Zoonotic Hemoparasites of Baboons (Papio Anubis) at the Human-Wildlife Interface in Kenya
In Kenya, the encroachment of baboon habitats has led to increased interaction between humans and non-human primates (NHP). This interaction may result in negative consequences such as increased chances of zoonotic parasite transmission. The current study investigated the protozoan hemoparasites (both zoonotic and non-zoonotic) of free ranging baboons (Papio anubis) at two human–baboon interface sites: Tsavo West National Park (TWNP) and Tana River Primate Reserve (TRPR). One hundred and six baboons were trapped, anaesthetized, physically examined, and sampled for blood and ectoparasites. Physical examination revealed lymphadenopathy in all (100%) baboons from Tsavo and 66.7% in Tana. Microscopic examination of blood smears indicated the following prevalence of hemoparasites in TWNP and TRPR baboons respectively; Hepatocystis kochi (70%, 64.4%), Entopolypoides macaci (0%, 4.4%), Babesia microti (0%, 4.4%), and Plasmodium spp. (1.7%, 0%). PCR screening results revealed slightly higher prevalence of hemoparasites in TWNP and TRPR baboons respectively: Hepatocystis kochi (87%, 90%), Babesia (10.8%, 16.7%) and Entopolypoides (8.7%, 5%). There was a significant difference in the prevalences of Entopolypoides macaci and Babesia microti between the two sites (P
Introduction
Wildlife diseases are a major concern to humans because most of them can be transmitted to domestic animals as well as humans [1]. They also form a sizeable proportion of emerging and re-emerging diseases which can threaten wildlife populations by altering species abundance and population structures [2, 3]. Human diseases of animal origin, also known as zoonoses, comprise more than 150 infections of various etiologies. Introduction of an animal pathogen into the human population and the subsequent extension or alteration of its host range leading to the successful maintenance of the corresponding pathogen by human-to-human transmission possesses a serious risk for world-wide health care [4]. The occurrence of a wide spectrum of zoonoses in non-human primates (NHPs) makes them a leading source of emerging and re-emerging diseases, hence posing a major challenge for the public and international health systems [5]. Scientific preparedness is therefore a prerequisite to finding appropriate solutions for these challenges [6].
NHPs are most likely to act as reservoirs of various zoonotic parasites (e.g. protozoa, bacteria and virus) and the possibilities of transmission of these parasites to humans is ever present [7, 8]. The bi-directional transmission of a variety of zoonotic parasites between humans and NHPs often occurs because of morphological, physiological, genetic and behavioral similarities between humans and NHPs [9]. Some of these zoonotic parasites are transmitted by vectors such as ticks and blood sucking dipteran flies which abound at the human-wildlife interfaces [10]. Because this study focused on hemoparasites of NHPs, we provide a background on zoonotic protozoal parasites. NHPs are infected with a number of zoonotic protozoal parasites, including toxoplasmosis, Chagas' disease, babesiosis, giardiasis, and leishmaniasis, can cause insidious infections, with asymptomatic animals being capable of transmitting disease [7].
The presence of Babesia microti (a zoonotic hemoprotozoan) infection has been reported in a few populations of free ranging baboons in Kenya [2, 8]. Babesiosis is a tick-transmitted disease caused by intraerythrocytic parasites of the genus Babesia. Babesial parasites require both a competent vertebrate (mammal e.g. man, domestic or wild animals) and invertebrate (tick) host to maintain transmission cycle. Because babesial parasites are capable of infecting various species of mammals, the awareness of their role as zoonotic agents of human disease and emerging tick-borne diseases is growing [10]. In domestic animals, Babesia divergens, a parasite of cattle, has been implicated as the most common agent of human babesiosis in Europe, causing severe disease in splenectomized individuals. In the US, Babesia microti, a babesial parasite of small mammals, has been the cause of over 300 cases of human babesiosis since 1969, resulting in mild to severe disease, even in non-splenectomized patients. This parasite is closely related to babesial parasites isolated from large wild ungulates in California [11]. In NHPs, Babesia microti infection is enzootic (constantly present in an animal community but in a small number of cases) and is often characterized by fatal haemolytic crisis in immune- compromised baboons [12].
