Effect of Dietary Energy Level on Some Reproductive Characteristics of Clarias jaensis Female (Boulenger, 1909), an Endogenous Species from Cameroon
To determine the level of food energy necessary for a good sexual maturation of Clarias jaensis females in captivity, 180 post fingerlings with average weight of 96 ± 7g were submitted in triplicates to four experimental isoproteic (40 % crude protein) diet with 3000, 3100, 3200 and 3300 Kcal/kg of energy. The study took place at the Application and Research Farm of the University of Dschang for 4 months. At the end of the test, 12 females presenting the sexual maturity characteristics (bloated belly containing mature oocytes) from each group were sacrificed. The results showed that FSH (0.81 ± 0.35 mlU/ml) and LH (0.66 ± 0.23 mlU/ml) were higher at 3100 Kcal/kg while the higher concentration of estradiol (450,00 ± 109,86 pg/ml), the higher weight of ovaries (31,88 ± 6,86g) and the higher rate of mature female (75%) were recorded with 3000 Kcal/kg. Other reproductive characteristics such as ovarian biochemical characteristics, GSI, HSI and oocyte characteristics were also better with lower energy diets. In conclusion, females with the best reproductive characteristics were obtained with low energy rations.
Abbreviations
FSH: Follicle Stimulating Hormone; LH: Luteinizing Hormone; TGS: Triglycerids; LDL: Low Density Lipoprotein Cholesterol;
HDLC: High Density Lipoprotein Cholesterol; GSI: Gonado Somatic Index; HSI: Hepatho-Somatic Index.
Introduction
The disappearance of fishery resources is linked to the fact that most of the products of aquatic origin consumed by the population are taken from the natural environment. Indeed, 50% of the proteins of animal origin consumed come from fishing, which unfortunately has already reached its limits due to overexploitation of resources. Hence, the need to find an alternative such as aquaculture which has not stopped growing in recent years [1]. To be efficient and sustainable, aquaculture must be based on the breeding of endogenous species able of easily resisting environmental constraints and producing better.
On the other hand, the breeding of endogenous species contributes to preserving and restoring biodiversity through repopulation [2, 3]. Among the endogenous species likely to be farmed are some of the Clariidae family, also called catfish. They represent almost a third of the freshwater fish known in the world [4, 5]. Their interest in fish farming was summarized by Ducarme C, et al. [6] and is linked to their ability to easily adapt to climate change, their omnivorous diet, their rapid growth correlated with the ability to efficiently transform compound feed and to their highly valued flesh [7, 8]. In Cameroon, Clarias gariepinus and Heterobranchus longifilis, non-native species, are the most cultivated catfish [9, 10]. In the Mbô floodplain of the West region of Cameroon, there is an endogenous Clariidae named Clarias jaensis which has proven to be a good candidate for aquaculture. The first works to our knowledge on this species are those of Pouomogne V [11], Zango P [12], Mfossa MD [13], Zango P, et al. [14], Etaba AD, et al. [15] and Efole ET, et al. [16] who worked on various aspects of its reproduction and growth. However, for its production to be effective, it would be necessary to determine all the standards for its breeding. It would therefore be interesting to initiate investigations relating to nutrition and their effects on the growth and reproduction characteristics of Clarias jaensis.
Nutrition fundamentally affects the production performance of fish through its levels of proteins, minerals, vitamins and energy [17, 18]. According to Barnabe G [19], the growth and maturation of the gonads does not occur in lean fish that do not have sufficient mobilizable energy reserves. Some studies on Clarias jaensis feeding have revealed that the energy and protein level of the ration influences the survival and growth [20, 21]. The work of Tsoupou KSG, et al. [22] revealed that the level of dietary energy has a significant impact on the reproductive characteristics of Clarias jaensis males. However, according to the literature, no study has yet been carried out on the effect of feed energy on the reproductive characteristics of the female of Clarias jaensis.
The general objective of this work is therefore to contribute to the preservation and enhancement of fisheries biodiversity in general and Clariidae in particular by determining their nutritional needs. More specifically, the aim was to evaluate the effect of the optimum energy level of the ration on the reproductive characteristics of Clarias jaensis female.
Material and Method
Area of the Study
The study carried out in the west region of Cameroon and at the Application and Research Farm of the University of Dschang (LN 5°44’-5°36’ and 5°44’-5°37’; LE 10°06’- 9°94’ and 10°06’-9°85’ at an altitude of 1392 -1396 m.
Animal Material
180 post fingerlings female of Clarias jaensis with average weight and average total length of 96 ± 7g and 22.0 ± 1.7 cm were captured from the wild in Nkam river flood plain nearby Santchou settlemen. They were transported in oxygenated buckets and stocked in a breeding pond at the study site. The fish were acclimatized for two weeks and were fed ad libitum with a standard diet (3A) containing 40% of protein and consisting of wheat bran, soybean meal, and fishmeal [7].
