the reaction medium. In rats, a study by Maxwell, et al. [15] involving CaE and reactions with organophosphorus compounds has shown that CaE appears to perform the role of a high-affinity/low-capacity detoxication process and this high CaE affinity is important for detoxication of very toxic organophosphates compounds that achieve lower in vivo concentrations. Thus, having a prompt and efficient catalytic activity of CaE in serum indicates that the species can capture stoichiometrically or hydrolyze more xenobiotics, avoiding to inhibit essential cholinesterases for the organism if they reach vital organs such as the heart and the brain.
Wheelock, et al. [19] testing the interaction of esterases and organophosphates in salmon tissues, detailed that understanding the change in the level of CaE, this activity can be used as biomarkers of fish exposure to agrochemicals. Among the species of neotropical fish evaluated, the curimbatá is probably the animal that is more resistant to contamination by organophosphates due to the higher CaE serum activity. However, piauçu has elevate serum BChE, which also gives it the ability to hydrolysis organophosphates efficiently.
Salles, et al. [7] demonstrated that the activity of AChE and BChE in serum of several fish species varies considerably, a fact corroborated by the results obtained with the serum CaE in the present study. Solé, et al. [20] characterizing different tissues esterases of a flatfish (Solea senegalensis) also found that in adult plasma the largest esterase activity is CaE. Thus, it is important to establish the basal level activity of each blood esterase per species, to choose the enzymatic biomarker that will best complete the desired monitoring.
The activity of pacu and dourado serum CaE has tended to decrease in the winter, therefore it is possible that during the winter species of neotropical fish are more susceptible to xenobiotic effects than in the summer. Dalzochio, et al. [12] compiled information on the use of biomarkers to assess the health of aquatic ecosystems in Brazil. Among their findings, they point out that the evaluation of more than one sampling site at different seasons may support in the interpretation of biomarkers responses. Solé, et al. [20] analyzed samples of goby sand fish (Pomatoschistus minutus) for one year to select biomarkers. The authors noted fluctuations in enzymatic activity, some of which are considered to be normal seasonal variations due to breeding period, but other changes may be due to local pollution inputs.
The enzymatic activities analyzed in the serum were important to establish the basal level for four neotropical fish species and to demonstrate that seasonal differences may occur. The results obtained are useful for inferring future analyzes, as well as for assisting in the plan of aquatic environments monitoring.
References
-
WHO (2004) International Programme on Chemical Safety – IPCS: Risk assessment terminology.
-
Gil F, Pla A (2001) Biomarkers as biological indicators of xenobiotic exposure. J Appl Toxicol 21: 245-255.
-
Van Der Oost R, Beyer JB, Vermeulen NPE (2003) Fish bioaccumulation and biomarkers in environmental risk assessment: a review. Environ Toxicol Pharmacol 13: 57- 149.
-
Vincze K, Scheil V, Kuch B, Köhler HR, Triebskorn R (2015) Impact of wastewater on fish health: a case study at the Neckar River (Southern Germany) using biomarkers in caged brown trout as assessment tools. Environ Sci Pollut Res 22: 11822-11839.
-
Ghazala SM, Ahmad L, Sultana S, Alghanim K, Al-misned F, et al. (2014) Fish cholinesterases as biomarkers of sublethal effects of organophosphorus and carbamates in tissues of _Labeo rohita_. J Biochem Mol Toxicol 28: 137- 142.
-
Arias ARL, Buss DF, Alburquerque C, Inácio AF, Freire MM, et al. (2007) Utilização de bioindicadores na avaliação de impacto e no monitoramento da contaminação de rios e córregos por agrotóxicos. Ciênc. saúde coletiva 12: 61- 72.
-
Salles JB, Cunha Bastos VLF, Silva Filho MV, Machado OLT, Salles CMC, et al. (2006) A novel butyrylcholinesterase from serum of _Leporinus macrocephalus_, a Neotropical fish. Biochim 88: 59-68.
-
Cunha Bastos VLF, Alves MV, Bernardino G, Ceccarelli PS, Cunha Bastos J (2004) Paraoxonase activity in sera of four neotropical fish. Bull Environ Contam Toxicol 72: 798-805.
-
Chanda SM, Mortensen SR, Moser FVC, Padilla S (1997) Tissue-specific effects of chlorpyrifos on carboxylesterase and cholinesterase activity in adult rats: an in vitro and in vivo comparison. Fundam Appl Toxicol 38: 148-157.
-
Satoh T (2005) Toxicological implications of esterases- From molecular structures to functions. Toxicol Appl Pharmacol 207: S11-S18.
-
Wheelock CE, Nakagawa Y (2010) Carboxylesterases – from function to the field: an overview of carboxylesterase biochemistry, structure–activity relationship, and use in environmental field monitoring. J Pestic Sci 35: 215-217.
-
Dalzochio T, Rodrigues GZP, Petry IE, Gehlen G, Silva LB (2016) The use of biomarkers to assess the health of aquatic ecosystems in Brazil: a review. Int Aquat Res 8: 283-298.
-
Morgan EW, Yan B, Greenway D, Petersen DR, Parkinson A (1994) Purification and characterization of two rat liver microsomal carboxylesterases (hydrolase A and B). Arch Biochem Biophys 315: 495-512.
-
Ellman GL, Courtney KD, Andres V, Feather-Stone RM (1961) A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem Pharmacol 7: 88- 95.
-
Maxwell DM (1992) The specificity of carboxylesterase protection against the toxicity of organophosphorus compounds. Toxicol Appl Pharmacol 114: 306-312.
-
Solé M, Kopecka J, García De La Parra LM (2006) Seasonal variations of selected biomarkers in sand gobies _Pomatoschistus minutus_ from the Guadalquivir Estuary, Southwest Spain. Arch Environ Contam Toxicol 50: 249-255.
-
Wheelock CE, Shan G, Ottea J (2005a) Overview of Carboxylesterases and Their Role in the Metabolism of Insecticides. J Pestic Sci 30: 75-83.
-
Thompson HM, Mackness MI, Walker CH, Hardy AR (1991) Species differences in avian serum B esterases revealed by chromatofocusing and possible relationships of esterase activity to pesticide toxicity. Biochem Pharmacol 41: 1235-1240.
-
Wheelock CE, Eder KJ, Werner I, Huang H, Jones PD, et al. (2005b) Individual variability in esterase activity and CYP1A levels in Chinook salmon (_Oncorhynchus_ _tshawytscha_) exposed to esfenvalerate and chlorpyrifos. Aquat Toxicol 74: 172-192.
-
Solé M, Vega S, Varó I (2012) Characterization of type ‘‘B’’ esterases and hepatic CYP 450 isoenzimes in Senegalese sole for their further application in monitoring studies. Ecotoxicol Environ Saf 78: 72-79.
- Genetic Improvement of Nile Tilapia (Oreochromis niloticus): Advances in Selective Breeding and Genomic Approaches for Sustainable Aquaculture
- Microplastics, Contaminants, and Waste Hotspots: Divergences and Faults in Prioritizing Control Efforts
- Creating a Healthier, More Vibrant Open and Closed Aquatic Environment. A Submersible, Centrifugal Magnetically Affixed Current Changing Aquarium Pump
- An Attempt to Assess Alpha Diversity and Sample Size: Using the Ostracod Assemblages off Kumamoto Port, Japan
- Assessment of the Efficiency of Common Fishing Gears and Crafts Used at Mohananda River of Chapai Nawabganj, Bangladesh
- Fish Productivity and Biodiversity Status of Sundarban Mangrove in Bangladesh