Description of Pseudobenedenia coriicepsi Sp. N. (Monogenea: Capsalidae) from the Antarctic Black Rockcod, Notothenia coriiceps Richardson in Coastal Waters of West Antarctica Using Novel SEM Images, Energy Dispersive X-Ray Analysis and Molecular Analysis
Morphological and metrical analyses of sclerotized parts of haptor and body morphology and anatomy of monogeneans from Antarctic rockcod Notothenia coriiceps Richardson revealed the presence of a new species, Pseudobenedenia coriicepsi sp. n., supported with molecular studies, EDXA and scanning electron microscopy. New species differs from Pseudobenedenia nototheniae Johnston, 1931 by characteristic shape of anterior hamulus, size of sclerotized structures of haptor, measurements of the body, measurements ratios, v-shaped consistent commissure in vitellaria field, relative proportions of body and haptor, unlobed testes, and by host species. Published drawings of sclerotized parts of haptor and measurements of the species of the genus Pseudobenedenia Johnston, 1931 are provided for comparison and discussed. Status of P. nototheniae as species complex and host-specificity in this group are discussed. Molecular data of the P. coriicepsi sp. n. (partial 18S and 28S) were generated to provide the first molecular analysis for any species of Pseudobenedenia to date. Phylogenetic analyses inferred from the two molecular markers indicate that P. coriicepsi sp. n. is closely related to the species of the genus Benedenia and formed a sister clade to it as positioned under the family Capsalidae.
Introduction
An Antarctic monogenean Pseudobenedenia nototheniae Johnston 1931 (Monogenea: Capsalidae) has an obscure identity. Originally, Johnston [1] mentioned that “specimens (of its host) were obtained from Notothenia angustata (currently valid – this paper) and Notothenia colbecki (now considered to be N. microlepidota Hutton, 1875, according to DeVitt, 1970 – this paper)” “gliding over the body” of the host, from “subantarctic islands of New Zealand, as identified by prof. W. B. Benham”. Later host identification was made by Waite [2] who considered N. angustata to be a synonym of N. macrocephala (currently another valid species, Paranotothenia magellanica). Then, Johnston while giving an updated full description of P. nototheniae studied the material collected from the head and the body of N. macrocephala at Macquarie Island in 1912 [3]. Waite [2] who worked with the collection of parasites, mentioned, “They (fish) were all of the same species, somewhat resembling rock cod”. While dealing with collections of Monogeneans, Johnston [3] mentioned a huge size variation of collected worms, that he considered as a new species P. nototheniae, the largest worm measured 7 mm by 4 mm, the smallest – 4.7 mm by 2.3 mm.
Remarkably, to the best of our knowledge, there were no published records on P. nototheniae from its type hosts Nothothenia microlepidota Hutton, 1875 and Paranototenia magellanica, except a few other publications [4, 5, 6, 7] that appeared after the original description, as well as the records of these hosts themselves were extremely rare_. All records on so-called _P. nothotheniae were from other species of fish [8, 9, 10, 11, 12, 13]. Curiously, P. nototheniae described by Johnston [1, 3] from N. microlepidota (old N. colbecki), was only mentioned from this host in a few publications of same author with co- authors [6, 7], but never registered as a new finding from this host by other researchers [12, 13]. Also, surprisingly no images are available for this fish host species in the FishBase and Elchmeyer Catalog of fishes [14, 15], Klapper et al. [13] though it is considered a valid species according to Dr. Agnes Dettai (personal communication) and Froese R, et al. [16]. Accordingly, we can guess that this fish species could go extinct, or was misidentified during the process of the worm description, or is extremely rare to be examined by parasitologists [17].
Another curious fact regarding the other fish host mentioned in the original description of P. nototheniae by Johnston is that the fish species Paranotothenia magellanica (Forster, 1801) was mentioned as Notothenia macrocephala or “similar to N. macrocephala”. Except few checklist publications not based on the freshly collected material but just on literature reviews [12, 18, 19], we did not find any published data on new collections of P. nototheniae from this particular fish species, except mentioning the host species [17] in Chile coastal waters.
Examining the fresh material of Pseudobenedenia collected from another teleost fish host, Notothenia coriiceps Richardson, 1844 in another location (Galindez Island, Argentine Islands, West Antarctica), at first glance, we noticed that all mature specimens’ average size range was never 7 mm by 4 mm, but closer to the smallest, e.g., 4.97 mm (3.75–6.00 mm). Despite the specimens were initially identified as P. nototheniae [4, 5], we scrupulously compared the morphology of our worms to Johnston’s descriptions [1, 3] and found other significant morphological differences from the original description of P. nototheniae, which led to the writing of this manuscript.
Dollfus and Euzet [8] also found some morphological features of P. nototheniae that disagreed with the description of Johnston [1, 3] and provided a redescription of P. nototheniae based on fresh material collected from N. rossii and from N. colbecki stored in the Helminthological Collection of the Zoological Department of Adelaide University [8]. Later, Szidat [20] published a very brief description of the second species of the genus Pseudobenedenia, P. lauriei Szidat, 1965 collected from Nothothenia neglecta near Scotia Bay. In 1968, Hargis and Dillon [21] discovered one more species, Pseudobenedenia shorti Hargis et Dillon, 1968, that was described from several fish hosts, three of them belonging to the family Nototheniidae Günther, 1861, namely Pseudotrematomus bernacchii (Boulenger 1902), Trematomus hansoni Boulenger, 1902, Pseudotrematomus centronotus (Regan 1914), and one to the family Zoarcidae Swainson, 1839 – Lycodichthys dearborni (DeWitt 1962), though the last host was discuss later by these authors as possibly erroneous. That description of P. shorti missed the information on the posterior anchors, and was based on a mixture of specimens of different ages. Later Timofeeva, et al. [11] described two new Pseudobenedenia species from the Atlantic region of Antarctica and Subantarctica – Pseudobenedenia dissostichii Timofeeva, Gaevskaja et Kovaliova, 1987 from a body surface of Macrias amissus Gill et Townsend, 1901, and Pseudobenedenia gibberifrons Timofeeva, Gaevskaja et Kovaliova, 1987 from pectoral fins of Gobionotothen gibberifrons (Lönnberg 1905). Yamaguti [22] described three species of Pseudobenedenia from Hawaiian fishes – Pseudobenedenia elongate Yamaguti, 1968 from gills of Coris gaimard (Quoy et Gaimard 1824), Coris flavovittata (Bennett 1828) and Coris sp., Pseudobenedenia merithe Yamaguti, 1968 from gills of Merithe macrocephala, the host that we did not find in modern fish data bases, and Pseudobenedenia ovalis Yamaguti, 1968 from the body surface of Etelis carbunculus Cuvier, 1828 and Priacanthus hamrur (Fabricius, 1775). However, Yamaguti’s descriptions contained no information on haptor structures, except very schematic drawings.
At the same time, publications of multiple records on Antarctic capsalids identified as P. nototheniae from different nototheniid fish hosts widened the list of hosts of this monogenean up to 11 teleost fish species such as Dissostichus eleginoides Smitt, 1898, D. mawsoni Norman, 1937, Lepidonotothen squamifrons (Günther, 1880), Trematomus bernacchii Boulenger, 1902, Trematomus sp. and some others [10, 12, 13]. In 1976, Gibson [10] synonymized P. lauriei with P. nototheniae without studying fresh material; his synonymy was based only on a brief description made by Szidat [20]. Since then, in all publications, the capsalids collected from N. coriiceps, as well as from a wide variety of other nototheniid fishes were identified as P. nototheniae, providing a wide range of metric characteristics and detailed morphological studies, and a question of host specificity was rarely brought up [10, 12, 13, 23, 24].
In Monogenea, morphology of hard parts of the haptor and the genital system (if there are any sclerotized structures) as well as specificity to fish hosts are considered as the key features in species discrimination [25, 26, 27, 28, 29]. In addition, the site of infection and host specificity are considered among the most important diagnostic features in Monogenea [25, 26, 27]. However, the original description of type-host species for P. nototheniae, as well as for P. lauriei, P. shorti, and all species described by Yamaguti [22] missed this crucial information that left a possibility of misidentification of these Pseudobenedenia species from various fish hosts wide open. We here are filling in for species of Pseudobenedenia available for us now and analysis of literature data. We suppose that Pseudobenedenia species being Monogeneans that live on a skin, are theoretically susceptible to various immune factors present in the mucus of the fish host that conflicts with a very wide range of hosts. A wide range of variations in some morphometric characteristics of Pseudobenedenia spp. that sometimes vary 3-fold is also of our concern. All this prompted us to perform an analysis of published literature data as well as to re-examine of newly collected material for clarifying the taxonomical status of available species of Pseudobenedenia using modern research techniques and approaches. Our study aimed to revise all available published descriptions of Pseudobenedenia species collected from N. coriiceps and provide strong support of presence of a new species of Pseudobenedeniae, P. coriiceps sp. n. on the base of morphological examination of fresh material using morphological and molecular data. For this the light microscopy and scanning electron microscopy (SEM) were used to give descriptions of sclerotized structures of the haptor and other taxonomically important characteristics of Pseudobenedenia. We also characterized P. coriiceps sp. n. molecularly (18S and 28S rRNA gene) to explore the phylogenetic relationships within the family Capsalidae Baird, 1853.
Material and Methods
Field studies and material collection were carried out in April 2014 – January 2015 near the Ukrainian Antarctic Station “Akademik Vernadsky”(Galindez Island, Argentine Islands, West Antarctica; 65˚15′S, 64˚16′W). A total of 106 individuals of Antarctic black rock cod N. coriiceps were caught in coastal water off the shore of Galindez Island at depths from 10 to 30 m using a fishing rod [4]. All fish collected were immediately transported to the laboratory, measured, and examined using the standard parasitological techniques [30]. Helminths were collected manually from the skin, washed in saline (0.9 % NaCl), and fixed in 70 % ethanol.