Experimental Diet
Four isoproteic experimental diets Table 1 were formulated with different energy levels 3000, 3100, 3200 and 3300 Kcal/kg of food. The chosen energy levels refer to those of Clarias gariepinus and the principal source of energy was palm oil [20]. The chemical compositions of different diets were analyzed according to AOAC methods [23].
| Energy level (MJ/kg) | ||||
|---|---|---|---|---|
| Ingredients (g/Kg) | 3000 | 3100 | 3200 | 3300 |
| Maize | 28 | 24.5 | 22.5 | 20.3 |
| Wheat bran | 6 | 5.1 | 3.5 | 3.8 |
| Cottonseed cake | 1.5 | 1.3 | 1 | 1.5 |
| Soybean meal | 3 | 4.4 | 6.0 | 4.5 |
| Fish meal | 56.7 | 57.0 | 56.5 | 57.0 |
| Shell meal | 0.1 | 0.1 | 0.1 | 0.1 |
| Bone meal | 0.1 | 0.1 | 0.1 | 0.1 |
| Palm oil | 2.6 | 5.5 | 8.3 | 10.7 |
| Premix 2% | 2 | 2 | 2 | 2 |
| Total | 100 | 100 | 100 | 100 |
| Chemical Composition | ||||
| Crude Protein (%) | 39.76 ± 0.49 | 40.06 ± 0.75 | 40.66 ± 0.37 | 40.56 ± 0.50 |
| Crude Energy (MJ/kg) (kcal/kg) | 13.75 ± 0.10 | 14.04 ± 0.16 | 15.82 ± 0.09 | 16.11 ± 0,11 |
| Crude Lipid (%) | 7.00 ± 0.00 | 11.33 ± 0.28 | 12.00 ± 0.00 | 15.33 ± 0.57 |
| Ash (%) | 2,93 ± 0.05 | 1.96 ± 0.05 | 2.70 ± 0.17 | 2.06 ± 0.05 |
Table 1: Composition of experimental diets. (Source: Adapted from [21]).
Experimental Design
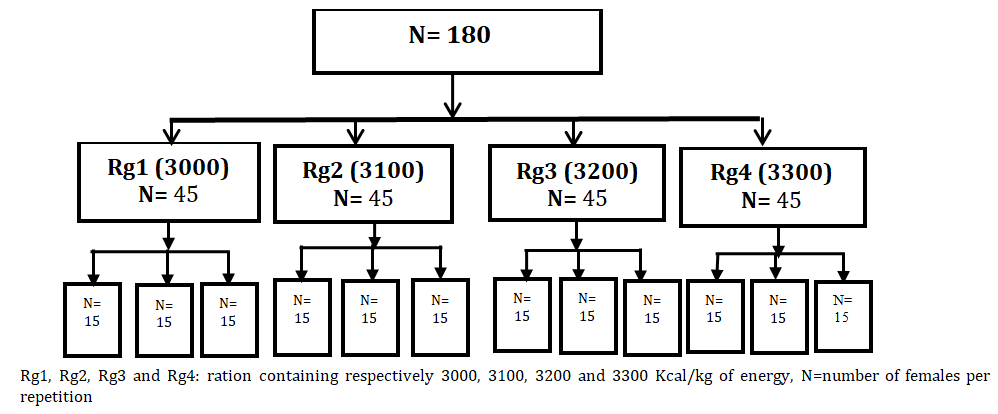
Fish were randomly divided into four comparable groups repeated 3 times with 15 females per repetition (Figure 1). Each group was randomly assigned to one of the experimental rations Rg1 (3000 Kcal/kg), Rg2 (3100 Kcal/ kg), Rg3 (3200 Kcal/kg) and Rg4 (3300 Kcal/kg).
Rg1, Rg2, Rg3 and Rg4: ration containing respectively 3000, 3100, 3200 and 3300 Kcal/kg of energy, N=number of females per repetition.
Conduct of the Test
The trial was conducted in 12 polystyrene happa (like cages) of 1m3, built in a pond of 100m² and depth 0.90 m. This pond was filled with water from a lake located 150m away. The meshes of the happa were cleaned once a week to facilitate the flow of water. The food was distributed twice a day (8 am and 6 pm) in pellet form during 4 months and representing 3% of the ichtyobiomasse. 20% of fish from each group were randomly taken each month, weighed individually using an ichtyometer (0.1cm precision) and SF-
400 balance (1g precision) in order to adjust the amount of food to distribute.
At the end of the test, 12 females presenting the characteristics of sexual maturity (bloated belly containing mature oocytes) from each group were sacrificed.
Biochemical Characteristics
To determine the biochemical characteristics, the pituitary glands (Figure 2a) and the ovaries (Figure 2b)
of the fish sacrificed from each batch at the end of the test were weighed, then crushed in a mortar placed on a piece of ice. The ground materials obtained were mixed with physiological fluid (NaCl) to obtain homogenates 1% for the pituitaries and 15% for the gonads [24]. The different homogenates were centrifuged at 3000 rpm for 30 min and the supernatant obtained was stored at – 20°C in tubes labeled for the determination of FSH (Follicle Stimulating Hormone) and LH (Luteinizing Hormone) in the pituitary glands, estradiol, total proteins, total cholesterol, HDLC (High density lipoprotein cholesterol) and TGS (Triglycerids) in the gonads using the methods described in the instructions for the following kits:
(a)
- The FSH: DIALAB commercial kit (Reference Z05302)
- The LH: DIALAB commercial kit (Reference Z02301)
- Testosterone: GOLDEN BIO TECHNOLOGIES commercial kit (Reference GBE - 2722)
- Estradiol: OMEGA DIAGNOSTICS commercial kit (Reference OD477)
- Total proteins and cholesterol, HDL and tissue TGS: enzymatic colorimetric method following the commercial kit instructions
- LDL (Low density lipoprotein cholesterol) were obtained using the formula below: LDLC = total cholesterol (HDLC + TGS/5)
(b) Figure 2: Pictures of pituitary gland (a) and ovaries (b) of Clarias jaensis.
Reproductive Characteristics
- Absolute fecundity (Af) = Number of oocytes per female
- Relative fertility (Rf) = Number of oocytes per g of female
- Oocyte diameter: measured on graph paper
- Gonado somatic index (GSI) = Gonad weight (g) / Total weight (g) X 100
- Hepatho-somatic index (HSI) = Liver weight (g) / Total weight (g) X 100
Histological Sections of the Gonads
For histological sections, the ovaries of the fish from each previously sacrificed batch were immersed in a fixative (Bouin’s solution) contained in labeled jars, then analyzed in the Animal Physiology laboratory of the University of Yaoundé I, according to the method described by Ngoula F [24].