Babesia-like parasites of the genus Entopolypoides macaci have also been reported to infect nonhuman primates; based on phylogenetic analysis of small-subunit rRNA (SSUrRNA) sequences of this parasite and on serological and epidemiological data. Bronsdon, et al. [12] suggest that the genus Entopolypoides is synonymous with that of Babesia. In primate centers, natural infections with this parasite have been shown in baboons (Papio cynocephalus) [12] long-tailed macaques (Macaca fascicularis), and rhesus macaques (Macaca mulatta), providing animal models in species closely related to humans to study parasite–host relationships of this pathogen [13, 14]. Brondson, et al. [12] report that high parasitemia can result in acute anemia in immune compromised or splenectomized baboons. Phylogenetic analysis of nuclear small-subunit rRNA gene sequences amplified from peripheral blood of a baboon chronically infected with E. macaci demonstrated this parasite to be most closely related to Babesia microti (97.9% sequence similarity); sera from infected animals did not react in indirect fluorescent-antibody tests with Babesia microti antigen, however, suggesting that they represent different species [12].
Habitat Encroachment and Potential Transmission of Protozoal Zoonoses
In Kenya, the rapid increase in human population and mobility of wildlife has dramatically increased the numbers of people interacting with non-human primates (NHPs). Human encroachment in search of agricultural land has in recent decades shifted to low potential rangelands which are the prime wildlife ecosystems [15]. This has created a myriad of problems like competition for resources, human-wildlife conflicts and habitat fragmentation [16]. The situation is compounded by the ever increasing bush-meat trade where NHPs are trapped and butchered by several communities in Africa [15].
Various studies have shown that NHPs have high interaction with humans hence making them likely agents of zoonoses transmission [17]. For example, a recent study by Mwangi, et al. [18] at one of our study sites found that NHPs including baboons, vervets, and sykes engaged in crop raiding and predation of domestic animals. The occurrence of these parasites in habitats where there is close human-NHP interaction needs to be examined to determine the extent to which human beings are exposed to zoonoses. The current study aimed at determining 1) The prevalence of zoonotic hemoparasites in free ranging baboons at human wildlife interface 2) The presence of potential vectors that transmit zoonoses and 3) The consequences of hemoparasite infection in these NHPs. Results from this study will create awareness to the people inhabiting these locations and also to the government for appropriate control measures to be put in place to avoid serious outbreaks as has been the case with other emerging diseases.
Materials and Methods
Ethical Approval
The study was approved by the Institute of Primate Research’s Animal Care and Use Committee (ACUC), a sub-committee of the Institutional Review Committee (approval number IRC/15/11). The research adhered to the legal requirements of the Kenya. Approval for capturing and sampling of baboons in the wild was also obtained from Kenya Wildlife Service, the government body in charge of wildlife in the Kenya.
Study Area
Our first study site was Nthongoni area which lies in Mtito Andei division on the southern part of Makueni district, Kenya. It borders Tsavo West National Park (TWNP) to the south and Chyulu national park to the North West and lies between N 1° 35’ S and 37° 10’ E and at an altitude of 600-1000m above sea level. The habitat is quite diverse and natural. The area has a bimodal rainfall pattern with short rains occurring between November and December and long rains between March and April [19]. Temperatures are high ranging between 20°C and 30°C [19]. The second study site was Tana River Primate Reserve (TRPR) which is located in Tana River County and supports a high diversity of plant and animal species.
It is located 100 km upstream from the delta and lies latitudes 2°15’ and 1°50’ south. The climate is hot and dry and rainfall ranges between 500 and 600mm per year and temperatures between 20 and 40°C. The area has patched of riverine forests, dry woodland and savanna habitats [20].
Field Trapping and Physical Examination
One hundred and six (106) baboons were trapped in TWNP (46) during November, 2011 and TRPR (60) during April, 2012. The baboons were captured as described by Moinde, et al. [21]. The baboons were habituated, then trapped before being anaesthetized using Xylazine / Ketamine mixture at the ratio of 20:1 respectively at a dose rate of 10mg/kg (0.5mg/kg and of Ketamine and 9.5mg/kg of Xylazine). Once under deep sedation, the animals were shaved on a small area on the limbs to avoid re-sampling in case of re-capture. The animals were physically examined, and clinical parameters, skin lesions and presence of ectoparasites were recorded. Blood samples were collected as described below. The animals were then observed for recovery before being released back to the wild. For subjects with slow recovery rates, the anesthesia was reversed using Atipamazole hydrochloride (Atepam®, Cipla ltd, India) at 0.5 mg/kg body weight to hasten recovery. Data on location, sex and age were recorded, we trapped both males and females and the animal ages ranged from juveniles to adults.