Methods of Microscopy
Preliminary laboratory examination including identification of the parasites to the genus level was performed in the Department of Parasitology of I. I. Schmalhausen Institute of Zoology NAS of Ukraine in Kyiv, using a Zeiss Axio Imager M1 compound microscope equipped with DIC optics and a digital imaging system (Carl Zeiss AG Light Microscopy, Göttingen, Germany). Further detailed studies of inner morphology of the parasite, whole mount preparations, light microscopic imaging, drawings and data analysis of measurements were held at the Parasitology Center (PCI), Scottsdale, Arizona, USA. Measurements are in millimeters (mm) unless otherwise noted; the range is followed by the mean values between parentheses. Width measurements represent maximum width. Microscope images were created using 10× and 40× objective lenses of a BH2 light Olympus microscope (Olympus Optical Co., Osachishibamiya, Okaya, Nagano, Japan) attached to an AmScope 1000 video camera (United Scope LLC, DBA AmScope, Irvine, California), linked to an ASUS laptop equipped with HDMI high-definition multimedia interface system (Taiwan-USA, Fremont, California). To study the inner anatomy, monogeneans were kept in glycerin for a few days and later fixed in glycerin jelly without strong pressure. Because the worms are very thick, after holding them in glycerin, part of the samples was fixed in glycerin jelly and then slowly warmed up the slide from below with gentle pressure on the coverslip. That made the worm body 3-4 times thinner and helped to get a better view of sclerotized structures of the haptor. Correspondingly, these flattened worms became unnaturally bigger and their soft structures and body parameters markedly differ a lot from the non-flattened ones; thus, we did not use artificially flattened specimens for measurements, only for studies of sclerotized structures [26, 27]. Several of monogeneans were stained in Mayer’s acid carmine, destained in 4% hydrochloric acid in 70% ethanol, dehydrated in ascending concentrations of ethanol (24 hours each), and cleared in 100% xylene then in 50% Canada balsam and 50% xylene (24 hours each). Whole worms were then mounted in Canada balsam. While studying the shape of sclerotized parts of haptor, we additionally used a technique of ratios used for Polystomatidae by Euzet, et al. [31, 32, 33, 34]. This feature was also used by us to create a diagnostic key for species of Polystoma of Ukraine [35]. Measurements are in millimeters (mm) unless otherwise noted; the range is followed by the mean values in parentheses. Width measurements represent maximum width.
Line Drawings
Line drawings were created by using a Ken-A-Vision micro projector (Ward’s Biological Supply Co., Rochester, N.Y.) which uses cool quartz iodine 150W illumination. Images of stained whole mounted specimens are projected vertically on 300 series Bristol draft paper (Starthmore, Westfield, Massachusetts), then traced and inked with India ink.
Specimens
One holotype and one paratype of Pseudobenedenia coriicepsi sp. n. were accessioned (P-2023-005) and cataloged (HWML-216939) at the University of Nebraska’s State Museum Harold W. Manter Laboratory (HWML) collection in Lincoln, Nebraska, USA.
Scanning Electron Microscopy (SEM)
Three specimens fixed and stored in 70% ethanol were processed for scanning electron microscopy (SEM) following standard methods [36]. This included critical point drying (CPD) (Tousimis Automandri 931.GL) and mounting on aluminum SEM sample mounts (stubs) using conductive double-sided carbon tape. Samples were sputter coated with an 80%–20% gold-palladium target for 3 minutes using a sputter coater (Quorum (Q150T ES) www.quorumtech.com) equipped with a planetary stage, depositing an approximate thickness of 20 nm. Samples were placed and observed in a FEI Helios Dual Beam Nanolab 600 (FEI, Hillsboro, Oregon) Scanning Electron Microscope (FEI, Hillsboro, Oregon). Samples were imaged using an accelerating voltage of 5 kV, and a probe current of 86 pA, at a high vacuuming SE detector.
Energy Dispersive X-Ray Analysis (EDXA)
The Helios Nanolab 600 was equipped with an Energy Dispersive X-Ray analysis (EDXA) TEAM Pegasus system (Mahwah, NJ) with an Octane Plus detector. The sectioned cuts were analyzed by EDXA. Spectra of selected areas were collected from the center and the edge of each cross-section. EDXA spectra were collected using an accelerating voltage of 15 kV, and a probe current of 1.4 nA. Data collected included images of the displayed spectra as well as the raw collected data. Relative elemental percentages were generated by the TEAM software.
Molecular Methods
DNA Extraction, PCR Amplification and Sequencing
Genomic DNA was extracted from four different isolates preserved in 90% molecular grade ethanol using Qiagen DNeasy tissue kit (Qiagen Inc., Valencia, California, USA) according to the manufacturer’s instructions. Polymerase chain reaction (PCR) amplifications were performed in 25 μl reactions containing 4 μl of extracted DNA, 5 µl 1 mM deoxyribonucleotide triphosphates (dNTPs, Biotools, Madrid, Spain), 0.90 µl of each primer, 2.5 µl of 10x Taq buffer (Biotools) with MgCl2, 0.60 µl of Taq polymerase (1U; Biotools) and 11.10 µl of distilled water. Partial 18S and 28S rDNA fragments were amplified using primers as mentioned in the Table 1. The following thermocycling profile was utilized: denaturation of DNA at 94°C for 3 min; 35 cycles of amplification at 94°C for 45 s, 55°C for 40 s and 72°C for 2 min with an final 10 min extension hold at 72°C. PCR products were purified prior to sequencing using PurelinkTM Quick Gel Extraction and PCR Purification Combo Kit (Invitrogen, Löhne, Germany). Amplicons were cycle-sequenced from both strands with the PCR primers mentioned in the table 1 using ABI BigDye™ chemistry (Applied Biosystems, Foster City, California) according to the manufacturer’s protocol in automated sequencer. Contiguous sequences were assembled and edited using MEGA v11 [37] and submitted to GenBank.
| Primer | Sequence (5’–3’) | Source |
|---|---|---|
| 18S rDNA | ||
| WormA | GCGAATGGCTCATTAAATCAG | Olson and Littlewood, 2002 [54] |
| 1270R | CCGTCAATTCCTTTAAGT | Olson and Littlewood, 2002 [54] |
| 930F | GCATGGAATAATGGAATAGG | Olson and Littlewood, 2002 [54] |
| WormB | CTTGTTACGACTTTTACTTCC | Olson and Littlewood, 2002 [54] |
| 28S rDNA | ||
| Ancy55F | GAGATTAGCCCATCACCGAAG | Plaisance et al. 2005 [55] |
| Ancy1200R | CACCATCTTTCGGGTCTCAACC | Plaisance et al. 2005 [55] |
| L300F | CAAGTACCGTGAGGGAAAGTTG | Littlewood et al., 2000 [56] |
| ECD2 | CCTTGGTCCGTGTTTCAAGACGGG | Littlewood et al., 2000 [56] |
Table 1: Primers used for PCR amplification and sequencing.
Phylogenetic Analyses
Newly generated contiguous sequences of the 18S and 28S dataset were assembled using MEGA v.11 and subjected to a BLASTn search on the NCBI GenBank database. 18S and 28S sequences were separately aligned using default parameters in MUSCLE implemented in GUIDENCE 2 [37, 38]. Molecular identification and evaluation of phylogenetic relationships were accomplished using 2 approaches, maximum likelihood (ML) and Bayesian inference (BI) implemented in MEGA v. 11 and TOPALi v. 2.5 [39], respectively. Proceeding to the analysis, jModelTest 2.1.4 [40] was used to estimate the best- fitting model of nucleotide substitution for both datasets. This was the invariant sites and gamma-distributed among site variation (GTR + I + G) model was found the best nucleotide substitution models for 18S and 28S rDNA data sets. Maximum Likelihood (ML) tree was generated with 1,000 repetitions of bootstrap support values. Bayesian Inference (BI) tree was generated in TOPALi v. 2.5 by running two independent MCMC runs of four chains for 107 generations, sampling tree topologies every 1,000 generations. Pairwise genetic distances (uncorrected p-distance model) for each region 18S and 28S were calculated in MEGA v.11. Species of the genus Polylabris [41] were used as outgroup for the 18S and 28S rDNA gene analysis.
Results
Examination of the fresh material collected from N. coriiceps revealed morphological differences between our specimens and those of P. nototheniae described by Johnston [1, 3]. After concluding different studies listed herein, we consider our samples as a new species Pseudobenedenia coriicepsi sp.n. as described following.
Description Of Pseudobenedenia coriicepsi (Figures: 1-19 & Table 2)
Type locality: coastal waters near Galindez Island (Argentine Islands, West Antarctica) (65˚15′ S, 64˚16′ W). Type host: Notothenia coriiceps Richardson. Site of infection: body surface. Material studied for description: 3 specimens stained with acetic carmine, 8 with glycerin jelly slides, and 4 for SEM and DNA analysis.