Statistical Analysis
The data on FSH, LH, estradiol and biochemical characteristics of the ovaries were subjected to one- way ANOVA (energy level). When there were significant differences between the means, they were separated by the Duncan test at 5% significance level. SPSS 20.0 statistical software was used for these analyses.
Results
FSH and LH Concentration
FSH and LH concentrations of the pituitary gland tended to decrease with energy level (Figure 3). FSH was significantly higher (0.81 ± 0.35 mlU/ml) with the Rg2 ration containing 3100 Kcal/kg of energy.
Similarly, LH concentrations were highest (0.66 ± 0.23 mlU/ml) with the Rg2 treatment, but comparable to the other three energy levels. The lowest values (0.33 ± 0.10 mlU/ml) and (0.66 ± 0.23 mlU/ml) for FSH and LH respectively, were obtained with the ration most concentrated in energy Rg4 (3300 Kcal/kg).
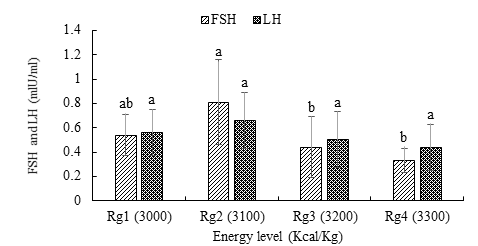
a, b: histograms bearing the same letters are not significantly different (p > 0.05). Figure 3: Effect of the energy level of the ration on the FSH and LH concentrations of the pituitary gland.
Weight of the Ovaries
The weight of the ovaries (Figure 4), although comparable between the different dietary energy levels, was the highest (31.88 ± 6.86g) with the Rg1 treatment (3000 Kcal/kg) and the lowest (27.05 ± 7.64) with the Rg3 ration 3200 Kcal/kg.
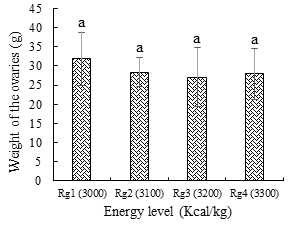
a: histograms bearing the same letters are not significantly different (p > 0.05). Figure 4: Weight of Clarias jaensis ovaries depending on the energy level of the ration.
Estradiol Concentration of the Ovaries
The concentration of estradiol in the ovaries decreased significantly with increasing energy level in the diet (Figure 5). It was the highest (450.00 ± 109.86 pg/ml) with the Rg1 ration containing 3000 Kcal/kg but comparable with the concentration recorded with the energy level 3100 Kcal/ kg (398.89 ± 142. 16 pg/ml). The lowest values (301.25 ± 99.02 pg/ml) were obtained in females fed with the Rg4 ration (3300 Kcal/kg). The relationship between estradiol concentration and energy level was very strong (R² = 0.9996).
a, b, c : histograms bearing the same letters are not significantly different (p > 0.05). Figure 5: Effect of the energy level of the diet on the estradiol concentrations of the ovaries.
Biochemical Characteristics of the Ovaries
The biochemical characteristics of Clarias jaensis ovaries are summarized in Table 2. Total Protein: The significantly highest total protein concentration was obtained with the Rg2 treatment (3100 Kcal/kg) followed by the value observed with the least energy concentrated ration (3000 Kcal/kg). The lowest concentration of total protein was obtained with the most concentrated energy treatment (3300 Kcal/kg) and comparable with the value recorded in batch Rg3 (3200 Kcal/kg). Total Cholesterol: The lowest total ovarian cholesterol concentrations were recorded with the Rg2 ration (3100 Kcal/kg) and were comparable to the values observed in the Rg3 and Rg4 treatments, the highest values being obtained with the lowest level of ovarian cholesterol energy (3000 Kcal/kg). HDLC (HDL density lipoprotein cholesterol): Ovarian HDLC concentrations tend to decrease with increasing energy levels. The significantly highest values were recorded with the Rg1 ration (3000 Kcal/kg). The lowest concentrations (P<0.05) were observed with the Rg2 ration. LDLC (Low density lipoprotein cholesterol): LDLC concentrations tended to increase significantly with increasing energy level. The highest values (P<0.05) were recorded with the Rg3 treatment (3200 Kcal/kg) followed by Rg4 (3300 Kcal/kg). The lowest concentrations were obtained with the energy level 3100 Kcal/kg. TGS (Triglycerids): The highest ovarian TGS concentrations were recorded with the Rg2 treatment (3100 Kcal/kg) followed by the value obtained with the Rg1 ration (3000 Kcal/kg). The lowest concentrations were observed with the treatment at 3300 Kcal/kg. However, no significant differences were recorded between treatments in females.
| Biochemical Characteristics | Energy Level (Kcal/kg) | ||||
|---|---|---|---|---|---|
| Rg1 (3000) | Rg2 (3100) | Rg3 (3200) | Rg4 (3300) | P | |
| T Prot (g/dl) | 7.83 ± 1.67a | 8.00 ± 1.35a | 4.50 ± 1.91b | 4.46 ± 2.09b | 0.00 |
| T Chol (mg/dl) | 59.90 ± 11.66a | 35.12 ± 6.33b | 37.01 ± 11.90b | 38.29 ± 14.28b | 0.04 |
| HDLC (mg/dl) | 79.66 ± 12.13a | 36.54 ± 10.8b | 51.13 ± 8.76b | 51.32 ± 1.87b | 0.01 |
| LDLC (mg/dl) | 79.47 ± 0.41a | 69.32 ± 0.58b | 109.16 ± 0.14c | 84.85 ± 0.20d | 0.00 |
| TGS (mg/dl) | 300.15 ± 67.62 a | 345.24 ± 49.98 a | 220.41 ± 56.38 a | 211.64 ± 54.35 a | 0.12 |
Table 2: Biochemical characteristics of Clarias jaensis ovaries depending on the energy level of the diet.