Sampling of Blood, Hematology and Microscopic Screening of Hemoparasites
For each baboon, a blood sample (4ml) was collected through venipuncture of the femoral vein into vacationers containing Ethylenediaminetetraacetic acid (EDTA) as anticoagulant. The blood was placed in a cool box and transported to laboratories at Institute of Primate Research. Thin blood smears were prepared, air dried, stained with absolute methanol for 5 minutes, rinsed with tap water then stained with 10% Giemsa for 20 minutes, then excess stain washed off with tap water again, dried and examined for presence of hemoparasites as described by Maamun, et al. [8]. Further, the smears were used to determine the differential leucocyte counts. Parasitemia was determined by counting at least 2000 red blood cells (RBC); erythrocytes from a quarter of each field were counted then multiplied by four. The number of infected erythrocytes was counted within the 2000 red blood cells. Percentage parasitemia was then calculated as = total counted parasites divided by number of erythrocytes counted x 100. The rest of the blood was frozen at -20°C
and was used for molecular screening of hemoparasites as described below.
Molecular Screening of Hemoparasites by Polymerase Chain Reaction (PCR)
DNA extraction was carried out on whole EDTA blood using Quick-gDNATM MiniPrep kit (Zymo Research Corporation) as per manufacturer’s instructions and then stored at -20°C. The master mix was prepared in 2ml thin walled PCR tubes for each parasite as follows; for Hepatocystis kochi, it was composed of 17.65 µl PCR water, 2.5 µl of 10x buffer, 1.25 µl of 50nM MgCl2, 0.5 µl of 10nM dNTPs, 1.25 µl each for forward primer and reverse primer, 0.1 µl Taq polymerase and 0.5 µl the DNA sample. For Babesia microti, it was made of 17.35 µl PCR water, 2.5 µl of 10x buffer, 1.5 µl of 25nM MgCl2, 0.5 µl of 20nM dNTPs, 0.25 µl each for forward and reverse primer, 0.15 µl Taq polymerase and finally 0.5 µl DNA sample. For Entopolypoides macaci, the master mix had 16.85 µl PCR water, 2.5 µl of 10x buffer, 2 µl of 25nM MgCl2, 0.5 µl of 10nM dNTPs and 0.25 µl each of forward and reverse primers, 0.15 µl of Taq polymerase and 2.5 µl of the sample DNA. The master mix was vortexed the taken for amplification (Appendix 1).
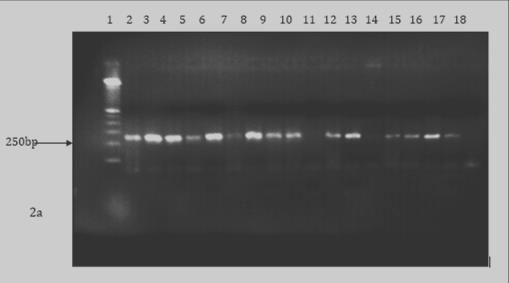
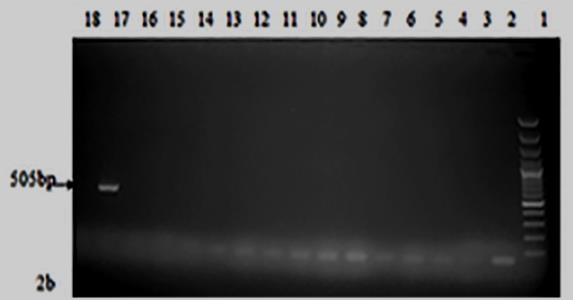