Deposited specimens: holotype and paratype (accession numbers P-2023-005), catalogued (HWML-216939), University of Nebraska, State Museum, Lincoln Nebraska, USA. ZooBank LSID#F81768B5-3C8E-45F1-9DEC-4574F3D51386
| Host | Paranotothenia magellanica, N. microlepidota, N. angustata (?) | N. rossi (rossi) | N. rossi (marmorata) | Notothenia neglecta | Notothenia coriiceps | Gobionotothen gibberifrons | Macrias amissus | Notothenia coriiceps |
|---|---|---|---|---|---|---|---|---|
| Parasite | Pseudo benedenia nototheniae | Pseudo benedenia nototheniae* | Pseudo benedenia nototheniae** | Pseudo benedenia lauriei | Pseudo benedenia nototheniae*** | Pseudo benedenia gibberifrons | Pseudo benedenia dissostichii | Pseudo benedenia coriicepsi sp. n. |
| Authority | Johnston, 1931 Johnston, 1937 | Dolfus & Euset 1964 | Timofeeva et al, 1987 | Szidat, 1965 | Willams et al., 1973 | Timofeeva et al, 1987 | Timofeeva et al, 1987 | Our data |
| Locality | Sub-Antarctic islands of New Zealand, Macquarie Isl. | Kergelen | Georgiy Isl., Mordvinov Isl. | Laurie Isl. | S Ornikean Isl. | Georgiy Isl., Mordvinov Isl. | Georgiy Isl., Mordvinov Isl. | Galindez Isl. |
| Site of infection | Body surface | Body surface | Body surface, oral cavity | Body surface | Body surface | Pectoral fins | Body surface | body surface |
| Number of radiating muscular septa | 6 ? | 7 | n/a | n/a | n/a | n/a | n/a | 6 |
| Body length | 4.7–7* | 4.8–6.8 | 5.9 (5.2–6.7) | 4.5 | 3.87 (3.1– 4.7)**** | 5.6 (4.7-6.2) | 8.7 (7.1-10.8) | 6.03 (4.75– 7.00) |
| Body width | 2.3–4 | 3–4.5 | 2.9 (2.6–3.8) | 2.5 | 2.46 (1.9–3.0) | 2.6 (2.1-3.7) | 3.9 (3.3-4.8) | 2.73 (2.00– 3.70) |
| Oral suckers diameter | 0.8 | 0.7–0.8 | 0.64 (0.51– 0.73) | 0.4 | 0.5 (0.38– 0.62) × 0.41 (0.35–0.48) 0.5 (0.4–0.59) × 0.41 (0.33– 0.52) | 0.76 (0.67- 0.88) | 1.04 (0.81- 1.24) | 0.45 (0.33– 0.52) × 0.48 (0.44–0.52) |
| Pharynx | 0.5 × 0.6 | 0.5 × 0.7 | 0.58 (0.42– 0.67) × 0.62 (0.44–0.69) | 0.3 | 0.37 (0.3–0.4) × 0.4 (0.28– 0.56) | 0.59(0.46-0.62) × 0.70(0.48- 0.80) | 0.88 (0.67- 1.12) ×1.07 (0.75-1.28) | 0.46 (0.33– 0.6) × 0.48 (0.35–0.7) |
| Ovary | 0.3 × 0.5–0.7 | 0.6 | 0.44 (0.38– 0.54) × 0.63 (0.44–0.71) | n/a | n/a | 0.48(0.37-0.55) ×0.67(0.43-.75) | 0.76 (0.67- 0.80) ×1.05 (0.91-1.15) | 0.47 (0.38– 0.55) × 0.50 (0.44–0.6) |
| Testes | 1.2–1.6 × 0.8–1.0 | 1.0 × 0.4 | 0.82 (0.64– 0.92) × 0.62 (0.49–0.77) | n/a | 0.61 (0.5– 0.76) × 0.39 (0.32–0.44) 0.62 (0.48– 0.76) × 0.41 (0.03–0.05) | 0.68(0.58-0.75) ×0.61(0.44- 0.75) | 1.62 (1.38- 1.95) × 0.91 (0.86-1.38) | 0.91 (0.73– 1.04) × 0.60 (0.49–0.78) |
| Bursa | 0.8 × 0.3 | n/a | 0.7 (0.62– 0.84) × 0.25 (0.24–0.27) | n/a | 0.31 (0.23– 0.42) × 0.15 (0.12–0.17) | 0.53(0.48-0.64) ×0.22(0.19- 0.24) | 1.16 (0.98- 1.39) × 0.34 (0.26-0.37) | 0.55 (0.50– 0.59) |
| Haptor diameter | 2.0–2.4 | 2 | 1.85 (1.48– 2.21) | n/a | 1.35 (1.06– 1.58) | 2.04(1.85-2.38) | 2.78 (2.46- 3.40) | 1.44 (1.14– 1.57) × 1.57 (1.40–1.75) |
| Anterior hamulus length | 0.38–0.53 | 0.5 | 0.51 (0.44– 0.54) | 0.4 | 0.4–0.42 | 0.58(0.55-0.64) | 0.80 (0.72- 0.86) | 0.51 (0.41– 0.60) |
| Posterior hamulus | 0.13–0.17 | 0.25 | 0.23 (0.18– 0.28) | 0.1 | 0.18–0.20 | 0.27(0.22-0.32) | 0.26 (0.21- 0.32) | 0.16 (0.13– 0.23) |
| Accessory sclerite | 0.23 | 0.37 | 0.34 (0.28– 0.37) | 0.28 | 0.32 (0.26– 0.38) | 0.39(0.33-0.45) | 0.53 (0.44- 0.59) | 0.38 (0.24– 0.46) |
| Egg | 0.2 × 0.1 | n/a | 0.22–0.13 | n/a | 0.32–0.09 | 0.24(0.22-0.27) ×0.14(0.10- 0.15) | 0.22 (0.19- 0.24) × 0.14 (0.10-0.16) | 0.31 (0.31– 0.31) × 0.13 (0.13–0.14) |
Table 2: Comparative morphometric data on _Pseudobenedenia s_pecies from Antarctic fish.
* Based on material from non-typical host N. rossi and original museum material Paranotothenia magellanica Non-typical host for P. nototheniae, no drawings provided *Non-typical host for P. nototheniae **Willams, et al. [9] measured the length of the body “from the anterior extremity to the peduncle”. We are not sure about other authors, but we assume that in all other cases, it is from the anterior end to the end of the haptor, as we provide it on our data * All measurements are in mm; n/a – not applicable Table 2:** Comparative morphometric data on _Pseudobenedenia s_pecies from Antarctic fish.
Gene Sequence
The 18S and 28S gene of ribosomal RNA sequences were deposited in GenBank under accession numbers: OQ803310, OQ803312, OR289962, OR289963 (for 18S); OQ820944, OQ820945, OR295462, OR295461 (for 28S), respectively.
Based on 8 mature specimens: body length 6.03 (4.75– 7.00), body width at the widest part 2.73 (2.00–3.70) (Figure 1 & Table 2). Head end with slightly cleft formed by muscle constrictions of anterior end of the body (Figure 2). Oral suckers 0.45 (0.33–0.52) × 0.48 (0.44–0.52) on both sides of mouth (Figure 2). Mouth well developed with muscular sucker-like opening, that has folds of irregular shape (Figures 1 & 10-11), diameter 0.48 (0.35–0.7). Pharynx 0.46 (0.33–0.6) × 0.48 (0.35–0.7), followed by oesophagus and two branches of intestine, extending to distal end of body. Intestine branches barely visible. Opening of bursa [0.55 (0.50–0.59)] with non-sclerotized cylindrical penis (Figures 1 & 12) on left site of body on ventral surface under center of oral sucker, with variable levels of inversion.
Penis surrounded by numerous sensory glands (Figure 12). Most ovary at right from midline, 0.47 (0.38–0.55) × 0.50 (0.44–0.6). Genital opening almost in midline of body, at level of left margin of oral opening. Two non-lobulated testes lie side by side, behind ovary, each measuring 0.91 (0.73–1.04) × 0.60 (0.49–0.78) with moderate perforations in mature specimens. Immature specimens’ testes without perforations. Single egg, if present, 0.31 (0.31–0.31) × 0.13 (0.13–0.14), with long coiled filament.
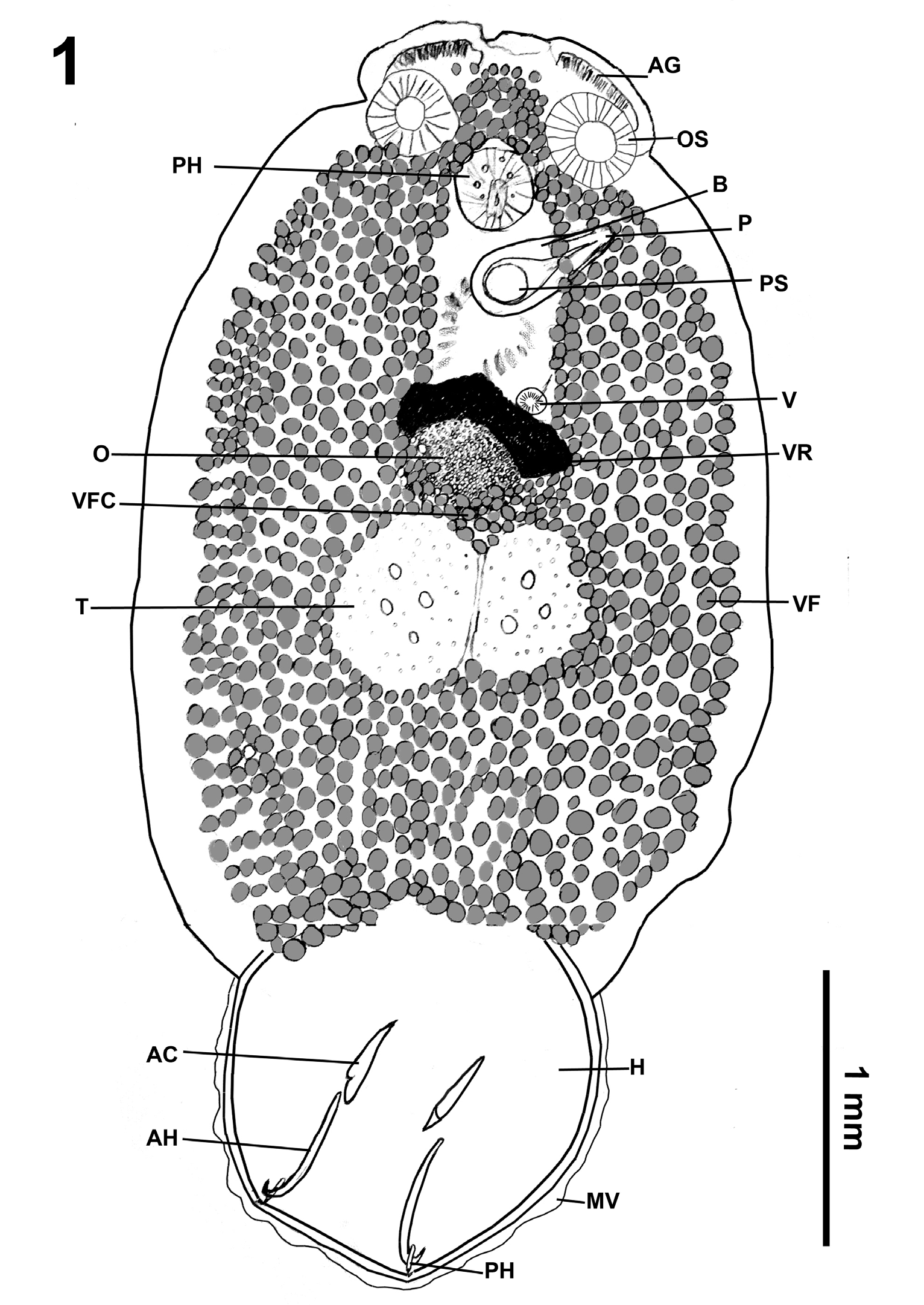
Figure 1: Total view of Pseudobenedenia coriicepsi sp. n. Abbreviations: AG – anterior glands, OS – oral sucker, PHA – pharynx, B – bursa, P – penis, PS –prostate sac, V – vagina, VR – vitelline reservoir, OV – ovary, VFC – vitellaria field commissure, T – testes, VF –vitellaria field, H – haptor, MV – marginal valve, AC – accessory sclerite, AH – anterior hamulus, PH – posterior hamulus.