a, b and c: means with the same letter on the same line are not significantly different (P>0.05) for the same parameter. N=12, T prot= Total proteins, T Chol = Total cholesterol, HDLC = High density lipoprotein cholesterol, LDLC = Low density lipoprotein cholesterol, TGS = Triglycerids, P = probability Table 2: Biochemical characteristics of Clarias jaensis ovaries depending on the energy level of the diet.
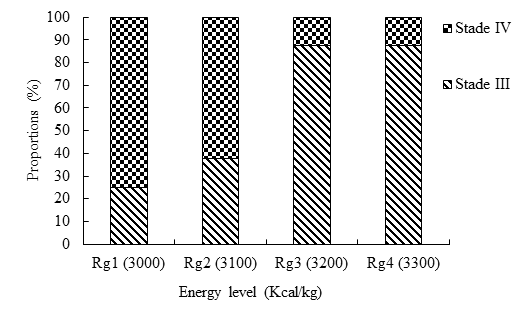
Stages of Sexual Maturity of Females
Females of stages III and IV of sexual maturity are found in all treatments (Figure 6). Thus, stage IV was the most represented with rates of 75 and 62.5% respectively in treatments Rg1 (3000 Kcal/kg) and Rg2 (3100 Kcal/kg). The females having received the most concentrated energy rations (3200 and 3300 Kcal/kg) were mainly (87.5%) at stage III of sexual maturity.
Fecundity (Absolute and Relative), Oocyte Diameter and Weight
The effect of the energy level of the ration on absolute fertility, relative fertility, oocyte weight and diameter is summarized in Table 3. It appears that these characteristics tended to decrease with the increasing level of energy of the ration except relative fertility which has rather increased.
Absolute Fertility and Relative Fertility
Absolute fertility (Af) tended to decrease with increasing energy levels. The highest value was recorded with 3000 Kcal/ kg of energy followed by the value obtained with the highest level of energy (3300 Kcal/kg), the lowest being obtained with the Rg3 ration (3200 Kcal/kg). Relative fertility (Rf), on the other hand, increased with increasing energy levels and the highest value was recorded with the highest energy level (3300 Kcal/kg). It was lower with the Rg1 ration (3000 Kcal/kg). However, no significant difference was observed between treatments for either absolute fertility or relative fertility.
Oocyte Weight and Diameter
The increase of the energy level in the ration tends to reduce oocyte weight and diameter. The highest values were recorded with the ration with the least concentrated energy (3000 Kcal/kg) and the lowest with the three other treatments in terms of oocyte weight and the Rg2 treatment (3100 Kcal/kg) for the diameter. However, the treatments showed no significant difference for either weight or oocyte diameter (P>0.05).
| Oocytes Characteristics N=12 | Energy level (Kcal/kg) | ||||
|---|---|---|---|---|---|
| Rg1 (3000) | Rg2 (3100) | Rg3 (3200) | Rg4 (3300) | P | |
| Af | 7234.68 ± 1730.98 | 6887.37 ± 275.67 | 6819.06 ± 1690.16 | 7027.89 ± 1504.84 | 0.938 |
| Rf | 35.00 ± 8.93 | 36.21 ± 3.33 | 36.78 ± 8.14 | 40.09 ± 7.52 | 0.579 |
| Oocytes weight (g) | 0.005 ± 0.001 | 0.004 ± 0.001 | 0.004 ± 0.001 | 0.004 ± 0.001 | 0.656 |
| Oocytes diameter (mm) | 0.66 ± 0.04 | 0.60 ± 0.35 | 0.63 ± 0.05 | 0.62 ± 0.04 | 0.326 |
Table 3: Oocyte characteristics of Clarias jaensis according to energy level.
Af = Absolute fertility, Rf = Relative fertility, P = probability, N = number of observations Table 3: Oocyte characteristics of Clarias jaensis according to energy level.
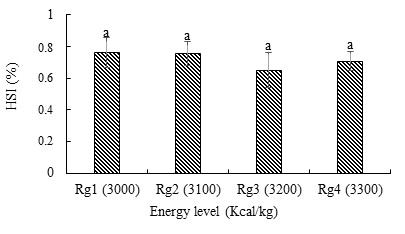
Gonado Somatic Index (GSI) and Hepato Somatic Index
The GSI Figure 7a was highest (15.18 ± 1.99%) in the Rg1 ration, with the lowest value (14.38 ± 3.11%) recorded in the Rg3 treatment. The hepato-somatic index Figure
7b was higher (0.076 ± 0.09%) in females with the Rg1 ration containing 3000 Kcal/kg of energy and lower (0.70 ± 0.06%) with the Rg3 treatment (3200 Kcal/kg). However, no significant differences were recorded between the treatments. However, no significant difference was noted between the treatment.
a,b : histograms bearing the same letters are not significantly different (p > 0.05) Figure 7: Gonado Somatic Index (a) and Hepato somatic Index (b) depending on the energy level of the ration Rapport Hépatho Somatique (RHS).