The parasite-specific DNA were then amplified using specific primer sets for Hepatocystis kochi (F-5'- CATTTACACGGTAGCACTAATCCTT-3' and R-5'- GGAATGGTTTTCAACATTGCAT-3'), Entopolypoides macaci (F- 5’ ATACAGCGAAACTGCGAATG 3', R 5' GAAGGGTTTAGATCCCCATCA 3’), Babesia microti (F 5' C TGCGGCTTAATTTGACTC-3', R-5'- GGATCACTCGATCGGTAGGA-3') (25) (Appendix 2). PCR amplifications were done on a PTC-200 Peltier Thermocycler. The amplification conditions for H. kochi were 95°C for 2 minutes of initial denaturation, denaturation was at 95°C for 45 seconds, primer annealing at 55°C for 30seconds and extension at 68°C for one minute with the three later steps going through 39 cycles. Final extension conditions were at 72°C for 5 minutes. Babesia microti initial denaturation was at 95° C for 5 minutes followed by denaturation at 95° C for 30 seconds then primer annealing at 59°C for 30 seconds and then extension at 72°C for 1 minute with the three later steps going through 30 cycles. The final extension took place at 72°C for 9 minutes. Entopolypoides macaci PCR conditions were 94°C for 5 minutes’ initial denaturation followed by denaturation at 94°C for 30 seconds, primer annealing 62°C for 30seconds and extension at 72°C for 1 minute with the three later steps going through 34 cycles then final extension at 72°C for 5 minutes (Appendix 3). The resulting PCR products were visualized on a 1.5% agarose gel stained with ethidium bromide then visualized under ultraviolet light
Results
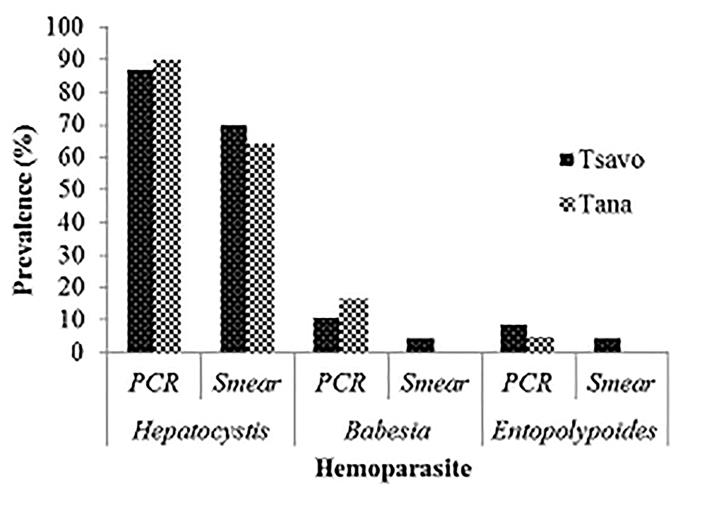
Prevalence of Zoonotic Hemoparasites in Free Ranging Baboons at Human Wildlife Interface: Two methods (microscopy and PCR) were utilized to screen for hemoparasite presence in two baboon populations and our results showed PCR analysis was a more sensitive diagnostic method compared to microscopy. Blood smear microscopy results indicated that TWPNP baboons had low prevalence of both B. microti (4.3%) and E. macaci (4.3%) whereas TRPR baboons tested negative for both parasites. However, results from PCR screening showed a slight increase in the prevalence of both parasites; TWPN baboons had a prevalence of 10.9% (5/46) for B. microti and 8.7% (4/46) for E. macaci while TRPR baboons had 16.7% (10/60) for B. microti and 5% (3/60) for E .macaci (Tables 1 & 2, Figures 1-3). We also found high prevalence of H. kochi at both sites by both microscopy and PCR screening; TRPR (70%) and TWNP (63%) results by microscopy and TRPR (87%) and TWNP (90%) by PCR screening.
| PCR | Blood smear | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | Prev (%) ±SE | N | Prev (%) ±SE | ||||||||||
| TRPR H. kochi | 60 | 90±3.9 | TRPR H. kochi | 60 | 70±6 | ||||||||
| TRPR B. microti | 60 | 17±4.9 | TWNP H. kochi | 46 | 63±7.2 | ||||||||
| TRPR E macaci | 60 | 5±2.8 | TWNP Babesia | 46 | 4±3 | ||||||||
| TWNP H. kochi | 46 | 87±5 | |||||||||||
| TWNP B. microti | 46 | 11±4.6 | |||||||||||
| TWNP E. macaci | 46 | 9±4.2 |
Table 1: Prevalence (Prev) of hemoparasites (%±SE) based on blood smear and PCR diagnosis.