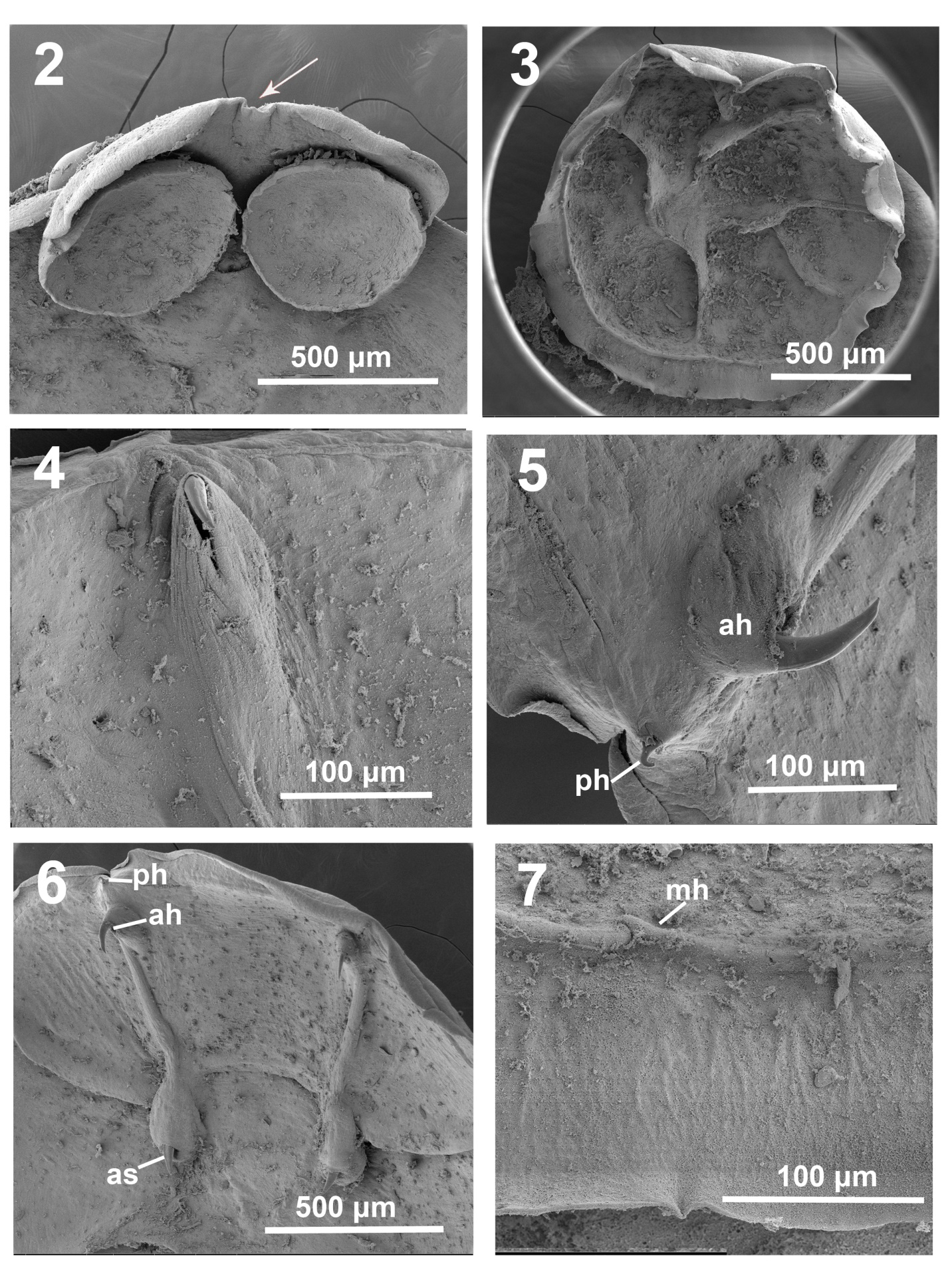
Figures 2-7: SEM images of Pseudobenedenia coriicepsi sp. n.: 2. Anterior end with oral suckers and small cleft 3. Haptor with 6 radiating septa. 4. Close view of anterior hamulus blade in a fibrous sac. 5. Duo of anterior and posterior hamuli in their natural position. 6. The whole view of all complex of main sclerotized structures, anterio and posterior hamuli and accessory sclerites. 7. Close view of a marginal hook.
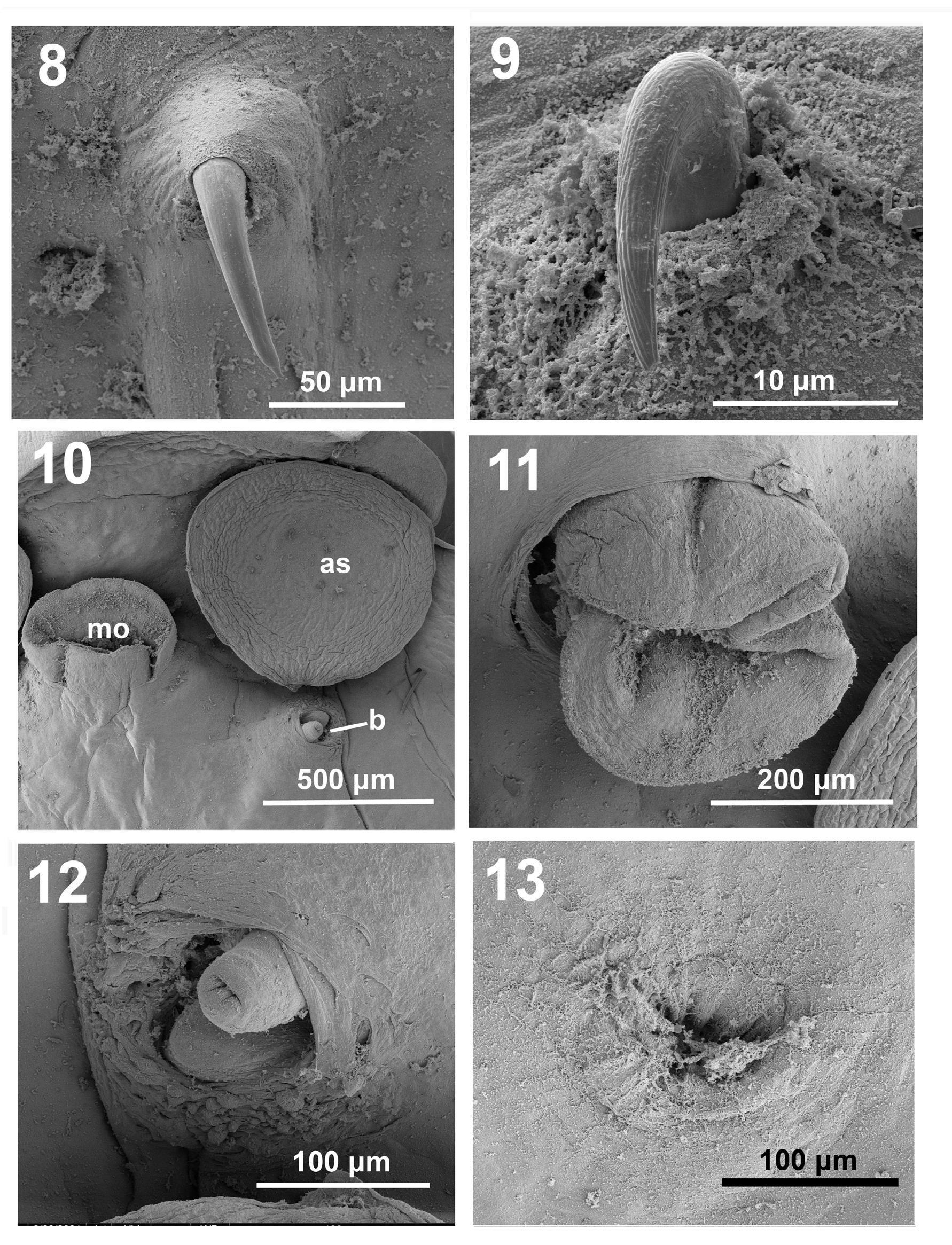
Figures 8-13: SEM images of Pseudobenedenia coriicepsi sp. n.: 8. Very close view of the blade of anterior hamulus. Note smooth surface. 9.Very close view of the blade of the posterior hamulus blade. Note ribbed surface. 10. Mouth opening (mo), left anterior sucker (as) and bursa opening under it. 11. Close view of mouth opening. 12. Close view of bursa opening with unsclerotised penis. 13. Close view of vaginal opening.
Body tapers posteriorly into short peduncle inserting into middle of antero-dorsal surface of haptor (Figure 1). Cuticle smooth, without fibers. Haptor 1.44 (1.14–1.57) × 1.57 (1.40–1.75) (Figures 1 & 3). Haptor (Figures 1& 3) comprises a disc-like muscular sucker divided by septa on ventral concave surface and armed with 3 pairs of sclerotized structures (anterior and posterior hamuli and accessory sclerites) (Figures 4-6 & 14B) and 14 pairs of marginal hooklets (Figure 7). Outer margin of haptor surrounded by thin marginal valve (Figure 3). Secretory gland cells condusive in adhesion on invaginated side of bowl-shaped haptor (Figure 4). Vitellaria field with characteristic v-shaped continuous commissure between ovary and testes (Figure 1).
Two notches with projecting blades of large anterior hamuli (Figure 6) in ventral distal part of haptor. Concave ventral surface of haptor divided into six loculi of unequal size and shape (Figure 1) and seventh central loculus. Two antero-lateral septa form big continuous septum along ventral surface of haptor (Figure 1). Accessory sclerites 0.38 (0.24–0.46) (Figure 14B), embedded in oval-shaped fibrous collars (Figures 5 & 6), with sturdy knife-shaped blade widening basally and two unequal roots, often one is barely developed (Figure 14B). Level of development of these roots variable in egg-bearing specimens. Fibrous collar surrounding accessory sclerites extending to muscular sac of corresponding anterior-posterior hamuli complex on each side. Anterior hamuli 0.51 (0.41–0.60) long (Figure 14B). Anterior hamulus smooth blade tip prominent (Figures 4 & 5), whole claw-shaped hamulus apparent through muscular- fibrillar tegument (Figures 4-6). Posterior hamulus adjoined to anterior hamulus (Figures 8 & 14B). Posterior hamuli 0.16 (0.13–0.23) long. Its ribbed blade tip (Figure 9) is also prominent, while base (handle) embedded in muscular- fibrous tissue. Marginal hooklet length 0.015 (Figure 7), on ventral surface of haptor. Haptor with complex musculature, and X-shaped tendons going through central part.