Histology of the Ovaries
The effect of the energy level of the ration on the structure of the ovaries of Clarias jaensis females fed with rations at 3000, 3100, 3200 and 3300 Kcal/kg of energy is illustrated respectively by photos a, b, c, d and e of Figure 8. Regardless of the energy level considered, females presented mature oocytes at stage III and IV whose lumen is filled with yolk. A few atretic oocytes were observed in the batch that received the most dietary energy (Figure 8e).
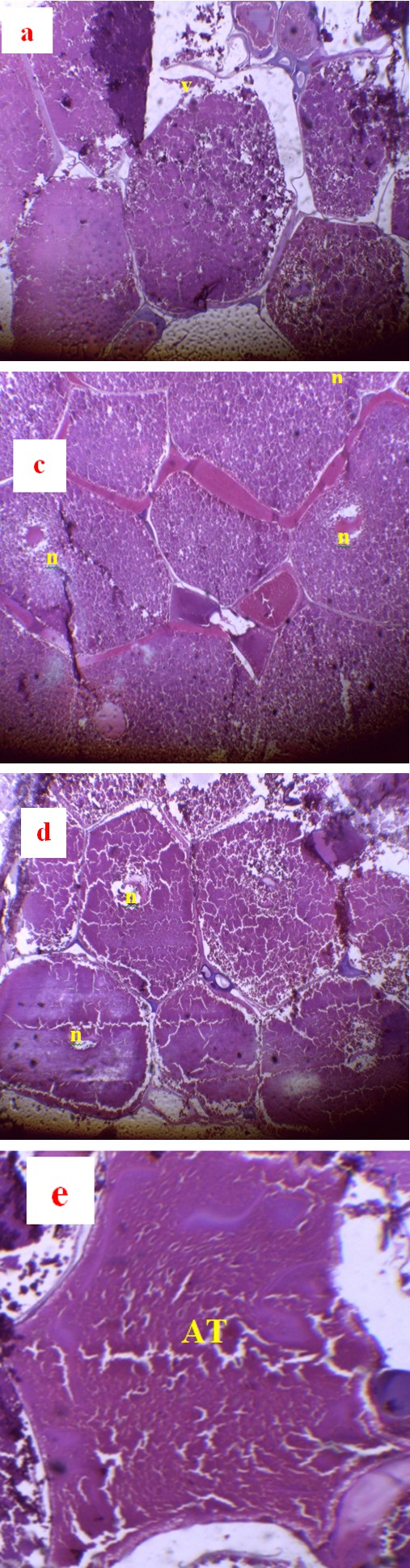
n=nucleus, v=yolk, AT=atresia a, b, c and d present the oocytes of females who received rations containing 3000, 3100, 3200 and 3300 Kcal/kg of energy respectively. (e)atretic oocyte present in females fed at 3300 Kcal/kg. Figure 8: Histological sections of the ovaries of Clarias jaensis fed with different levels of food energy.
Discussion
Reproduction consumes energy that the animal obtains from its food. Fish that do not have sufficient mobilizable energy reserves cannot reach sexual maturity [19]. FSH and LH are hormones involved in gonad development, steroidogenesis and ovulation [25]. The FSH and LH concentrations of females were highest with the 3100 Kcal/ kg energy ration and lowest with the 3000 Kcal/kg energy ration. However, the concentration of estradiol was higher with the 3000 Kcal/kg ration although those of FSH and LH were low in this batch. This observation could be explained by negative feedback exerted by estradiol on the pituitary gland, which would have led to a reduction in gonadotropin secretion. Furthermore, regardless of energy level, estradiol concentrations were low compared to those recorded by Sabet SS, et al. [26] in the cyprinidae (Rutilis frisii kutum) and by Cornish in Oreochromis mosambicus in the natural environment. The difference observed between these results would be linked to the species, environmental factors and breeding techniques.
The highest values of total protein concentration in the ovaries were observed with the ration at 3100 Kcal/kg followed by those at 3000 Kcal/kg (p < 0.05). We do not have enough information on normal protein values in fish gonads. However, in fish plasma, these values are in the range 3-5 g/ dl [27]. The values obtained in the ovaries of Clarias jaensis were lower than those observed in Clarias batrachus which had not been deprived of food [28]. Low energy levels would have led to strong mobilization of proteins into lipoproteins during gonad maturation [22]. Indeed, protein concentrations increase significantly during the sexual maturation process of fish [27].
According to the work carried out by Barnhart RA [29], the main variation factors for total cholesterol concentrations are species and nutrition. In our experience, total cholesterol varied significantly with energy level, the highest value being recorded with the Rg1 treatment (3000 Kcal/kg) as was estradiol. Females fed at 3000 Kcal/kg would have mobilized more cholesterol which was not entirely used for the production of estradiol. The low concentrations of LDLC in the ovaries of females from the same batch would be linked to their high use in the transport of estradiol. Cholesterol is the major component present in animal tissue [30]. It is modified by an enzymatic system to produce steroid hormones in tissues such as the brain, adrenal glands, ovaries and testes [30, 31]. Note that the concentrations of protein, cholesterol and estradiol contained in the gonads of females from the group fed at 3000 Kcal/kg were the highest, which shows that they were more mature and ready to reproduce. According to Yeganeh S [27], these three characteristics increase significantly during sexual maturity.
HDLC concentrations in the ovaries of females from the group fed the low-energy diet were the highest. This result could be explained by the fact that the high concentration of estradiol in this same batch would have stimulated a strong production of HDLC which was used by the growing oocytes for the synthesis of the yolk.
Triglyceride concentrations tended to decrease with increasing energy level, however, no significant difference was recorded between treatments. However, females from the groups that received the least energy recorded the highest values. These females would have stored energy useful for reproduction in the form of triglycerides. Indeed, according to Sujatha K [32], TGS are stored in the body to be used as an energy source.