| N | Mean±SEM | |||||
|---|---|---|---|---|---|---|
| H. kochi TWNP | 46 | 0.116±0.04* | ||||
| H kochi TRPR | 60 | 0.143±0.04* | ||||
| Babesia TWNP | 46 | 0.05±0.023* | ||||
| Entopolypoides TWNP | 46 | 0.02±0.015 |
Table 2: Mean parasitemia %±SEM for reported hemoparasites from blood smears. NB: *denotes that the parasitemia was statistically
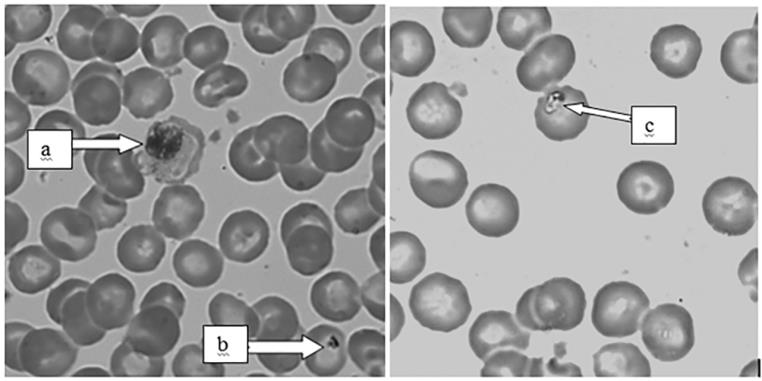
Presence of Potential Vectors that Transmit Zoonotic Hemoparasites
In both sites, the baboons were infested with hard (Ixodidae) ticks with TWNP baboons having a higher prevalence (28%) of ticks than TRPR (23.3%). The species of ticks found in the baboons were Rhipicepalus pulchellus, Rhipicephalus simus simus and Hyalomma truncatum (Figure 4). The ticks were found in the armpit, inguinal and perineal and ventral abdominal regions of the baboons. A substantial proportion of baboons had All Tana River baboons had fever (100%); a fever of between 38.8°C and 41.6°C and with an average of 40°C while in Tsavo, 98% had fever of between 38.6°C and 41.8°C with an average of 40.3°C.
erythematous lesions from points where ticks had recently fallen off (55% in TWNP and 48.3% in TRPR).
Consequences of Hemoparasite Infection in Baboons

truncatum with prominent scutum and prominent capitulum.
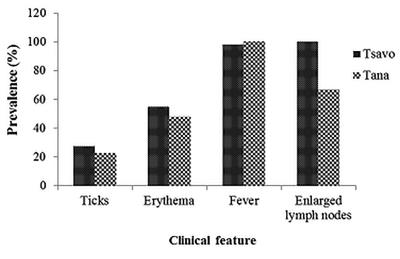
Tsavo represents TWNP and Tana represents TRPR. Erythema lesions appeared in areas where ticks are normally lodged, fever range was between 38.6°C-41.8°C, superficial lymph nodes were enlarged twice or over the normal size.
Discussion
The current study reported the occurrence of parasites at the wildlife-human interface in TRPR and TWNP in Kenya. The following hemoparasites were detected through microscopy and molecular screening; H. kochi, B. microti, and E. macaci, with the most prevalent being Hepatocystis kochi while the rest having very low prevalences. Hepatocystis kochi has been reported previously in baboons, African green monkeys and Sykes monkeys [8, 22]. The parasite though highly endemic has not been shown cause clinical disease in NHPs although in the current study. The zoonotic significance of the parasite is also not known.
Baboons in these two populations were also infested with B. microti which is a zoonotic hemoparasite. The prevalence (8.7%) of B. microti found in the current study was higher than reported by Akinyi, et al. [23] in baboons from Amboseli national park in Kenya and lower than reported by Mamuun, et al. [8] in baboons and Nakayima, et al. [24]. Maamun, et al. [8] also detected presence of B. microti in R. simus ticks, indicating that this tick is involved in transmission of the parasite. Babesiosis due to B. microti is an emerging zoonotic disease, which is a mainly asymptomatic infection, with malarial like signs and can lead to severe disease and death in Immuno compromised patients Various wildlife species, including NHPs are likely to be reservoir hosts of B. microti and the primary vectors of Babesia are Ixodid ticks, especially genus Ixodes [25]. The presence of Babesia infection in natural baboon populations presents more opportunities for challenging areas of research or chemotherapy where human babesiosis is concerned.
A third haemoparasite that was also identified in this study was Entopolypoides macaci, which is closely related B. microti. E. macaci has been reported before in NHPs and the prevalence reported in this study is lower than previously reported in other baboon populations [12, 8]. In baboons, the parasites can acute hemolytic crises following immunosuppression and affect results of baboon as research model [12]. Molecular identification of this parasite was not done and future studies should endeavor to characterize this parasite.