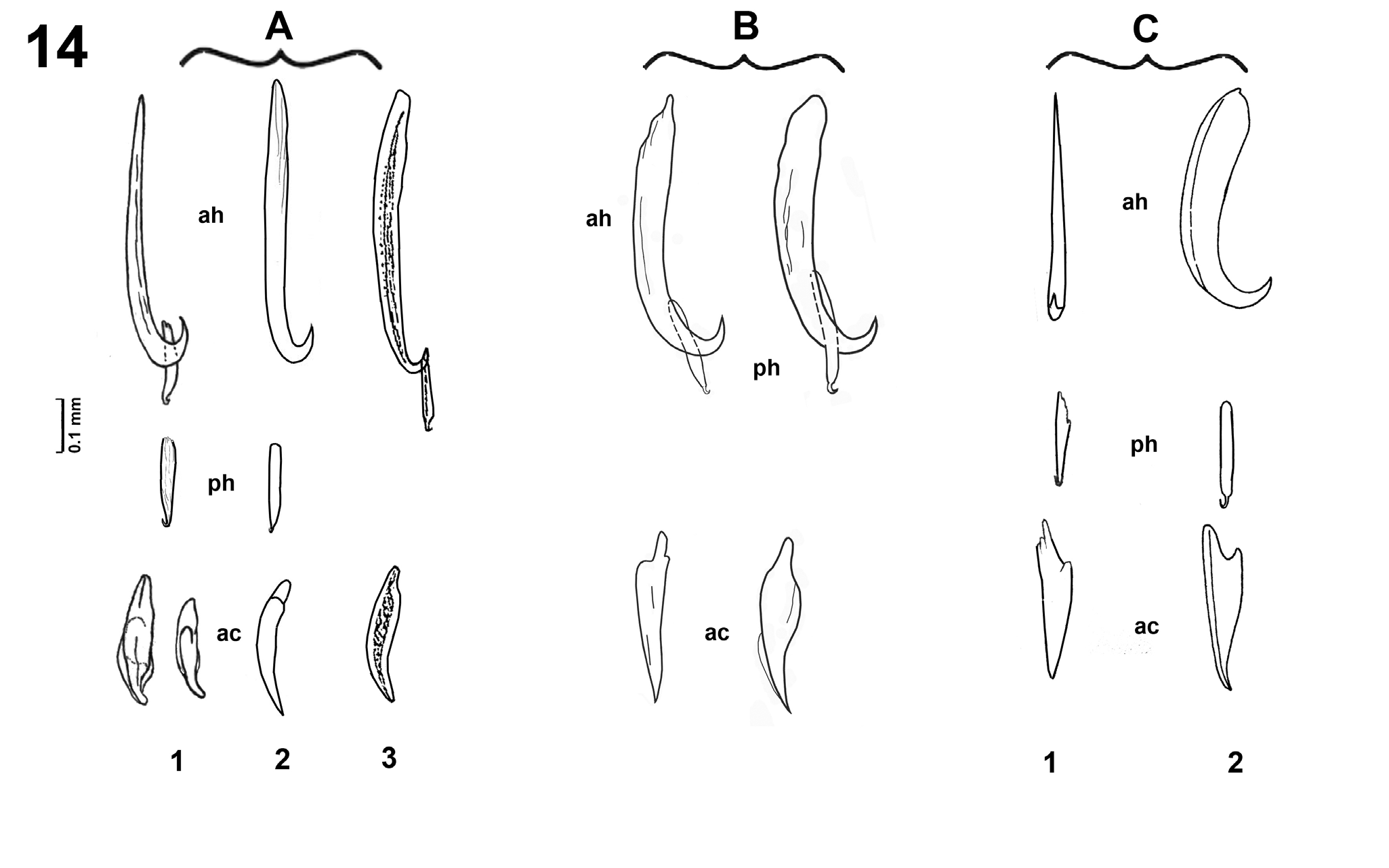
Figure 14: Comparison of shape of sclerotized structures of haptor (anterior and posterior hamuli and accessory sclerite) based on adaptation from published data and our data. A – “Pseudobenedenia nototheniae” type*.
- From Johnston, 1937, original description. Host Notothenia microlepidota.
- From Dollfus and Euzet,
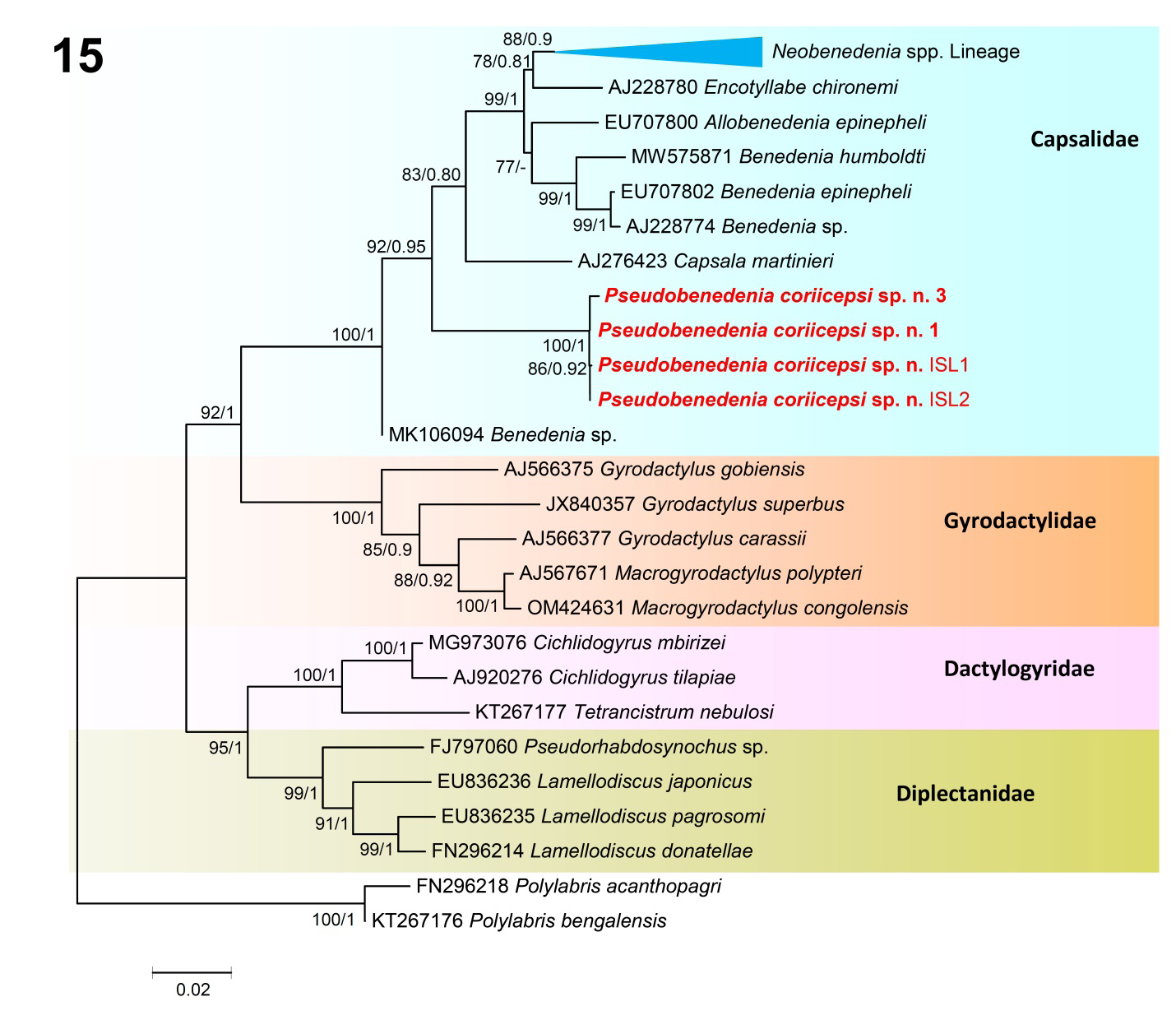
Figure 15: Phylogenetic relationship between species of the monogeneans based on the 18S dataset using Maximum likelihood (ML) and consensus Bayesian inference (BI). Numbers near internal nodes show maximum likelihood bootstrap percentage values and Bayesian posterior probabilities. GenBank accession numbers precede species names. Scale-bar indicates the expected number of substitution per site. Unsupported values of BI are shown by hyphen. Species sequenced in the present study are shown in bold. Families are indicated in the right side.
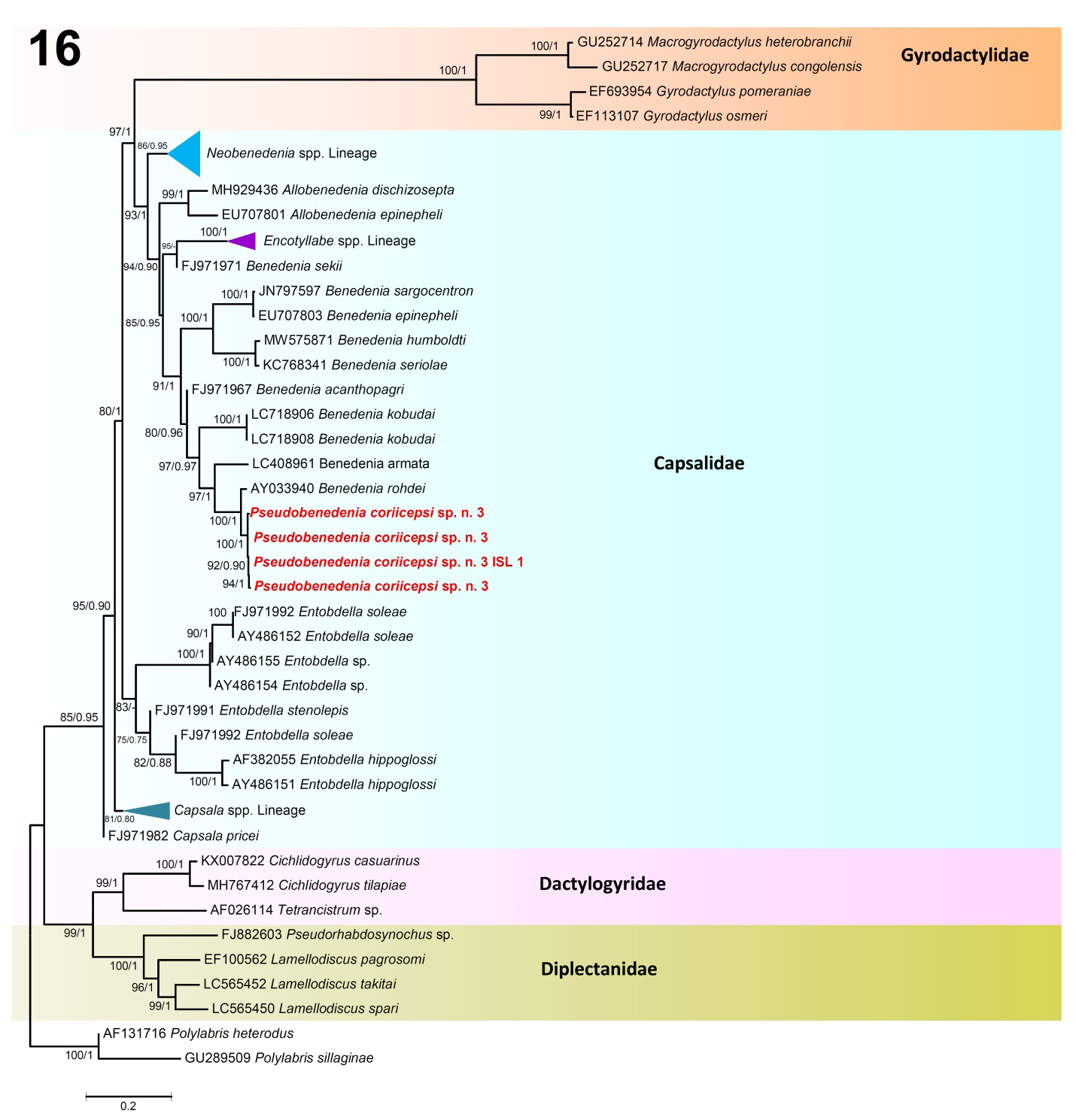
Figure 16: Phylogenetic relationship between species of the monogeneans based on the 28S dataset. Bootstrap values and Bayesian posterior probabilities are shown next to the nodes as ML/BI. Scale-bar indicates the expected number of substitution per site. Unsupported values of BI are shown by hyphen. The present study species are shown in bold. Families are indicated in the right side.