In female fish, the average number of oocytes is considered an important criterion for evaluating reproductive performance [33]. The fecundities obtained during this work were not significantly affected by the energy level. This result is different from that observed by El-Sayed FM, et al. [34] in Oreochromis niloticus. The differences observed would be due to endogenous and exogenous factors such as age, species, physiological state and rearing conditions. However, absolute fertility was highest with the ration at 3000 Kcal/ kg and relative fertility with that at 3300 Kcal/kg of energy. These results are close to those recorded in Clarias jaensis induced with HCG at 4000 IU and captured in a natural environment [12, 35].
Oocyte diameter and weight help determine egg quality. As well as absolute and relative fecundity, oocyte weight and diameter were highest with the ration with the least energy concentration (3000 Kcal/kg).
Whatever the ration considered, the oocyte diameter was lower than that reported by Zango P [12] in Clarias jaensis in a natural environment and the oocyte weight was lower than that recorded in Clariidaes by Le gouessant [36]. The difference observed could be explained by a variation in the size, weight and age of the females. The absolute fecundity of females was the highest with the ration at 3000 Kcal/kg as well as the live weight. This result is similar to that of Alla YL, et al. [37] who reported that the more the size and live weight of Heterobranchus longifilis female increase, the higher its fecundity. The fecundity, oocyte diameter and gonad weight recorded in this study show that females fed at 3000 Kcal/kg are more mature than those in other batches. The high estradiol concentrations in females fed rations containing low energy levels would have stimulated high oocyte production and maturation.
The gonado somatic index (GSI) is a characteristic that accounts for the progression of gametogenesis in fish. A high GSI at a period indicates that the gonads are mature at that period and the broodstock are ready to reproduce [38, 39, 40]. The GSI recorded during this study was not significantly affected by energy level. However, it was highest with the ration containing the least energy (3000 Kcal/kg). Çek S, et al. [33] and Reidel A, et al. [41] recorded similar results in females of Clarias gariepinus and Rhamdia quelen. These results are also similar to those recorded in females of Oncorhyncus mikiss fed three foods, containing different energy levels [42]. Estradiol and GSI were highest with the diet containing the lowest energy level. Several studies have shown that the increase in GSI and the development of oocytes are associated with the level of estradiol in plasma [38, 40]. The increase in GSI indicates that oocyte maturation progresses with increasing estradiol levels [43, 44].
The hepatho-somatic index (HSI) varied significantly with energy level. It generally decreased with increasing energy level and was highest with the 3000 Kcal/kg. Çek S, et al. [33] recorded similar results in Clarias gairepinus fed rations with different digestible energy levels. Lincoln RF, et al. [45] reported that the HSI is high when the liver is engaged in the synthesis of vitellogenin, this could explain the presence of a high quantity and quality of oocytes with the 3000 Kcal/kg ration in Clarias jaensis. However, the ration containing the lowest energy level recorded the highest GSI and HSI in females. These results do not corroborate those reported by Bedoui RF, et al. [46] who found that in the natural environment, the RGS of liza aurata increases while the RHS decreases. However, they are similar to the results recorded in Upeneus moluccensis by Saad A [47] who found that the two relationships evolved synchronously.
According to the other reproductive characteristics cited above, females fed at 3000 and 3100 Kcal/kg of energy were classified as mature compared to those having received higher energy levels. These results are similar to those reported by Çek S, et al. [33] who obtained mature broodstock of Clarias gariepinus with rations containing the lowest levels of digestible energy. These results are confirmed by examination of histological sections of the gonads. Whatever the energy level, the histological sections of the ovaries presented oocytes whose cytoplasm was filled with yolk. Some atretic oocytes were recorded with the ration containing the most energy (3300 Kcal/kg). This result is similar to that of Çek S, et al. [33] in Clarias gariepinus and Reidel A, et al. [41] in Rhamdia quelen. These authors explain that excess energy could have caused follicle atresia by affecting the metabolic activities of the liver, which is the basis of vitellogenin synthesis linked to the growth of vitellogenic follicles. According to Leonardo AFG, et al. [48] atresia is due to the absence of hormonal induction at the time of ovulation, which leads to a reduction in the volume of oocytes, an increase in white oocytes and their reabsorption by phagocytosis. Babin P, et al. [49] rather think that atresia is caused by environmental stress and changes in hormonal concentrations.
Last and not a least explanation, palm oil contains less unsaturated fatty acids. Since overall need in omega 3 and 6 in fish nutrition is well established, using alternate lipid sources could have probably provided more challenging results [50].
Conclusion
Energy level significantly affected reproductive characteristics except LH, ovarian weight, triglycerides, oocyte characteristics, GSI, HSI and K factor. FSH and LH concentrations were higher with the 3100 Kcal/kg treatment while other reproductive characteristics were generally better with the 3000 Kcal/kg treatment. Using alternate lipid source than palm oil for dietary energy may be tried.
Acknowledgements
We are grateful to Mr Louis Enoc and Raphael Ewane, the local fisherman for donating the post fingerlings of Clarias jaensis required for this research work.
Conflicts of Interest
The authors declare no conflict of interest
References
-
FAO (2022) The global situation of fisheries and aquaculture. Towards a blue transformation. FAO, Italy, pp: 294.
-
Fostier A, Jalabert B (2004) Domestication and reproduction in fish. INRA Productions Animales 17(3): 17-30.
-
Lazard J (2017) Aquaculture systems facing climate change. Cahiers Agricultures 26(3): 34001.
-
Teugels GG, Adrians D (2003) Taxonomy and phylogeny of claridae. An oevrview. Phylogeny, USA 1: 466-487.
-
Diogo R (2005) Morphological evolution, Adaptations, Homophasies, constraints and Evolutionary Trends: Catfishes as a case study on general phylogeny and Macroevolution. Science Publishers, USA, pp: 491.
-
Ducarme C, Micha JC (2003) Intensive production technique for African catfish. _Clarias gariepinus_. Tropicultura 21(4): 189-198
-
Lacroix (2004) Fish farming in tropical areas. GFA Terra Systems, Germany, pp: 231.
-
Rukera TS, Micha JC, Ducarme C (2005) Adaptation trials for mass production of _Clarias gariepinus_ juveniles in rural conditions. Tropicultura 23(4): 231-244.
-
Lazard J, Lecomœ Y, Stomal B, Weigel JY (1991) Fish farming in Sub-Saharan Africa: Situations and projects in French-speaking countries proposals for action. Ministry of Cooperation and Development, pp: 158.
-
Efole ET (2011) Biotechnical optimization of fish farming in ponds as part of the sustainable development of Family Farms in Cameroon. Doctoral thesis in Agronomic, Agri- food, Horticultural and Landscape Sciences (fisheries). European University of Brittany, pp: 215.
-
Pouomogne V (2008) Capture-based aquaculture of Clarias catfish: case study of the Santchou fishers in western Cameroon. In: Lovatelli A, Holthus PF (Eds.), Capture-based aquaculture. Global overview. FAO Fisheries Technical Paper, Italy 508: 93-108.
-
Zango P (2009) Reproductive characteristics compared in the natural environment of _Clarias gariepinus_ and _Clarias jaensis_. University of Dschang. Master of Sciences thesis, specialty: Aquaculture pp: 98.
-
Mfossa MD (2012) Promoting local know-how in fish farming practices: case of traditional ponds in the Santchou plain (West Cameroon). European academics pp: 77.
-
Zango P, Tomedi ETM, Efole TE, Tiogue CT, Efole TE, et al. (2017) Comparative survival and growth performances of Oreochromis niloticus associated with _Clarias jaensis_ and _Clarias gariepinus_. Cameroon Forum for Biological Sciences 11(1): 1-8.
-
Etaba AD, Tomedi EM, Djoko H, Tchoumboue J (2016) Growth performance of African catfish Clarias jaensis Boulanger, 1909 (Pisces: Clariidae) in ponds fertilized with chicken droppings and pig manure. International Journal of Innovation and Applied Studies 17(4): 1294- 1301.
-
Efole ET, Tchouante TCG, Miegoue E, Tsoupou KSG, Nana TA, et al. (2016) Effect of Level of Desmodium Uncinatum incorporation in the Diet on Growth Performance of _Clarias jaensis_ (Boulenger, 1909) in the Highlands of west Cameroon. Journal of Agriculture and Veterinary Science 9(12): 74-82.
-
Smith CE, Osborne MD, Piper RG, Dwyer WP (1979) Effect of diet composition on performance of rainbow trout broodstock during a three year period. Progressive fish culturist 41(4): 185-188.
-
Fernandez-Palacios H, Izquierdo MS, Robaina L, Valencia A, Salhi M, et al. (1997) The effect of dietary protein and lipid from squid and fish meals on egg quality of broodstock for gilthead seabream (_Sparus aurata_). Aquaculture 148(2-3): 233-246.
-
Barnabe G (1991) Biological and ecological bases of aquaculture. Lavoisier pp: 520.
-
Tsoupou KSG, Kpoumie NA, Tonfack AP, Komguep NR, Tchouante TCG, et al. (2023) Survival, Growth Response, Chemical and Biochemical Characteristics of the Carcass of _Clarias jaensis_ (Boulenger, 1909) Post Fingerlings Fed Various Dietary Energy. J Aquac Res Dev 14(7): 777.
-
Komguep NR, Efole TE, Defang FH, Nana TA, Mube KH, et al. (2021) Effects of Dietary Protein Level on Growth Performance of Pre-growing Catfish _Clarias jaensis_ (Boulanger 1909) in Captivity. J Aquac Res Dev 12: 658.
-
Tsoupou KSG, Efole ET, Tchoumboue (2018) Reproductive performance of male broodstock Clarias jaensis (Boulenger, 1909) fed various dietary energy. International Journal of Fisheries and Aquatic Studies 6(4): 264-270.
-
AOAC (2000) Official methods of analysis. Association of Official Analytical Chemists, USA.
-
Ngoula F (2008) Effects of organophosphate and carbamate pesticides on some reproductive parameters in adult male rats. Animal Physiology, University of Yaounde I pp: 200.
-
Yadetie F, Male R (2002) Effect of 4-nonylphenol on gene expression of pituitary hormones in juvenile atlantic salmon. Aquatic toxicology 58(1-2): 113-129.
-
Sabet SS, Mohammad RI, Bagher AF, Saeed G (2016) Variations in Plasma Sex Steroid Hormones of the Wild Caspian Cyprinid Fish, Kutum (_Rutilus frisii_ Kutum). Journal of Cell and Molecular Research 8(1): 25-34.
-
Yeganeh S (2011) Seasonal changes of blood serum biochemistry in relation to sexual maturation of female common carp (_Cyprinus carpio_). Comparative Clinical Pathology pp: 5.
-
Prasad KN (2015) Effects of Prolonged Starvation on Cholesterol Content of Gonads in _Clarias batrachus_. Our Nature 13(1): 26-30.
-
Barnhart RA (1969) Effects of certain variables on hematological characteristics of rainbow trout. Trans Am Fish Soc 98(3): 411-501.
-
Gower DB (1988) The biosynthesis of steroid hormones. Elsevier Science Publishers 18: 3-28.
-
Harris M (1985) Endocrine Physiology. Quarterly Journal of Experimental Physiology 71(3): 493-494.
-
Sujatha K, Joice AA, Senthilkumaar P (2013) Total protein and lipid content in edible tissues of fishes from Kasimodu fish landing centre, Chennai, Tamilnadu. European Journal of Experimental Biology 3(5) : 252- 257.
-
Çek S, Yilmaz E (2009) The effect of varying dietary energy on gonad development at first sexual maturity of the Sharptooth catfish (_Clarias gariepinus_ Burchell, 1822). Aquaculture International 17(6): 553-563.
-
El-Sayed FM, Mansour CR, Ezzat AF (2005) Effect of dietary lipid source on spawning performance of Nile tilapia (_Oreochromis niloticus_) broodstock reared at different water salinities. Aquaculture 248(1-4): 187- 196.
-
Zango P, Tomedi ETM., Efole TE, Tiogue CT, Nguenga D, et al. (2016) Performances de reproduction du poisson chat endogène du Cameroun _Clarias jaensis_ (Boulenger, 1909) in a controlled environment. International Journal of Biology and Chemical Sciences 10(2): 533-542.
-
Le gouessant (2009) The _Clarias gariepinus_ case. High quality fish seed and feed production are the basis for developing the aquaculture industry. Technical special issue, pp: 16.
-
Alla YL, Ouattara M, Blé MC, Atse BC (2010) Study of the fertility of the African catfish Heterobranchus bidorsalis (Geoffroy Saint-Hilaaire, 1840) under breeding conditions. Tropicultura 28(1): 31-36.
-
Heidari B, Roozati S, Yavari L (2010) Changes in plasma levels of steroid hormones during oocyte development of Caspian Kutum (_Rutilus frisii_ kutum, Kamensky, 1901). Animal Reproduction 7(4): 373-381.
-
Abdolhay HA, Daud SK, Ghilkolahi SR, Pourkazemi M, Siraj SS, et al. (2010) Fingerling production and stock enhancement of Mahisefid (_Rutilus frisii_ kutum) lessons for others in the south of Caspian Sea. Review Fish Biology and Fisheries 21(2): 247-257.
-
Taghizadeh V, Imanpoor MR, Mehdinejad N (2013) Study the seasonal steroid hormones of common carp in Caspian Sea, Iran. Springer Plus 2: 193.
-
Reidel A, Wilson RB, Aldi F, Elizabeth R (2010) The effect of diets with different levels of protein and energy on the process of final maturation of the gametes of _Rhamdia_ _quelen_ stocked in cages. Aquaculture 298(3-4): 354-359.
-
Memis D, Gün H (2004) Effects of Different Diets on the Growth Performance, Gonad Development and Body Composition at First Sexual Maturity of Rainbow Trout (_Oncorhynchus mykiss_). Turkey journal of veterinary animal sciences 28(2): 315-322.
-
Rinchard J, Kestemont P (1996) Comparative study of reproductive biology in single-and multiple-spawner cyprinid fish. I. Morphological and histological features. Journal of Fish Biology 49(5): 883-894.
-
Adebiyi FA, Siraj SS, Harmin SA Christianus A (2013) Plasma sex steroid hormonal profile and gonad histology during the annual reproductive cycle of river catfish _Hemibagrus nemurus_ (Valenciennes, 1840) in captivity. Fish Physiology and Biochemistry 39(3): 547-557.
-
Lincoln RF, Scott P (1984) Sexual Maturation in Triploid Rainbow Trout, _Salmo gairdneri_, Richardson. Journal of Fish Biology 25(4): 385-392.
-
Bedoui RF, Gharbi H, EL Abed A (2002) Reproductive period and sexual maturity of _Liza aurata_ (Mugilidae fish) from the eastern and southern Tunisian coasts. Bulletin de l’INSTM 29: 11-15.
-
Saad A (1998) Reproductive cycle and fecundity of Upeneus moluccensis (Blecker, 1855). Indi-pacific species in Syrian sea waters (Eastern Mediterranean sea). In: Leonart J (Ed.), Marine popultions dynamics. CIHEAM, Spain, pp: 87-92.
-
Leonardo AFG, Romagosa E, Batlouni SR, Borella MI (2006) Occurrence and significance of ovarian and follicullar regression in cachara _Pseudoplatystoma_ _fasciatum_ (Linnaeus, 1766): a histology approach. Arq Bras Med Veterinária e Zootecnica 58(5): 831-840.
-
Babin P, Cerda J, Lubzens E (2007) The Fish Oocyte: From Basic Studies to Biotechnological Applications. In: 2nd (Edn.), Springer, pp: 508.
-
National Research Council (2011) Nutritional requirements of fish and shrimps. Agricultural Research Service, USA 20001(800): 624-6242.
- Genetic Improvement of Nile Tilapia (Oreochromis niloticus): Advances in Selective Breeding and Genomic Approaches for Sustainable Aquaculture
- Microplastics, Contaminants, and Waste Hotspots: Divergences and Faults in Prioritizing Control Efforts
- Creating a Healthier, More Vibrant Open and Closed Aquatic Environment. A Submersible, Centrifugal Magnetically Affixed Current Changing Aquarium Pump
- An Attempt to Assess Alpha Diversity and Sample Size: Using the Ostracod Assemblages off Kumamoto Port, Japan
- Assessment of the Efficiency of Common Fishing Gears and Crafts Used at Mohananda River of Chapai Nawabganj, Bangladesh
- Fish Productivity and Biodiversity Status of Sundarban Mangrove in Bangladesh