The current study highlights the presence of the zoonotic hemoparasites in baboons in areas of high NHP- human interactions. Between 22% and 28% of the sampled baboons had high tick infestation and these ticks can bite humans hence leading to zoonotic disease transmission. Humans are likely to increase the risk of contact with arthropod vectors such as ticks when walking in areas with a lot of vegetation such as the animal parks. For instance, individuals living around the two sites we studied have been reported to access the parks for various reasons including hunting for bush meat [18]. Consistent with previous studies in Kenya, the tick species identified in baboons included Rhipicephalus simus, Rhipicephalus pulchellus and Hyalomma truncatum [23, 8]. Further studies should be carried out in the study areas to determine the prevalence of human babesiosis and Entopolypoides infection.
Apart from disease transmission, other detrimental effects of ticks on baboons include reduction in packed cell volume and as seen in the current study erythema and skin wounds [23]. Fever was also evident in these baboons and the most likely cause of these would be the hemoparasites [26]. The hemoparasites were also likely to be the cause of lymph node enlargement evident in most of the baboon being systemic infections. However, because wild animals are likely to be infected by many other parasites, it is very difficult to claim causality [27, 28, 29].
Conclusion
The current study has highlighted presence of vector transmitted Apicomplexan parasites in baboons. Apart from H. kochi, the rest of the hemoparasites have zoonotic potential [30]. These hemoparasites clinically present as malaria and they also have intraerythrocytic stages hence chances of misdiagnosis when blood smears are used. There is needed to relook at the diagnostic methods that are capable of identifying cases of zoonotic infections such as Babesia and Entopolypoides in health facilities. This is because with poor diagnosis, there are chances of abuse of malarial drugs in treating these infections which accelerates development of drug resistance. In addition, with the misdiagnosis, the pathogens would not be cleared in the population and transmission would continue.
Acknowledgements
We also acknowledge assistance by Mr. James Ndungu of the Animal science department, Institute of Primate Research who assisted with animal trapping and sample collection. We also thank Mr. Mburu for his assistance during field work. We are grateful to Mr. Kenneth Waititu who helped in laboratory analysis. Finally, we acknowledge the Institute of Primate Research administration for facilitating this project.
Funding
The study was supported by funding from the National Council of Science, Technology and Innovation, Ministry of Science and Technology of Kenya (Grant Ref. Number: NCST/5/003/w/2nd CALL/55).
Conflict of Interest
I am one of the editors of the journal
References
-
Gortazar C, Ferroglio E, Höfle U, Frölich K, Vicente J (2007) Diseases shared between wildlife and livestock: A European perspective. Eur J Wildl Res 53: 241-256.
-
Acha PN, Szyfres B (2003) Zoonoses and Communicable Diseases Common to Man and Animals. 3rd (Edn.), PAHO, Washington.
-
Meslin F (1995) Zoonoses: a worldwide challenge (perspectives on future roles of veterinarians in public health). 8th International Conference of Institutions of Tropical Veterinary Medicine, Berlin.
-
Bastone P, Truyen U, Löchelt M (2003) Potential of Zoonotic Transmission of Non-Primate Foamy Viruses to Humans. J Vet Med B 50(9): 417-423.
-
Legesse M, Erko B (2004) Zoonotic intestinal parasites in Papio anubis (baboon) and Cercopithecus aethiops (vervet) from four localities in Ethiopia. Acta Trop 90(3): 231-236.
-
Voller A (1972) Plasmodium and Hepatocystis. In: Fiennes RNTW (Eds.), Pathology of simian primates, Part II: Infectious and Parasitic Diseases. New York.
-
Esch KJ, Peterson CA (2013) Transmission and epidemiology of zoonotic protozoal diseases of companion animals. Clin Rev 26(1): 58-85.
-
Maamun JM, Suleiman MA, Akinyi M, Ozwara H, Kariuki T, et al, (2011) Prevalence of Babesia microti in free-ranging baboons and African green monkeys. J Parasitol 97(1): 63-67.
-
Wallis J, Lee RD (1999) Primate conservation: The prevention of disease transmission. Int J Primatol 2(6): 803-826.
-
Mbaya AW, Aliyu MM, Nwosu CO, Ibrahim UI (2008) Captive wild animals as potential reservoirs of hemo and ectoparasitic infections of man and animals in the arid region of northeastern Nigeria. Veterinarski Arhiv 78(5): 429-440.