Differential Diagnosis
Pseudobenedenia coriicepsi sp. n. can be differenced from P. nototheniae described by Johnston [1, 3] by the different shape of anterior hamuli, anterior hamulus length to accessory sclerite length ratio 1.4 (versus 2.1 in P. nototheniae), and anterior hamulus length to anterior hamulus blade length ratio 6.9 (versus 4.8 in P. nototheniae), by a characteristic shape of vitellaria field with a v-shaped continuous commissure in the area between ovary and testes; non-lobed testes, with moderate perforations, almost central position of genital opening, smooth cuticle without fibers, smaller body measurements, including inner organs, and by different host species.
Remarks
Host identification is crucial in the designation of Monogenean species. Mistakes in host identification can lead to confusions when finding of monogenean on non-typical hosts, especially in the possible presence of hybrid host forms [26, 27]. The shape of sclerotized parts of the haptor has proven to be a consistent species characteristic easy to measure and having a stable shape as well as sclerotized structures of copulatory system, which have not been of utility in the family Capsalidae Baird, 1853 [27]. While analyzing published data and comparing morphological and metrical parameters of available literature on Pseudobenedenia, we paid close attention to these factors. Figure 14 shows an adaptation of drawings of sclerotized structures of haptor by similarity and by host species together with our new data. New species under study clearly differs from original description by more massive and robust blade of anterior hamulus. Also there is a clear difference in the shape of the accessory sclerite that is wider in the middle part and narrows more sharply to the tip. There are two roots in the accessory sclerite, one usually was almost invisible, and another more developed. For the comparison, we put three groups of P. nototheniae drawings close, and it’s obviously looks like the same species morphologically and were named as the same species in publications. Drawing A1 Figure 14 is Pseudobenedenia from N. microlepidota [3], drawing A2 is Pseudobenedenia from N. rossii and N. microlepidota (it is not clear from what species of host this parasite was collected) [8], and drawing A3 is Pseudobenedenia from unidentified species as well, but in this case the range of hosts was even wider (N. rossii, Trematomus sp., Parachaenichthys georgianus (Fisher), Champsocephalus gunnari Lönnberg, Harpagifer bispinis (Schneider) [10]. In our part, we studied material from N. coriiceps and found only shape B, but never shape A.
Figure 14C shows sclerotized structures of Pseudobenedenia from the one host N. neglecta, drawing C1 is from the original description of P. lauriei (not in a lateral view) [20], and C 2 is Pseudobenenia from the same host, a good lateral view [9]. It is hard to compare anterior hamuli, but accessory sclerite has large similarities, and definitely differs from the original description and from our new species. It is evident that the lateral view of the anterior hamulus in Willams, et al. [9] (Figure 14C2) demonstrates different shape wider and more robust, with different angle of blade to base position in it. So, the species from N. neglecta in Willams, et al. [9] is clearly different from P. nototheniae considering the drawings made in Johnston [3], Dollfus and Euzet [8] and Gibson [10] (Figures 14A1-3) and from new species collected from N. coriiceps (Figure: B). Accordingly, it is quite possible that Willams, et al. [9] were dealing with P. lauriei in their paper (which they somehow omitted in their list of references and the text of the paper).
The ratio between parts of hamuli is a reliable feature for species identification helps to distinguish species of Monogenea more precisely than variable measurements [31, 32, 33, 34, 35]. One should calculate ratios in each specimen, and later calculate the trending ratio. In our study, we used anterior hamulus length to accessory sclerite length ratio and anterior hamulus total length to anterior hamulus blade length ratio to describe P. coriicepsi sp. n. more precisely. To accomplish this, we used only specimens where all these structures were strictly positioned in profile and totally flattened. Comparative measurements of sclerotized structures of Pseudobenedenia species presented in Table 2 seem to overlap and do not show clear distinction between species from different fish hosts on many parameters. That is, possibly, one of the reasons of expanding of species complexes. However, using the ratio between the parts of hamuli gave us a good stable pattern, which proved that all species of Pseudobenedenia collected from N. coriiceps followed the same trend of ratios. The ratio between the parts of hamuli clearly distinguished P. coriicepsi sp. n. from P. nototheniae based on the description of Johnston [1, 3] (from two other host species), as well as from Pseudobenedenia species that Dollfus and Euzet [8] examined from N. rossi (considering their species as P. nototheniae) and from other Pseudobenedenia species described by other authors. This is sufficient justification that this feature is one of the most useful for morphometrical species diagnostics of Pseudobenedenia.
Using the light microscopy and by SEM, we did not find any fibers in mature or immature specimens on both sides of the body of P. coriicepsi sp. n. from N. coriiceps. These fibers were mentioned in the description of P. nototheniae by Johnston [1] as “extremely abundant”, as well as in Willams, et al. [9] in P. nototheniae from N. neglecta, but were not mentioned in Szidat`s short description of P. lauriei from N. neglecta [20].
Three prominent structures on the anterior end, two oral suckers and oral muscular mouth opening behind them gave this parasite its early name “tristoma” more than 110 years ago [42]. Also, numerous sensory glands surrounding the penis (Figure 12) have been registered in Pseudobenedenia and even for the family Capsalidae for the first time. Previously, these structures were studied in other groups of helminths and the pattern of their arrangement were considered to be very good species-specific characteristics by Amin, et al. [43].
It is impossible to make differential diagnosis of our new species with P. lauriei, P. shorti, as well as all species of Pseudobenedenia from the Hawaiian fish hosts since their descriptions miss crucial information on sclerotized structures of the haptor, but it differs from all above species by host species. If sclerotized structures of haptor are not positioned in profile, it makes difficult to see the morphology of the anchors and the accessory pieces of the haptor and can cause misdiagnoses.
We found a shape of the vitellaria field (that overlap intestine branches in mature specimens) also a useful feature to differentiate our new species from P. nototheniae. In case of P. nototheniae commissures of vitellaria have a branching pattern in the middle part of the body, in P. coriicepsi it is a clear v-shaped continuous commissure that is always between the ovary and the testes (Figure 1).
EDAX Results
The percent weights of phosphorus (P), sulfur (S), and calcium (Ca) of different parts of anterior hamulus and accessory sclerite in 2 specimens of P. coriicepsi sp. n. are detailed in Table 3. Phosphorus demonstrated the highest level in the cross-section part of anterior hamulus center, but low on the anterior body end and was absent on the haptor. The highest levels of sulfur were noted in the anterior hamulus center, and levels of Ca were higher in haptor.
| Phosphorus | Sulfur | Calcium | |
|---|---|---|---|
| Anterior body end | 0.25 | 0 | 1.45 |
| Haptor | 0 | 1.3 | 2.38 |
| Anterior hamulus edge | 0.01 | 0.01 | 0.67 |
| Anterior hamulus center | 0.44 | 8.83 | 1.95 |
Table 3: Composition of elements* in anterior hamulus and body parts of _Pseudobenedenia coriicepsi_ sp. n.
*Elements not measured. Data is based on % weights. Table 3: Composition of elements* in anterior hamulus and body parts of Pseudobenedenia coriicepsi sp. n.
Molecular Data and Phylogenetic Analyses
The phylogeny based on the 18S and 28S datasets (Figures 15 & 16) give a good diagnostic comparison among the representatives of Monogenea for which molecular data is available in GenBank, including the present species. Pseudobenedenia coriicepsi sp. n. in both datasets 18S and 28S was resolved in a separate and strongly supported clade (bootstrap values = 99–100%; posterior probability = 1.00) (Figures 15 & 16).
In the 18S phylogenetic tree based on ML and BI analysis, the taxa appeared to be subdivided into four main clades (Figure 15). Clade B included the gyrodactylid species of the genera Macrogyrodactylus Malmberg, 1957 and Gyrodactylus von Nordmann, 1832, forming a group with strong support (ML = 100; BI = 1.00) (Figure 15). They show a sister relationship with species of clade A that comprise the species belonging to the family Capsalidae represented by the genera Benedenia Diesing, 1858_, Capsala_ Bosc, 1811_, _Encotyllabe Diesing, 1850_, Allobenedenia_ Yamaguti, 1963_, and _Neobenedenia Yamaguti, 1963 along with isolates of Pseudobenedenia coriicepsi sp. n. clustering within a very well supported clade (100/1) (Figure 15). The dactylogyrids (clade C) contained parasites of the genera Cichlidogyrus Paperna, 1960 and Tetrancistrum Goto & Kikuchi, 1917 while their sister clade (D) comprises representatives of the genera Pseudorhabdosynochus Yamaguti, 1958 and Lamellodiscus Johnston & Tiegs, 1922 (Figure 15). The outgroup species of the genus Polylabris are allocated to the family Microcotylidae Taschenberg, 1879. The intraspecific genetic divergence among isolates identified as Pseudobenedenia coriicepsi sp. n. ranged from 0.00% to 0.02%. The genetic divergence among the isolates of Pseudobenedenia coriicepsi sp. n. with respect to capsalid monogeneans presented in the figure (Figure 15) ranged from 2.67% to 4.50%.