-
Kjemtrup AM, Conrad PA (2000) Human babesiosis: an emerging tick-borne disease. Int J Parasitol 30(12- 13): 1323-1337.
-
Bronsdon MA, Homer MJ, Magera JM, Harrison C, Andrews RG, et al. (1999) Detection of enzootic babesiosis in baboons (Papio cynocephalus) and phylogenetic evidence supporting synonymy of the genera Entopolypoides and Babesia. J Clin Microbiol 37(5): 1548-1553.
-
Emerson CL, Tsai CC, Holland CJ, Ralston P, Diluzio ME (1990) Recrudescence of Entopolypoides macaci Mayer, (1933) (Babesiidae) infection secondary to stress in long-tailed macaques (Macaca fascicularis). Lab Anim Sci 40(2): 169-171.
-
Vooberg-vd Wel A, Kocken CH, Zeeman AM, Thomas AW (2008) Detection of new _Babesia microti-_like parasites in a suppressed Plasmodium cynomolgi infection. Am J Trop Med Hyg 78 (4): 643-645.
-
Mwale S (2000) Changing Relationships: The History and Future of Wildlife Conservation in Kenya. Swara 22: 11-17.
-
Ottichilo WK, de Leeuw J, Skidmore AK, Prins HHT, Said MY (2000) Population trends of large non- migratory wild herbivores and livestock in the Masai Mara ecosystem, Kenya, between 1977 and 1997. Afr J Ecol 38(3): 202-216.
-
Butynski TM, Mwangi G (1994) Conservation status and distribution of the Tana River Red Colobus and Crested Mangabey. Report to Zoo Atlanta, the Kenya Wildlife Service pp: 1-58.
-
Mwangi KD, Akinyi M, Maloba F, Ngotho M, Kagira J, et al. (2016) Socioeconomic and Health Implications of Human-Wildlife Interactions in Nthongoni, Eastern Kenya. Afr J Wildl Res 46(2): 87-102.
-
Sheldrick D (2002) Annual De-Snaring Report by David Sheldrick De-snaring teams. David Sheldrick Wildlife Trust, Kenya.
-
Karere GM, Oguge NO, Kirathe J, Muoria PK, Moinde NN, et al. (2004) Population size and distribution of primates in the lower Tana River, Kenya. Int J Parasitol 25(2): 351-365.
-
Moinde NN, Higashi H, Suleiman MA, Hau J (2004) Habituation, capture and relocation of endangered arboreal non-human primates: Experience from relocation of Sykes monkeys (Cercopithecus mitis albotorquatus) on the coast of Kenya. Anim Welfare 13: 343-353.
-
Moore JA, Kuntz RE (1975) Entopolypoides macaci Mayer (1934) in African baboon (Papio cynocephalus 1776). J Med Primatol 4: 1-7.
-
Akinyi MY, Tung J, Jeneby M, Patel NB, Altmann J, et al. (2013) Role of grooming in reducing tick load in wild baboons (Papio cynocephalus). Anim Behav 85(3): 559-568.
-
Nakayima J, Hayashida K, Nakao R, Ishii A, Ogawa H, et al. (2014) Detection and characterization of zoonotic pathogens of free-ranging non-human primates from Zambia. Parasit Vectors 7: 490.
-
Yabsley MJ, Shock BC (2013) Natural history of Zoonotic Babesia: role of wildlife reservoirs. Int J Parasitol 2: 18-31.
-
Demessie Y, Derso S (2015) Tick Borne Hemoparasitic Diseases of Ruminants: A Review. Adv Biol Res 9(4): 210-224.
-
Homer MJ, Aguilar Delfin I, Telford SR, Krause PJ, Persing DH (2000) Babesiosis. Clin Microbiol Rev 13(3): 451-469.
-
Jeneby MM, Ngeiywa, Yole DS, Mwenda JM, Suleman MA, et al. (2008) Enzootic simian piroplasm (Entopolypoides macaci) in wild-caught Kenyan non- human primates. J Med Primatol 37(6): 329-336.
-
Tung J, Primus A, Bouley AJ, Severson TF, Alberts SC, et al. (2009) Evolution of a malaria resistance gene in wild primates. Nature 460(7253): 388-392.
-
Wolfe ND, Daszak P, Kilpatrick AM, Burke DS (2005) Bush meat hunting, deforestation, and prediction of zoonotic disease. Emerg Infect Dis 11(12): 1822- 1827.
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System