For the alignment of the 28S rDNA dataset, four newly generated sequences of Pseudobenedenia coriicepsi sp. n. along with other genera of the family Capsalidae retrieved from GenBank were used (Figure 16). The 28S analysis corroborated the 18S dataset and provided molecular confirmation for the identification of Pseudobenedenia coriicepsi sp. n. The phylogenetic analysis illustrated the groupings of the representatives present in the tree as shown in two main clades, A and B (Figure 16). Clade A was subdivided into A1 to A5 lineages; Lineage A1 was represented by species of gyrodactylid monogeneans; Lineage A2 received strong bootstrap and posterior probability values and was represented by species of the genera Neobenedenia, Allobenedenia, Encotyllabe, and Benedenia, also including the sequences obtained from Pseudobenedenia coriicepsi sp. n. (Figure 16). Pseudobenedenia coriicepsi sp. n. was resolved in a separate branch of the tree, but the species of Benedenia were found closely associated, joining this lineage. The A3 lineage formed a separate strongly supported lineage for the species of Entobdella, while the A4 and A5 lineages represented Capsala species (Figure 16). Furthermore, Clade B subdivided into lineages B1 and B2, in which dactylogyrids and diplectanids monogenean species clustered, respectively, and thus clearly highlights their separation. The newly generated sequences from isolates of Pseudobenedenia coriicepsi sp. n. clustered with strong support (100/1.00) and were sister to species of Benedenia. The genetic divergence among the isolates of Pseudobenedenia coriicepsi sp. n. was 0.00% to 0.03%. The distance values between the present sequences of Pseudobenedenia coriicepsi sp. n. and the sequences of Benedenia species ranged from 1.45% to 2.18%, while the Pseudobenedenia coriicepsi sp. n. 28S sequences showed genetic divergence with the closest Benedenia species, B. rohdei from Lutjanus carponotatus from Heron Island, Great Barrier Reef, Queensland, Australia, at 1.45% (Figure 16).
Discussion
One of the most important characteristics for species definition in monogeneans is their strict host specificity [26]. Nevertheless, more than 11 fish species were reported to be hosts for P. nototheniae [12, 13]. However, ambiguity in distinguishing of hosts of species of Pseudobenedenia in some publications is common. This issue was first noted in the original descriptions of P. nototheniae [1, 3] stating that the material was obtained from at least two different species of fish (N. angustata and N. colbecki) where N. angustata was considered to be a synonym of N. macrocephala. We can thus assume that the material for the original description of P. nototheniae could have been based on a mixture of at least two species of fish hosts. Further historical records of Pseudobenedenia suggests two possibilities (1) the descriptions of new species from new fish hosts, (2) the identification of capsalids on other hosts together with notes of new features that were missing in the original description of P. nototheniae. For example, presence of the 7th septa of the haptor of “P. notothenia” from N. rossi [8], or extending the range of metric variability of this species to numbers that differ 3-4 fold [10]. The second group of researchers expanded Pseudobenedenia species complex with a wide range of hosts and significantly blurred the diagnostic features of this group of Monogeneans. Another disturbing example – while describing the new species of P. shorti, the authors mentioned: “Though we have indicated Rhigophola deaborni as a host of this species it is likely that the record on which it is based is erroneous” [21]. Later, P. shorti was synonymized to Pseudobenedenoides shorti by Gibson [10]; we are not discussing the correctness of this synonymy in this paper.
We assume now Johnston [1, 3] in his original descriptions of P. nototheniae was not only dealing with worms from different fish hosts but more likely he also used a mixture of juvenile and adult worms because he mentioned the size range of P. nototheniae covering the smallest worms and the largest worms. This problem with the first species descriptions led to much confusion in other studies of Monogenea making one of the reasons of expanding the species complexes, that we here assumed to be P. nototheniae. There are few other cases in capsalid monogeneans that exhibited a broad host specificity, apparently caused by the reasons mentioned above [44, 45, 46].
Gibson [10] reported the presence of P. nototheniae in a wide range of fish hosts, namely in N. rossi Richardson, Trematomus sp., Parachaenichthys georgianus, Champsocephalus gunnari Lonnberg, Harpagifer bispinis (Schneider). He mentioned that this material was collected by a large number of collectors and “not least amongst these workers have been helminthologists”. Whittington [46], in his review of the diversity of Capsalidae gave a few examples of well-known Neobenedenia “melleni” identity issues and stated that known capsalids with low host specificity could be revealed during detailed studies as a complexes of different species. Thus, Timofeeva, et al. [11] while studying the capsalids from 23 species of nototheniid fishes from Atlantic part of Antarctica and Subantarctica and analyzing the details of morphology of sclerotized structures, host specificity, and details of reproductive system, discovered three new species – P. gibberifrons, P. dissostichii and Pseudobenedeniella branchialis Timofeeva, Gaevskaja et Kovaliova, 1987 from different nototheniid hosts.
Willams, et al. [9] collected the specimens of Pseudobenedenia from N. neglecta and were the first researchers of this group who paid a proper attention to such critical details of morphology of Monogeneans as sclerotized structures of haptor. They mentioned that Johnston’s description [1, 3] and Dollfus and Euzet [8] presented “lack the detailed numerical data”, thus, “the comparison with their newly collected material “was inappropriate”, which we certainly agree with. Willams, et al. [9] noted that though body anatomy had been studied very thoroughly in the works of Johnston [1, 3], and later by Dollfus and Euzet [8], the details of opisthohaptor structures “remain uncertain and…conflicting”. Somehow Willams, et al. [9] omitted in their work the description of P. laureri by Szidat [20], erroneously mentioning that their work was the first record of Pseudobenedenia from Notothenia neglecta.
Following the nomenclature of Llewellyn [47], we named the sclerotized structures of haptor: anterior pair as accessory sclerites, the middle pair as anterior hamuli, and posterior pair as posterior hamuli (Figures 4-6). In the first description of P. nototheniae, Johnston [1] did not provide any graphic material for sclerotized structures of haptor, only their metrical characteristic, with some misprinted data; he corrected the information on sizes of accessory sclerite in the latter publication [3]. Later, Johnston provided drawings of sclerotized structures in the second description [3], but he named them differently as hook 1 (for accessory sclerite), hook 2 (for anterior hamulus) and hook 3 (for posterior hamulus). Later in literature, some authors blindly repeated this order but mistakenly named accessory sclerites as “anterior hooks”, anterior hamuli as ‘central hooks” [24] that we corrected here in Table 2. Secretory gland cells of the haptor were noticed with the SEM (Figure: 4); they have not been reported for Pseudobenedenia before [9].
Analyzing previously published data on the prevalence, intensity and abundance of Pseudobenedeniae specimens collected from N. coriiceps and from other teleost fish species at the vicinity of Galindez Island [4, 5, 48, 49] as well as from the South Orkney Islands region [50], we can conclude that out of 14 fish species examined during 2014–2022, the monogeneans of genus Pseudobenedenia were found only in fish of the genus Notothenia – mostly in N. coriiceps and very rarely in N. rossii. Our preliminary results concerning Pseudobenedenia specimens collected from N. rossii requires further research.
Therefore, in addition to the mentioned above, we consider the current taxonomic status of P. nototheniae as species complex that needs further research using fresh material from the type host, N. microlepidota. Yamaguti [22] described P. elongate, P. merithe and P. ovalis from Hawaiian fishes, but since any detailed information on haptor structures was lacking in his description, it is impossible to use those species for reliable comparisons. Moreover, two of these species have a few fish hosts, some of them were not identified to the species, and some of the described species were found on gills of fish; thus, we suppose that these tropical Pseudobenedenia species also need thorough re- examination in future.
We assume that in the case of Monogeneans collected from Antarctic fish, this is very rare that these helminths are examined by a specialist on monogeneans. Taking into account the specifics of this group of helminths, we emphasize the importance of using only mature monogenean specimens in species descriptions, and on application of uniform fixation techniques because wide range of helminth fixation methods, improper identification of fish hosts and discrepancy of other approaches eventually lead to misidentifications of Antarctic monogeneans or puffing of species complexes.
Data for P. coriicepsi sp. n. that we received in this study with EDXA are collected for the first time for Capsalidae, but were made for other groups of Monogenea before [51, 52]. In this paper we are establishing the baseline on this data for future comparisons for this group of Monogeneans, these are descriptive taxonomic characters same like morphometric or molecular data [53].
To the best of our knowledge, this is the first molecular study for any Pseudobenedenia species, as no data on these monogeneans is available on the GenBank. Inclusively, our study initiates the data generation for 18S and 28S sequences for Pseudobenedenia species. Both 18S and 28S sequence analyses and phylogenetic reconstructions confirmed the validation of P. coriicepsi sp. n., including its morphological identification, and confirmed Pseudobenedenia as a genus under Capsalidae. The phylogenetic analyses inferred from two molecular markers, 18S and 28S, in the present study reliably showed that the new species P. coriicepsi sp. n. is closely related to the other species of the genus Benedenia. For a more robust phylogeny and to better understand the relationships among the Capsalidae, additional sequence evidence from extensive taxon sampling is required. Finally, further molecular studies with vast additional sampling will tackle the relationships among the members of the Capsalidae and the identification of this group of parasites.
Acknowledgments
The Department of Biology, Brigham Young University (BYU), Provo, Utah, USA and the Institute of Parasitic Diseases (PCI), Scottsdale, Arizona supported this project. We thank Michael Standing, Electron Optics Laboratory (BYU) for his technical help and expertise. Our special thanks go to the librarian Rhiannon Bell, Hamilton College, Clinton, NY, USA and to Dr. Sara M. Rodríguez, Austral University, Valdivia, Chile for their help with rare manuscripts. We are grateful to the Head, Department of Zoology, Chaudhary Charan Singh University, Meerut, Uttar Pradesh, India for providing laboratory facilities. We are thankful to Dr. Gabor R. Racz, parasitology collection manager of University of Nebraska State Museum, Lincoln, Nebraska, USA for his help with museum collections.
References
-
Johnston TH (1931) New Trematodes from the Subantarctic and Antarctic. Austr J Experim Biol Medical Sci 8: 91-98.
-
Waite ER (1916) Fishes. Scientific reports, (Australasian Antarctic Expedition (1911 - 1914). Ser C Zool and Botany. Government Printer, Adelaide 3: 1.
-
Johnston TH (1937) Trematoda. Australasian Antarctic Expedition 1911-1914 10(1): 1-29.
-
Kuzmina TA, Salganskij OO, Lisitsyna OI, Korol EM (2020) Helminths of Antarctic rockcod Notothenia coriiceps (Perciformes, Nototheniidae) from the Akademik Vernadsky Station area (Argentine Islands, West Antarctica): new data on the parasite community. Zoodiversity 54(2): 99-110.
-
Kuzmina TA, Dykyy IV, Salganskij OO, Lisitsyna OI, Korol EM, et al. (2021) Helminth diversity in teleost fishes from the area of the Ukrainian Antarctic station “Akademik Vernadsky”, Argentine Islands, West Antarctica. Zoodiversity 55(3): 251-264.
-
Johnston TH, Mawson PM (1943) Endoparasites from the subantarctic islands of New Zealand. Rec S Austral Mus 7(3): 237-243.
-
Johnston TH, Mawson PM (1953) Parasitic nematodes and trematodes from Campbell and Auckland Islands (Cape Expedition). Rec Dominion Mus 2 (2): 63-71.
-
Dollfus RF, Euzet L (1965) Supplement to the description of _Pseudobenedenia nototheniae_ T.H. Johnston, 1931 (monogenetic trematode) parasite of a teleost of the genus _Notothenia_ Richardson from the Kerguelen Island. Bull Mus Nation D’Hist Natur 2: 36(6): 849-857.
-
Williams IC, Ellis C, Spaull VW (1973) The structure and mode of action of the posterior adhesive organ of _Pseudobenedenia_ _nototheniae_ Johnston, 1931 (Monogenea: Capsaloisea). Parasitol 66(3): 473-485.
-
Gibson DI (1976) Monogenea and Digenea from fishes. Discov Rep 36: 179-266.
-
Timofeeva TA, Gaevskaya AV, Kovaleva AA (1987) Capsalids (Monogenea) of the notothenioid fishes from the Atlantic region of Antarctica and Subantarctica. Tr Zool Inst 161: 78-93.
-
Oğuz MC, Tepe Y, Belk MC, Heckmann RA, Aslan B, et al. (2015) Metazoan parasites of Antarctic fishes. Turkiye Parazitol Derg 39(2): 174-178.
-
Klapper R, Münster J, Kochmann J, Klimpel S, Kuhn T (2017) Biodiversity and host specificity of Monogenea in Antarctic fish species, in: Biodiversity and evolution of parasitic life in the Southern Ocean. Paras Res Monog, ed by: Klimpel S, Kuhn T, Heinz Mehlhorn H. Springer Cham 9: 33-48.
-
Elchmeyer Catalog of fishes.
-
Froese R, Pauly D (2023) FishBase. World Wide Web electronic publication.
-
Muñoz G, Rebolledo M (2018). Comparison of the parasite community of two notothens, _Notothenia rossii_ and _N. coriiceps_ (Pisces: Nototheniidae), from King George Island, Antarctica. Journal of Helminthology 93(6): 732-737.
-
Bychowsky BE, Nagibina LF (1967) New Capsalidae (Monogenoidea) from Pacific fishes Parazitologiia. 1(6): 521-528.
-
Hine PM, Jones JB, Diggles BK (2000) A checklist of parasites of New Zealand fishes, including previously unpublished records. National Institute of Water and Atmospheric Res Techn Rep no 75 Wellington.
-
Szidat L (1965) Estudios sobre la fauna de parasitos de peces antarticos. I. Los parasitos de _Notothenia neglecta_ Nybelin. Servic Hidrogr Naval Secr Mar de la Rep Argent 910: 1-84.
-
Hargis WJJr, Dillon WA (1968) Helminth parasites of Antarctic fishes: the superfamily Capsaloidea Price, 1931. Proc Biol Soc Wash 81:403-412.
-
Yamaguti S (1968) Monogenetic trematodes of Hawaiian fishes. University of Hawaii Press, Honolulu, pp: 287.
-
Lawler AR (1981) Zoogeography and host-specificity of the superfamily Capsaloidea Price, 1936 (Monogenea: Monopisthocotylea). Spec Pap Mar Scien 6: 216-217.
-
Lutnicka H, Zdzitowiecki K (1984) On some problems related to the occurrence of _Pseudobenedenia_ _nototheniae_ Johnston, 1931 (Monogenea, Capsalidae, Trochopodinae) off the South Shetland Islands. Act Ichthyol Piscator 14: 141-147.
-
Bykhovsky BE (1957) Monogenetic flukes. Their system and phylogeny. Acad Sci USSR Moscow, Leningrad, pp: 509.
-
Gusev AV (1983) Methods for collection and preparation of monogeneans parasitizing fish. Leningrad: Nauka, USSR, pp: 46.
-
Gusev AV (1985) Class Monogenea. In: Bauer ON (Ed.), Keys to parasites of freshwater fish of fauna of USSR Vol 2 Parasitic multicellular Leningrad, USSR Nauka, pp: 425.
-
Poulin R (2002) The evolution of monogenean diversity. Int J Parasitol 32(3): 245-254.
-
Whittington ID, Deveney MR, Wyborn SL (2001) A revision of _Benedenia_ Diesing, 1985 including a redescription of _B. sciaeniae_ (van Beneden, 1856) Odhner, 1905 and recognition of Menziesia Gibson, 1976 (Monogenea: Capsalidae). J Nat Hist 35(5): 663-777.
-
Weber EP III, Govett P (2009) Parasitology and necropsy of fish. Compendium on Continuing Education for the Practicing Veterinarian 31(2): E12.
-
Euzet L, Combes C, Batchvarov G (1974) Sur un nouveau Polystomatidae Européen, parasite de l’amphibien _Bufo_ _viridis._ Laur Vie Milieu 24: 129-139.
-
Buchvarov G (1980) _Polystoma mazurmovici_ n sp (Polystomatidae Gambl, 1896), a parasite of _Rana_ _dalmatina_ Bonaparte, 1839 in Bulgaria. Sci Stud Univof Plovdiv Biol 18(4): 183-190.
-
Buchvarov GK (1984) _Polystoma skrjabini_ n sp (Polystomatidae Gambl, 1896) - a parasite of _Hyla_ _arborea_ (Linne, 1758), in Bulgaria. Compt l`Acad Bulg Scien 37(5): 697-699.
-
Rubtsova N Yu, Heckmann RA (2017) Morphological and structural differences of normal adult and sub-adult bladder forms of _Polystoma integerrimum_ (Fröhlich, 1798) (Monogenea: Polystomatidae) from the common frog, _Rana temporaria_. Sci Parasitol 18(1-2): 38-53.
-
Rubtsova N Yu (2018) Redescription and variability of _Polystoma mazurmovici_ and _P. skuratovitchi_ (Monogenea, Polystomatidae), with a key to Polystoma from Anurans of Ukraine. Vestn Zool 52(2): 91-100.
-
Lee RE (1993) Scanning Electron Microscopy and X-ray Microanalysis. Prentice Hall, Englewood Cliffs NJ, pp: 458.
-
Tamura K, Glen S, Kumar S (2021) MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol Biol Evol 38: 3022-3027.
-
Sela I, Ashkenazy H, Katoh K, Pupko T (2015) GUIDANCE2: accurate detection of unreliable alignment regions accounting for the uncertainty of multiple parameters. Nucl Acids Res 43: 7-14.
-
Milne I, Lindner D, Bayer M, Husmeier D, McGuire G, et al. (2009) TOPALI v2: a rich graphical interface for evolutionary analyses of multiple alignments on HPC clusters and multi-core desktops. Bioinform 25: 126- 127.
-
Darriba D, Taboada GL, Doallo R, Posada D (2012) ModelTest 2: More models, new heuristics and parallel computing. Natur Methods 9: 772.
-
Euzet L, Cauwet A (1967) _Polylubris diplodi_ n g n sp (Monogenea, Microcotylidae), parasite de Téléostéens du genre _Diplodus_ Sparidae. Bull Mus Nat Hist Nat 39(1): 213-220.
-
Waite ER (1909) Vertebrata of the subantarctic islands of New Zealand. In Chilton C. (Ed.) The Subantarctic Islands of New Zealand. Vol II Wellington, Philos Insto Canterb pp: 542-600.
-
Amin OM, Rubtsova NYu, Heckmann RA (2020) Accessory copulatory structures in the bursa of male acanthocephalans. Sci Parasitol 21(3): 102-119.
-
Ogava K, Bondad-Reantaso MG, Wakabayashi H (1995) Redescription of _Benedenia epinepheli_ (Yamaguti, 1937) Mesere, 1938 (Monogenea: Capsalidae) from cultured and aquarium marine fishes of Japan. Can J Fish Aquat Sci 52: 62-70.
-
Garcia RGGF, Pradi-Garcia MM, del Vall MT, Rodriguez- Diego JG (2000) Nuevas especies de hospederos y localizacion para _Pseudobenedenia_ _nototheniae_ y _Pseudobenedenoides shorti_ (Monogenea: Capsalidae) en peses antarcticos. Rev Salud Anim 22: 61-63.
-
Whittington ID (2004) The Capsalidae (Monogenea: Monopisthocotylea): a review of diversity, classification and phylogeny with a note about species complexes. Fol Parasitol 51: 109-122.
-
Llewellyn J (1963) Larvae and larval development of monogeneans. Adv Parasitol 1: 287-326.
-
Kuzmina TA, Laskowski Z, Salganskij OO, Zdzitowiecki K, Lisitsyna OI, et al. (2022) Helminth assemblages of the Antarctic black rockcod, _Notothenia coriiceps_ (Actinopterygii: Nototheniidae) in coastal waters near Galindez Island (Argentine Islands, West Antarctic): temporal changes in the endoparasite community. Act Parasitol 67(1): 207-217.
-
Syrota YY, Kuzmin YI, Lisitsyna OI, Salganskiy OO, Dykyy IV, et al. (2023) Infection patterns of helminth community in black rockcod _Notothenia coriiceps_ in West Antarctica over a 6-year term. Parasitol Res 122(3): 853-865.
-
Kuzmina T, Salganskiy O, Vishnyakova K, Ivanchikova J, Lisitsyna O, et al. (2022) Helminth diversity in teleost fishes from the South Orkney Islands region, West Antarctica. Zoodiversity 56(2): 135-152.
-
Rubtsova NYu, Heckmann RA, Smit WS, Luus-Powell WJ, Halajian A, et al. (2018) Morphological studies of developmental stages of _Oculotrema hippopotami_ (Monogenea: Polystomatidae) infecting the eye of _Hippopotamus amphibius_ (Mammalia: Hippopotamidae) using SEM and EDXA with notes on histopathology. Kor J Parasitol 56(5): 463-475.
-
Rubtsova NYu, Heckmann RA (2019) Structure and morphometrics of _Ancyrocephalus paradoxus_ (Monogenea: Ancyrocephalidae) from _Sander lucioperca_ (Percidae) in Czechia. Helminthol 56(1): 11-21.
-
Amin OM, Heckmann RA, Sharifdini M, Albayati NY (2019) Moniliformis cryptosaudi n. sp. (Acanthocephala: Moniliformidae) from the long-eared hedgehog _Hemiechinus auritus_ (Gmelin) (Erinaceidae) in Iraq, a case of incipient cryptic speciation related to _M. saudi_ in Saudi Arabia. Acta Parasitol 64: 195-204.
-
Olson PD, Littlewood DT (2002) Phylogenetics of the Monogenea-evidence from a medley of molecules. Int J Parasitol 32: 233-244.
-
Plaisance L, Littlewood DTJ, Olson PD, Morand S (2005) Molecular phylogeny of gill monogeneans (Platyhelminthes, Monogenea, Dactylogyridae) and colonization of Indo-West Pacific butterflyfish hosts (Perciformes, Chaetodontidae) Zool Scr 34: 425-436.
-
Littlewood DTJ, Curini-Galletti M, Herniou EA (2000) The interrelationships of Proseriata (Platyhelminthes: Seriata) tested with molecules and morphology. Mol Phylogenet Evol 16: 449-466.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind