New Annotated Records of Helminth Parasites with Light Microscopy II. Acanthocephala (Polymorphida)
We give an account of 8 species of polymorphid acanthocephalans in 3 families collected from 14 various host species in different geographical locations. Each species account is annotated and morphologically presented using light microscopy showing characteristic diagnostic features. None of these accounts has been previously published. All geographical records and many host associations are new. This treatment includes Centrorhynchus spinosus (Kaiser, 1893) adults from Strix aluco Linn. In Iran and cystacanths from Osteopilus septentrionalis (Duméril & Bibron) in Florida; Sphaerirostris lancea (Westrumb, 1821) from Vanellus vanellus (Linn.) in Ukraine; Sphaerirostris picae (Rudolphi, 1819) Golvan, 1956 from various hosts in Ukraine, Iran, Bulgaria and Egypt (Centrorhynchidae); Plagiorhynchus (Prosthorhynchus) cylindraceus (Goeze, 1782) Schmidt & Kuntz, 1966 from Turdus merula Linn. in Iran and Sturnus vulgaris Linn. in Wisconsin; Plagiorhynchus (Prosthorhynchus) nicobarensis Soota & Kansal, 1970 from S. vulgaris in Iran (Plagiorhynchidae); Andracantha gravida (Algret, 1941) Schmidt, 1975 from Gavia immer (Brunnich) in Florida; Arhythmorhynchus brevis Van Cleave, 1916 from Casmerodius albus (Linn.) in Florida, and Southwellina hispida (Van Cleave, 1925) Witenberg, 1932 (= Polymorphus ardeae Belopolskaya, 1958) from Ardea cinerea Linn. in Macedonia and from Nycticorax violaceus (Linn.) in Florida.
Omar M Amin* and Nataliya Y Rubtsova
Keywords: Acanthocephala; Polymorphida; Wildlife; Descriptions; Distribution; Light Microscopy
Introduction
We have collected thousands of parasitic organisms from vertebrates over the years as part of an overall research program from North America and elsewhere in the world. Other collections were gifted to us for diagnosis or for joint research efforts. Significant collections were published. A few others were just saved and put aside after having been processed as whole mounts as reference material for future examination. We have started this series of investigations by exploring a small group of digeneans, cestodes, and nematodes that have not been previously studied or published by Amin and Rubtsova [1]. The polymorphid acanthocephalans are the subject matter of this study.
We have studied morphometrically, geographically, and historically adults and cystacanths of Centrorhynchus spinosus (Kaiser, 1893) from birds and amphibians, respectively, and described males from Iran. We have similarly studied Sphaerirostris lancea (Westrumb, 1821) and described males and females from Ukraine as well as 4 populations of another centrarchid acanthocephalan, S. picae (Rudolphi, 1819), from Ukraine, Iran, Bulgaria, and Egypt to evaluate its intraspecific variability. Our few specimens of Plagiorhynchus cylindraceus (Goeze, 1782) from the US and from Iran are compared with those from South Africa, Russia, Ukraine, and Taiwan. Plagiorhynchus (Prosthorhynchus) nicobarensis Soota and Kansal, 1970 was studied from a new population in Iran that varied from its original Indian specimens and our material of Andracantha gravida (Algret, 1941) Schmidt, 1975 showed marked variations from those described from Cuba and Florida. Our specimens of Southwellina hispida (Van Cleave, 1925) Witenberg, 1932 were originally described as Polymorphus ardeae Belopolskaya, 1958 from Ardea cinerea Linn. in Macedonia and we have also described more specimens from Nycticorax violaceus (Linn.) in Florida. The ecological implications of this taxonomic treatment are inescapable. The new host records in added geographical locations often unrelated to those of original descriptions reflect adaptations to a wider range of host specializations in different environments.
Materials and Methods
The specimens reported in this presentation were collected using routine methods for the examination of relevant hosts. For microscopical examination, specimens were placed in water overnight or until fully extended then fixed in 70% ethanol. Acanthocephalans were punctured with a fine needle and subsequently stained in Mayer’s acid carmine, de-stained in 4% hydrochloric acid in 70% ethanol, dehydrated in ascending concentrations of ethanol (24 hr each), and cleared in 100% xylene then in 50% Canada balsam and 50% xylene (24 hr each). Whole worms were then mounted in Canada balsam. Optical microscope images were acquired using a BH2 light Olympus microscope (Olympus Optical Co., Osachi-shibamiya, Okaya, Nagano, Japan) attached to an AmScope 1000 video camera (United Scope LLC, dba AmScope, Irvine, California), linked to an ASUS laptop equipped with HDMI high-definition multimedia interface system (Taiwan-USA, Fremont, California). Measurements are in micrometers unless otherwise noted; the range is followed by the mean values between parentheses when appropriate. Width measurements represent maximum width.
Results
Eight species of acanthocephalans reported from vertebrate hosts in 14 different geographical regions of the world are listed in Table 1 along with their hosts and localities from where they were collected. Morphological notes along with light microscopy images of major diagnostic features are included. Collectors’ information when known and dates are also included.
Centrorhynchus spinosus (Kaiser, 1893) Van Cleave, 1924
Polymorphida (Centrorhynchidae) (Figures 1, 2) Van Cleave [2] (p. 513) corrected the record of C. spinosus in his following statement “Echinorhynchus spinosus was originally described by Kaiser from an unrecorded host from Florida.” The present writer (Van Cleave) [3], misinterpreting Kaiser’s description, described and renamed the same species Centrorhynchus spinosus (based on one female_). Several birds, all from Florida, have been listed as hosts for this species.” The acanthocephalan has been since found in Russia [4] and Van Cleave [2] also described females from the “Galapagos hawk”. We have 2 males and one fragmented gravid female collected from _Strix aluco Linn. by Dr. Ali Halajian in Iran representing new records. All internal structures of the female were obscured by eggs. This appears to be a host-specific parasite found in only very few hosts including the great grey owl Strix nebulosa Forster in Russia [4].
| Parasite | Host | Distribution |
|---|---|---|
| Centrorhynchidae | ||
| Centrorhynchus spinosus (Kaiser, 1893) Van Cleave, 1924 | Strix aluco, Linn, tawny owl | Iran |
| Centrorhynchus spinosus (Kaiser, 1893) Van Cleave, 1924 cystacanth | Osteopilus septentrionalis (Duméril & Bibron), Cuban tree frog | Tampa, Florida |
| Sphaerirostris lancea (Westrumb, 1821) | Vanellus vanellus (Linn.), Northern lapwing | Ukraine |
| Sphaerirostris picae (Rudolphi, 1819) Golvan, 1956 | Pica pica (Linn.), Eurasian magpie and Corvus sp., crows & ravens | Ukraine |
| Sphaerirostris picae (Rudolphi, 1819) Golvan, 1956 | Pica pica (Linn.), Eurasian magpie | Bulgaria |
| Sphaerirostris picae (Rudolphi, 1819) Golvan, 1956 | Corvus corone Linn., Carrion crow & Corvus frugilegus Linn., Rook | Iran |
| Sphaerirostris picae (Rudolphi, 1819) Golvan, 1956 | Corvus corone Linn., Carrion crow | Nile Delta, Egypt |
| Plagiorhynchidae | ||
| Plagiorhynchus (Prosthorhynchus) cylindraceus (Goeze, 1782) Schmidt & Kuntz, 1966 | Turdus merula Linn., blackbird | Iran |
| Plagiorhynchus (Prosthorhynchus) cylindraceus (Goeze, 1782) Schmidt & Kuntz, 1966 | Sturnus vulgaris Linn., common starling | Kenosha, Wisconsin |
| Plagiorhynchus (Prosthorhynchus) nicobarensis Soota & Kansal, 1970 | Sturnus vulgaris Linn., common starling | Iran |
| Polymorphidae | ||
| Andracantha gravida (Algret, 1941) Schmidt, 1975 | Gavia immer (Brunnich), common loon | Franklin Co., Florida |
| Arhythmorhynchus brevis Van Cleave, 1916 | Casmerodius albus (Linn.), great egret | Florida |
| Southwellina hispida (Van Cleave, 1925) Witenberg, 1932 | Ardea cinerea Linn., grey heron | Nestos River Delta, East Macedonia |
| Southwellina hispida (Van Cleave, 1925) Witenberg, 1932 | Nycticorax violaceus (Linn.), yellow- crowned night heron | Snell Island, Florida |
Table 1: Acanthocephala (Polymorphida) reported from wildlife in this paper and their hosts and geographical distribution.
Available measurements of females especially those by Van Cleave [2, 5, 6] were confused giving trunk length of 20.00-60.00 mm, proboscis length of 0.65-1.00-1.30 mm, 30- 34 proboscis hook rows of 22-24 hooks each measuring 38- 48-60 long anteriorly and 50 posteriorly. Van Cleave [5] gave a count of 24 hooks per row but reported in 1924 9-12 rooted hooks anteriorly and 14-16 spine-like hooks posteriorly. He [6] (p. 296) emphasized “the great variability in the size of the proboscis hooks and thorns..…that were even continuous in the same females.” The determination of C. spinosus from India by Pujatti [7] was apparently in error as recognized by Golvan [8]. Similarly, Schmidt and Neiland [9] also recognized erroneous identifications of three Old World parasites of hawks that have the same number and arrangement of hooks as C. spinosus. Yamaguti (1963, p.126) referred to 30.00-45.00 mm long males and Nickol’s [10] description of C. spinosus summarized previous taxonomic treatments and is used here as a reference point for comparison. Richardson and Nickol [11] provided a useful key to species of Centrorhynchus in North America which identifies C. spinosus with 8-11anterior hooks in each longitudinal row and with trunk lacking anterior conspicuous inflation which agrees with our specimens. Similarly, Khokhlova [12] discussed the status of C. spinosus in the USSR and provided a key of the 15 known species of Centrorhynchus there.
First Description of Male Centrorhynchus spinosus from Strix aluco in Iran: Based on two mature adults; one is fragmented. Trunk long, cylindroid, slightly inflated anteriorly, 23.37 mm long by 0.92 wide anteriorly. Proboscis (Fig. 1) 0.95-1.09 long. Anterior proboscis 525-650 long by 333-354 wide with 9-14 rooted hooks per row measuring maximum of 52-75 anteriorly and becoming most robust at middle and transitional posteriorly. Posterior proboscis 315-425 long by 364-385 wide with 8-9 curved spiniform hooks per row measuring maximum of 45-48 anteriorly. Internal anatomy of proboscis shows receptacle process (Fig. 1). Proboscis receptacle double walled, 1.30-1.37 mm long by 0.27 mm wide with cephalic ganglion near middle. Lemnisci longer than receptacle, 2.17 mm long by 0.16 mm wide. Testes close but not contiguous, 0.62-1.07 mm long by 0.40-0.50 m wide, pre-equatorial, 4.50 mm from anterior trunk. Four tubular cement glands 11.25 mm long by 0.42 mm wide anteriorly. Saefftigen’s pouch 2.80 mm long. Eggs oblong with concentric shells and some polar prolongation of fertilization membrane (Fig. 2).
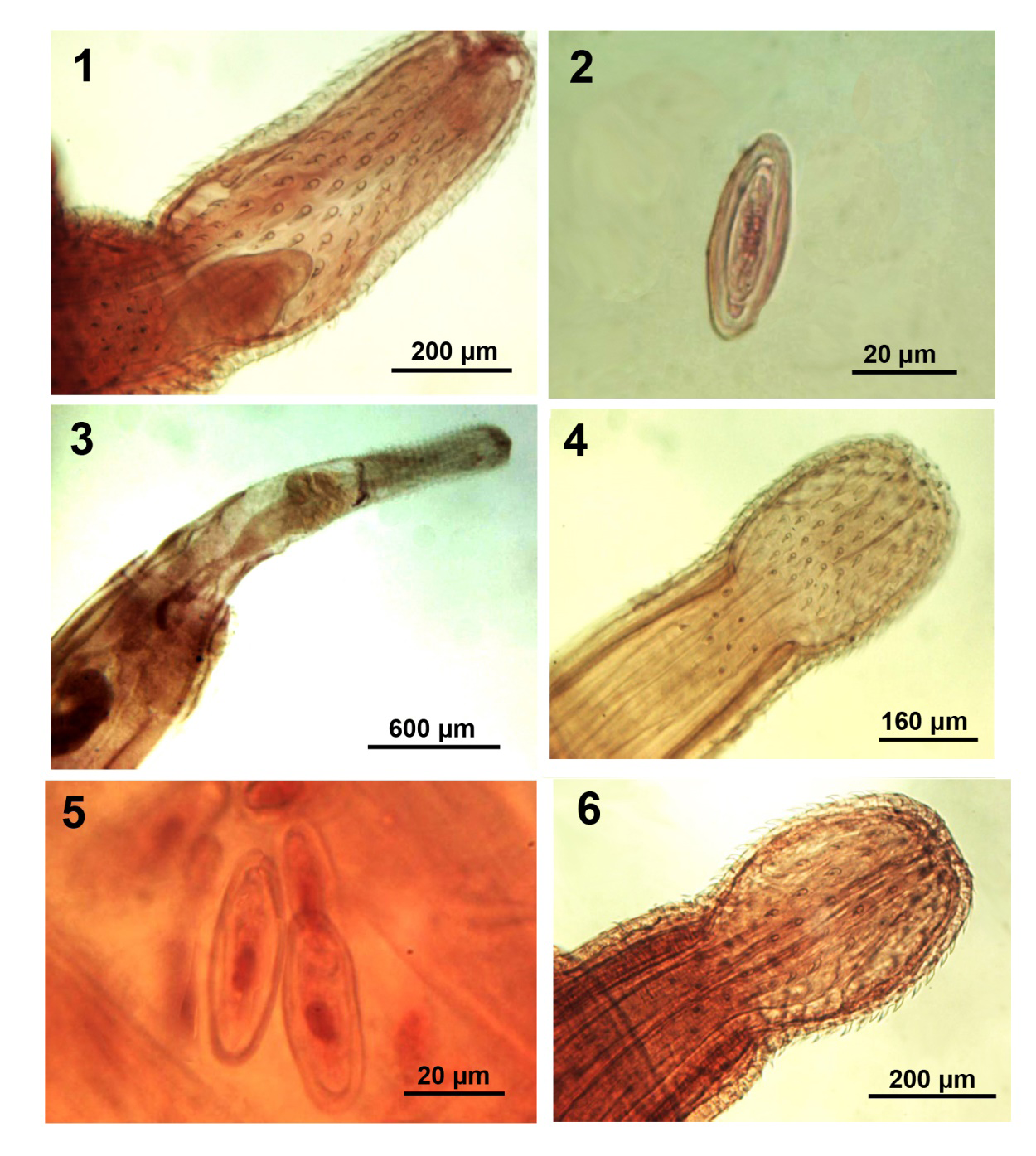
Figures 1-6: Morphology of polymorphid acanthocephalans from wildlife. 1, 2. Proboscis and egg of an adult Centrorhynchus spinosus from Strix aluco in Iran. Note the receptacle process (Fig. 1) 3. Anterior portion of a cystacanth of Centrorhynchus spinosus from Osteopilus septentrionalis in Florida. 4, 5. Proboscis and eggs of Sphaerirostris lancea from Vanellus vanellus in Ukraine. 6. Proboscis of Sphaerirostris picae from Pica pica in Bulgaria.
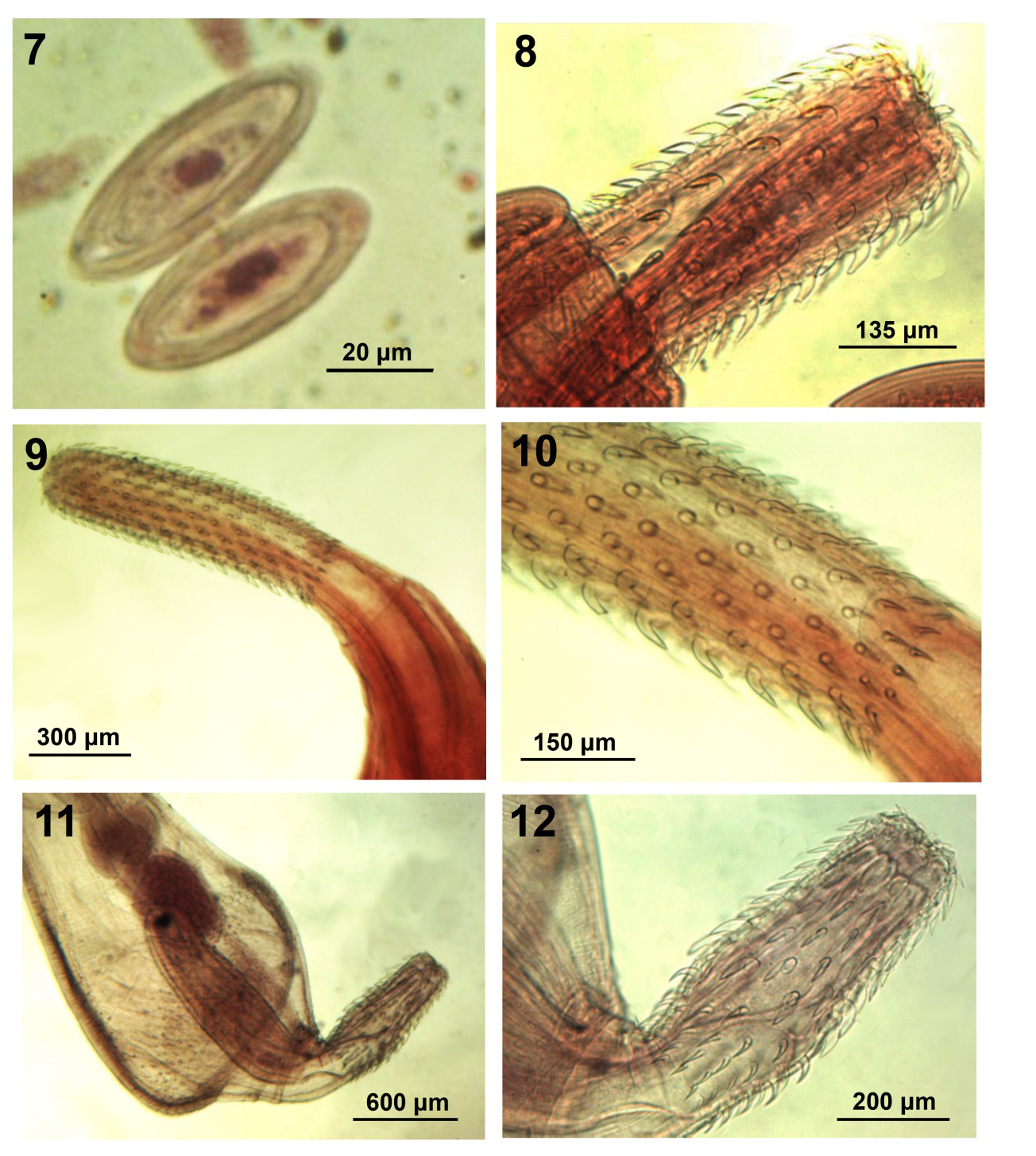
Figures 7-12: Morphology of polymorphid acanthocephalans from wildlife. 7. Eggs of Sphaerirostris picae. 8. Partially invaginated proboscis of Sphaerirostris picae. 9, 10. Proboscis of Plagiorhynchus (Prosthorhynchus) nicobarensis from Sturnus vulgaris in Iran. Note the basal hooks in perfect circle (Fig. 10). 11, 12. Anterior portion and proboscis of Andracantha gravida male from Gavia immer in Florida.
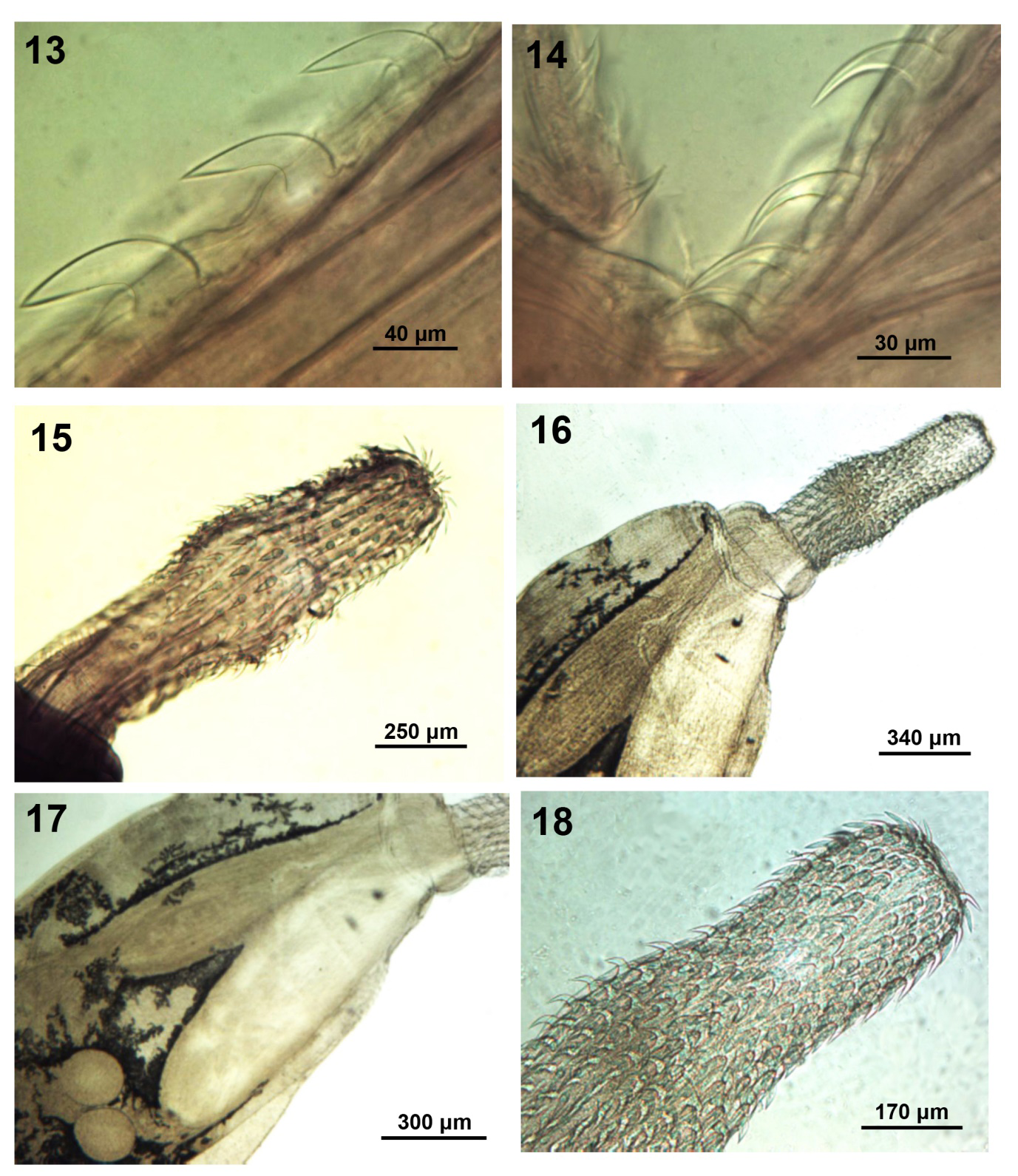
Figures 13-18: Morphology of polymorphid acanthocephalans from wildlife. 13, 14. Middle and posterior hooks and anterior trunk spines of the male specimen. 15. Proboscis of Arhythmorhynchus brevis from Casmerodius albus in Florida. 16-18. Anterior parts of a male Southwellina hispida from Ardea cineria in Macedonia showing the comparable shape and size of the lemnisci and the receptacle as well as the anterior half of the proboscis. Note the small round testes (Fig. 17).
- Specimens: Deposited at the Harold W. Manter Laboratory (HWML) Parasitology Collection no. 217063, Lincoln, Nebraska.
- Remarks: Compared to specimens described by above quoted observers, our specimens were only very mildly inflated at anterior trunk and at middle of proboscis and had a fewer spiniform hooks per row (8-9) in posterior proboscis. The number of the anterior rooted hooks was considerably more stable than that of the more variable posterior spiniform hooks. However, 8-10 spiniform hooks per row were counted in Khokholova’s [12] (Fig. 83b) specimens from Russia, the Volga region, Western Siberia, Azerbaijan, Kazakhstan, and Turkmenistan, even though she reported 10-15 such hooks per row. In addition, the testis appeared more rounded and separated in the Russian specimens of Khokhlova [12]
(Fig. 82c, p. 168). The proboscis of specimens from Louisiana [10] was extremely wide at middle; unlike our specimens or the Russian material. The eggs in our gravid female specimen were fusiform, 52-58 long by 20-25 in diameter, with blunt tips, concentric rings and minor polar prolongation (Fig. 2). Except for minor differences in measurements and the distant rounded testes, our specimens from Iran were morphologically most similar to those reported by Khokhlova [12] from Russia to Turkmenistan. Lisitsyna [13] reported Ukrainian specimens with a total of 23 hooks including transitional ones, and 8-10 spiniform hooks (Fig. 39a), testes very close to each other as in our specimens, but the lemnisci were very thick finger-like. In our specimens, the anatomy of the proboscis (Fig. 1) shows the receptacle process previously demonstrated in Sphaerirostris picae (Rudolphi, 1819) Golvan, 1956 by Amin, et al. [14].
Centrorhynchus spinosus (Kaiser, 1893) Van Cleave, 1924 cystacanths
Polymorphida (Centrorhynchidae) (Figure 3) We obtained 2 cystacanths of C. spinosus collected from a Cuban tree frog Osteopilus septentrionalis (Duméril & Bibron) in Tampa, Florida where the species was originally described. Nicole found the worms in the “small intestine area” of the body cavity in July, 2008, making the tree frog a paratenic host; a new record. Other species of frogs have been listed as paratenic hosts of various species of Centrorhynchus [15]. The proboscis armature of the two specimens fit that of the description of adults given above and keyed out to C. spinosus using the key of Richardson and Nickol [11] allowing for variations due to the different developmental stage.
The cystacanths (Fig. 3) had a bluntly pointed encased ovoid-shaped trunk, 1.42-1.47 mm long by 0.57-0.62 mm wide. Proboscis 1.06 mm long with 25-26 rows of 26 hooks each, divided by a mild constriction into 2 parts. Anterior proboscis 603-624 long by 218-260 wide, slightly inflated with 13 rooted hooks per row measuring 45-50 subapically; most robust hook just posterior to inflation. Posterior proboscis gradually widening basally, with 12-13 curved spiniform hooks per row measuring 35-37 posteriorly. Receptacle twisted and primordia of lemnisci visible. • Specimens: Deposited at the Harold W. Manter Laboratory (HWML) Parasitology Collection no. 217064, Lincoln, Nebraska.
Sphaerirostris lancea (Westrumb, 1821) Golvan, 1956
Polymorphida (Centrorhynchidae) (Figures 4, 5) Golvan [8] created Sphaerirostris as a subgenus of Centrorhynchus Lühe, 1911 with 21 species and several synonymies later increased to 26 species [16]. Twenty valid species are currently recognized [14]. Proboscis armature was a primary criterion distinguishing species apart. Sphaerirostris lancea is known from birds in the families Scolopacidae, Turdidae, Charadridae, Burhinidae, Recurvirostridae, Cuculidae in the Palearctic as well as in China, Taiwan, Egypt, and the USA [13, 17]. Lisitsyna provided us with 6 specimens (2 large females and 4 smaller males) from the lapwing Vanellus vanellus (Linn.) collected in Ukraine in 1988 [12]. Lisitsyna [13] provided detailed descriptions of S. lancea from Ukraine and Petrochenko [17] described it under Centrorhynchus from Russian Turkestan after Skryabin [18]. The brief description by Khokhlova [12] and her figure no. 86 are directly based on those provided by Petrochenko [17].
• General description: Trunk ovoid anteriorly and cylindrical posteriorly. The length proportion of the anterior ovoid to the posterior cylindrical trunk was 1:1.3-1.5 in our largest gravid females. This proportion was 2.3:1 in the specimens from Ukraine (Lisitsyna, [13]) (Fig. 1a) and the specimens from Russia (Petrochenko [17], Fig. 108). In our younger male specimens, the proportion was closer to 1:1. Cylindrical posterior trunk appearing to grow at a faster rate than the anterior trunk in maturing females. Proboscis in two parts; spheroid anteriorly and cylindrical posteriorly (Fig. 4). Anterior 7-8 hooks on anterior proboscis largest with prominent roots directed posteriorly followed by a transitional hook. Last 3 hooks smaller, slenderer with miniature anteriorly directed roots. Proboscis receptacle double- walled with spherical cephalic ganglion slightly posterior to its middle. Two slender, finger-like equal lemnisci not reaching anterior testis, attached posteriorly with filaments to body wall.
• Males: Based on 4 mature specimens with sperm. Trunk 5.87-11.00 mm long with anterior ovoid trunk 3.75- 6.75 (5.40) mm long by 1.12-1.87 (1.56) mm wide and posterior cylindrical trunk 2.12-4.37 (3.49) mm long by 0.42-0.82 (0.64) mm wide. Total proboscis 600-700 (640) long by 300-350 (330) wide anteriorly, with 11- 13 hooks per row. Proboscis receptacle 0.82-0.92 (0.88) long by 0.25-0.27 (0.26) wide. Lemnisci 1.50-1.57 mm long by 0.13 mm thick in one 11.00 mm long male. Testes in anterior trunk, ovoid, nearly contiguous. Anterior testis 0.82-1.05 (0.94) mm long by 0.42-0.50 (0.47) mm wide. Posterior testis 0.67-1.25 (1.00) mm long by 0.45- 0.50 (0.46) mm wide. Cement glands unequal, beginning at level of posterior testis, 2.10-4.25 (3.42) mm long by 0.12-0.17 (0.14) mm wide. Saefftigen’s pouch 0.75-1.30 (1.07) mm long by 0.25-0.50 (0.37) mm wide anteriorly.
• Females: Based on 2 large gravid specimens. Trunk 16.12-17.5 mm long with anterior ovoid trunk being 7.12-7.50 mm long by 2.35-2.40 mm wide and posterior cylindrical trunk being 9.00-10.00 mm long by 1.15- 1.37 mm wide. Total proboscis 603-832 long by 416 wide anteriorly, with 11-12 hooks per row. Proboscis receptacle 0.86-1.04 mm long by 0.28 wide. Genital pore subterminal and eggs ellipsoidal with concentric membranes (Fig. 5), 42-48 long by 17-22 wide.
• Specimens: Deposited at the Harold W. Manter Laboratory (HWML) Parasitology Collection no. 217065.
• Remarks: All measurements fell within the range recorded by Petrochenko [17] and Lisitsyna [13] except that the eggs were somewhat immature and smaller. In the specimens from Ukraine, the eggs appeared oblong with rounded ends (Lisitsyna [13], Fig. 4), the lemnisci were thick, the testes were round and distant, and the cement glands began at the level of anterior testis (Lisitsyna [13], Fig. 41a). In the specimens from Russia, the testes were ovoid and contiguous [17, 18], like ours.

Figures 19, 20: Morphology of polymorphid acanthocephalans from wildlife. Two eggs of Southwelliona hispida from Nycticorax violaceus in Florida showing the rounded polar prolongation of the fertilization membrane characteristic of this species.
Sphaerirostris picae (Rudolphi, 1819) Golvan, 1956
Polymorphida (Centrorhynchidae) (Figures 6, 7) Petrochenko [17] did not recognize Golvan’s [8] Sphaerirostris and did not include S. picae among his keyed out 51 species of Centrorhynchus. Khokhlova [12] did not include S. picae among her keyed out 11 species of Sphaerirostris and Yamaguti [4] recognized S. picae under Centrorhynchus and Lisitsyna [13] provided a detailed description of S. picae among 5 species of Sphaerirostris from Ukraine that she keyed out. Amin, et al. [14] recognized 20 valid species of Sphaerirostris, redescribed S. picae from Pica pica (Linn.) in Iran, discussed its seasonal distribution, and compared its intraspecific variability in populations from Iran, Bulgaria, Hungary, Ukraine, Armenia, France, and Russia. Amin, et al. [14] also described a special receptacle process at the anterior end of the proboscis receptacle extending into the proboscis. A similar structure was previously described in Corynosoma hamanni (Linstow, 1892) by [19, 20]. Radwan, et al. [21] studied the epidemiology of S. picae in Egypt.
In the present study, we evaluate the intraspecific variability in 4 additional populations of S. picae from Ukraine (provided by Olga Lisitsyna), Bulgaria (by Zlatka Dimitrova), Iran (by Ali Halajian), and Egypt (by Nahla Radwan) (Table 2) to compare with that in eight other populations reported earlier in Amin et al. [14].
| Males | Females | |||||||
|---|---|---|---|---|---|---|---|---|
| Population | Ukraine | Iran | Bulgaria | Egypt | Ukraine | Iran | Bulgaria | Egypt |
| Host | Pica pica, Corvus sp. | Corvus corone, C. frugilegus | Pica pica | Corvus corone | Pica pica, Corvus sp. | Corvus corone, C. frugilegus | Pica pica | Corvus corone |
| Sample size | 2 | 2 | 1 | 2 | 14 | 3 | 1 | 5 |
| Trunk length (mm) | 10.58- 12.50* | 9.00-10.75 | 9.75 | 11.87 | 11.00- 15.00 | 11.62-13.00 | 12.50 | 9.50- 14.75 |
| Trunk width (mm) | 2.00-3.00 | 1.50-2.10 | 1.82 | 2.15 | 0.92-3.17 | 1.12-2.50 | 1.75- 2.58 | |
| Ant. proboscis length X width | 390-500 X 375-420 | 426-550 X 343-400 | 425 X 350 | 500 X 400 | 475-550 X 375-425 | 500-575 X 325-447 | 500 X 400 | 500-575 X 425- 475 |
| Post. proboscis length X width | 300 X 450 | 158-312 X 260-374 | 200 X 350 | 300 X 375 | 175-250 X 300-400 | 150-333 X 325-437 | 175 X 350 | 2.75- 4.25 X 425-450 |
| Hook rows | 30-34 | 32-38 | 34 | 32-34 | 34-38 | 31-37 | 34 | 32-34 |
| Spiniform hook rows | 30-32 | 27-36 | 32 | 32-34 | 32-36 | 32-35 | 32 | 32-34 |
| Hooks/row | 8 | 8-10 | 10 | 9-10 | 9 | 8-10 | 8 | 9-10 |
| Spiniform hooks/row | 4 | 2-4 | 2-3 | 3-4 | 2-4 | 2-5 | 3 | 3-4 |
| Largest hook L | 57 | 55 | 43** | 50 | 52 | 58 | 50 | 50 |
| Largest spiniform hook L | 42 | 40 | 32 | 38 | 52 | 47 | 42 | 43 |
| Proboscis receptacle length X width (mm) | 1.17-1.62 X 0.35- 0.47 | 1.12-1.52 X 0.25-0.37 | 1.00 X 0.30 | 1.20 X 0.38 | 1.06-1.73 X 0.36- 0.45 | 1.30-1.62 X 0.26-0.35 | 1.15 X 0.37 | 1.12- 1.37 X 0.37- 0.40 |
| Lemnisci length (mm) | 2.37-2.40 | 0.78-2.92 | 2 | 2.5 | 2.27 | 1.25-2.60 | 2.57 | 1.75- 2.72 |
| Ant. testis length X width (mm) | 1.20-1.30 X 0.52- 0.75 | 1.37-1.80 X 0.67-0.95 | 1.17 X 0.67 | 1.25-1.62 X 0.62- 0.75 | XXX | XXX | XXX | XXX |
| Post. testis L X W | 1.12-1.25 X 0.95- 1.12 | 0.90-1.77 X 0.67-0.95 | 1.12 X 0.70 | 1.25-1.62 X 0.65- 0.70 | XXX | XXX | XXX | XXX |
| Cement gland L (mm) | 5.50-5.72 | 2.70-4.32 | 4.5 | 6.50-7.00 | XXX | XXX | XXX | XXX |
| Cement reservoir length X width (mm) | 1.00 X 0.37 | 0.83-1.90 X 0.52-0.75 | ___ | ___ | XXX | XXX | XXX | XXX |
| Female reproductive syst. length (mm) | XXX | XXX | XXX | XXX | 1.87-2.08 | 1.52-2.50 | ___ | 1.87- 2.24 |
| Egg length X diameter | XXX | XXX | XXX | XXX | 52-65 X 17-23 | 52-65 X 17-24 | 42-55 X 18-24 |
Table 2: ** Intraspecific variability among populations of _Sphaerirostris picae_ from various hosts in Ukraine, Iran, Bulgaria a
*All measurements are in micrometers except where otherwise noted. Extreme measurements are bolded. Table 2:** Intraspecific variability among populations of Sphaerirostris picae from various hosts in Ukraine, Iran, Bulgaria and Egypt.
- General Description: Based on specimens from all new 4 populations. Trunk fusiform, spheroid. Anterior proboscis with flat apical end and cylindrical-conical posterior proboscis with more widely spaced spiniform hooks and fewer hook rows (Fig. 6). No significant sexual differentiation in proboscis armature. Anterior hooks strong, 5th hook usually largest, with prominent roots followed posteriorly with transitional then smaller posterior hooks with anteriorly directed roots. Receptacle double-walled with oval cephalic ganglion near its center, lemnisci longer than receptacle and not reaching anterior testis.
- Males: Testes pre-equatorial, occasionally overlap. Four cement glands staggering anteriorly at level of posterior testis and joining posteriorly in 2 cement reservoirs. Subterminal gonopore.
- Females: Vagina with 2 sphincters, genital opening terminal in juveniles but subterminal in mature adults, and eggs oval with external sculpturing and concentric shells (Fig. 7).
- Specimens: Deposited at the Lincoln, Nebraska Harold W. Manter Laboratory (HWML) Parasitology Collections nos. 217066, 217067, 217068, 217069 from Ukraine, Bulgaria, Iran, & Egypt, respectively.
- Remarks: Table 2 provides a taxonomic comparison among the 4 populations of S. picae studied here. Specimens of the four populations appear similar with measurements falling within the same range of intraspecific variations, with minor exceptions marked by bolded font. Amin, et al. [14] noted that “confusion in the taxonomy of S. picae …. can be traced to intraspecific variability among various geographical populations and the choice of variable taxonomic criteria to distinguish between them.” A similar study was performed in 8 populations of another species of Acanthocephala, Mediorhynchus papillosus Van Cleave, 1916 from 11 host species in Europe, Taiwan, and the USA by Amin and Dailey [22] who noted that “Distinctiveness of geographical populations appears to be related to geographical restrictions, intermediate and definitive host specificity and distribution, and host feeding behavior.”
Plagiorhynchus (Prosthorhynchus) cylindraceus (Goeze, 1782) Schmidt & Kuntz, 1966
Polymorphida (Plagiorhynchidae) (Figure 8) Schmidt and Kuntz [23] synonymized Prosthorhynchus Kostylew, 1915 with Plagiorhynchus Lühe, 1911 and reduced the two genera to subgenera of the genus Plagiorhynchus s. lat. We have 3 semi-contracted adult females of (P.) (P.) cylindraceus that we have not previously reported. Two specimens collected from a common blackbird Turdus merula Linn. in Iran by Dr. Ali Halajian in 2011, and one specimen from a starling Sturnus vulgaris Rafinesque in Kenosha, Wisconsin by Bob Sewell in autumn, 1974. Available measurements include only proboscis size 1.07-1.14 mm long by 0.27 mm wide, largest proboscis hooks 83-94 long, number of hooks per row 13-14, and receptacle size 1.99-2.00 mm long by 0.32-0.36 mm wide.
- Remarks: The shape of the cylindrical proboscis and hooks (Fig. 8), and the above measurements are compatible with those described for P. (P.) cylindraceus from South Africa [24] or its synonyms Plagiorhynchus formosus (Van Cleave, 1918) Travassos, 1926 from North America and Plagiorhynchus taiwanensis Schmidt and Kuntz, 1966 from Taiwan or from Ukraine) [13], and Russia [12]. The contorted shape and limited measurements available in our 3 specimens from Iran and Wisconsin do not avail further detailed morphometric comparisons with specimens from other populations. Only Amin, et al. [24] provided detailed comparisons between P. (P.) cylindraceus and its synonyms P. (P.) formosus, and P. (P.) taiwanensis.
- Specimens: Deposited at the Lincoln, Nebraska Harold W. Manter Laboratory (HWML) Parasitology Collections nos. 217070, 217071 from Iran and Wisconsin, respectively.
Plagiorhynchus (Prosthorhynchus) nicobarensis (Soota & Kansal, 1970) Zafar & Farooqui, 1981
Polymorphida (Plagiorhynchidae) (Figures 9, 10) This acanthocephalan was originally described from 3 females collected from the ground thrush Zoothera citrina (Latham) at the Car Nicobar, India by Soota and Kansal [25] (Table 3). Ten years later, Zafar and Farooqui [26] redescribed it from an unknown number of males collected from warblers on mainland India. We are describing P. (P.) nicobarensis for the first time from the common starling in Iran; new host and locality records, and comparing its morphometrics with those in the original descriptions (Table 3).
- Description of our Iranian Specimens: Trunk elongated-fusiform. Proboscis cylindrical with hooks in parallel longitudinal rows with strong posteriorly directed roots except basal rootless spiniform hooks (Figs. 9, 10). Anterior hooks except apical largest. Basal hooks in perfect ring. Receptacle double-walled with cephalic ganglion near middle, lemnisci long, subequal, reaching posterior testis. Testes oval, contiguous, pre-equatorial, anterior testis occasionally totally overlapping proboscis receptacle. Cement glands 4, tubular, in 2 pairs. Saefftigen’s pouch prominent, pyriform, thick walled, overlapping posterior end of cement glands. Female reproductive system long, with subterminal gonopore, one sphincter and long straight uterine bell. Eggs elliptical with concentric shells.
- Specimens: Deposited at the Harold W. Manter Laboratory (HWML) Parasitology Collection no. 217072, Lincoln, Nebraska.
| Specimens | Original descriptions | Our material | ||
|---|---|---|---|---|
| Locality | Car Nicobar Island, Bay of Bengal | Aligarh, North Indian mainland | Iran | Iran |
| Reference | Soota & Kansal (1972) | Zafar & Farooqui (1981) | This paper | This paper |
| Host | Zoothera citrina (Latham), ground thrush | Turdoides caudatus (Dumont), common babbler | Sturnus vulgaris Linn., common starling | Sturnus vulgaris Linn. common starling |
| Worm sex | Females | Males | Females | Males |
| Sample size | 3 | “Few” | 7 | 6 |
| Trunk L X W (mm) | 7.00-8.50 X 1.10- 1.50* | 8.00-10.00 X 1.50-2.50 | 7.00-11.00 X 0.82- 1.40 | 5.50-6.57 X 0.70- 1.22 |
| Proboscis L X W (mm) | 1.10 X 0.30 | 0.85 X 0.20 | 0.97-1.25 X 0.27-0.32 | 0.92-1.12 X 0.25- 0.32 |
| Hook rows | 14-16** | 18-20 | 18-20 | 19-20 |
| Hooks/row | 14-15 | 16 | 14-15 | 14-15 |
| Largest hook | --- | 75 anteriorly | 73-85 anteriorly | 70-82 |
| Smallest basal hooks | Rootless | Ca. 40 | 57-62 | 55-62 |
| Receptacle L X W (mm) | 1.62 X 0.30 | 1.65 X --- | 1.67-2.20 X 0.30-0.42 | 1.50-2.37 X 0.32- 0.40 |
| Lemnisci L X W (mm) | Equal 2.00 X 0.10 | Subequal 2.65 & 2.75 X -- | 2.55-3.12 X 0.13 2.70-3.50 X 0.13 | 2.40-3.17 X 0.06 2.57-3.22 X 0.15 |
| Ant. testis length X width (mm) | XXX | 1.27 X 0.65 | XXX | 0.55-0.90 X 0.37- 0.67 |
| Post. testis length X width (mm) | XXX | 1.10 X 0.75 | XXX | 0.52-0.80 X 0.37- 0.55 |
| Cement gland L (mm) | XXX | 3.65 | XXX | 1.72-2.85 |
| Saefftigen’s pouch length X width (mm) | XXX | 1.50 X --- | XXX | 0.57-1.12 X 0.23- 0.34 |
| Bursa length | XXX | 300-360 | XXX | --- |
| Female reproductive syst. length (mm) | ___ | XXX | 1.46 | XXX |
| Egg length X width | 70 X 35 | XXX | 58-63 X 25-30 | XXX |
Table 3: ** Comparative morphometries between specimens in the original description of _Plagiorhynchus (Prosthorhynchus)_ _nicoba
*All measurements in micrometers unless otherwise noted. Extreme measurements are bolded. Table 3:** Comparative morphometries between specimens in the original description of Plagiorhynchus (Prosthorhynchus) nicobarensis from India and our specimens from Iran.
The morphology and organization of the Iranian specimens was similar to that of specimens in the original descriptions but showed variations. The measurements of all structures were comparable (Table 3). However, males from Iran had relatively smaller trunk and reproductive structures and females had slightly smaller eggs. Males from Aligarh had smaller basal hooks and females from Nicobar were reported with fewer proboscis hook rows. In general, Indian specimens had somewhat smaller receptacle. The above variations are considered to be of intraspecific nature in the various geographical populations. Updates to the key to species of Plagiorhynchus (Prosthorhynchus) in Amin, et al. [24] will need to take the above variations into account. The 3 definitive hosts (Table 3) are omnivorous and insectivorous [27, 28] and their invertebrate diet must include intermediate hosts commonly infected with larval acanthocephalans in India and Iran. For instance, invertebrates like the isopod Aramadillidium vulgare Latreille are naturally infected with plagiorhynchid acanthocephalans like P. cylindraceus that commonly infect starlings [29, 30].
Andracantha gravida (Algret, 1941) Schmidt, 1975
Polymorphida (Polymorphidae) (Figures 11-14) Schmidt [31] erected the genus Andracantha for 3 species of Corynosoma Lühe 1904 with two fields of trunk spines and genital spines that he redescribed. Andracantha gravida was originally described as Corynosoma gravida by Alegret [32] from the double crested cormorant Phalacrocorax auratus floridanus (Audubon) in Cuba. The description and measurements of the Cuban material fell within the range of those described by Schmidt [31] (with proboscis length correction) also from the brown pelican Pelecanus occidentalis carolinensis Gmelin from Florida and Louisiana.
We currently possess one adult male specimen (Fig. 11) collected by DJF from a common loon Gavia immer (Brunnich) in Franklin County, Florida on 28 June, 1983. This is a new host record for A. gravida. Our male specimen is similar to those described by Schmidt [31] but with some distinct variations. Our specimen was smaller, 2.62 mm long by 1.05 wide but with similar measurements of proboscis (675 by 250) (Fig. 12), receptacle (1.02 mm by 0.27 mm wide), cement glands (280 by 146), largest dorsal hook (73 by 31 wide at base), largest ventral hook (83 by 31 wide at base) (Fig. 13). Posterior hooks smallest compared to largest anterior trunk spines (Fig. 14). Equal length of anterior, trunk and genital spines (30-33) but somewhat smaller testes (400 by 275) (Fig. 11). The distribution of anterior and trunk spines was similar to Schmidt’s [31] specimens but our male specimen had only 1 genital spine as opposed to 18 marked in Schmidt’s [31] (Fig. 2).
• Specimens: Deposited at the Harold W. Manter Laboratory (HWML) Parasitology Collection no. 217073, Lincoln, Nebraska.
Arhythmorhynchus brevis Van Cleave, 1916
Polymorphida (Polymorphidae) (Figure 7) Schmidt [33] recognized two types of acanthocephalans in the former assemblage of Arhythmorhyynchus Lühe, 1911. “In one, the posterior trunk is very long and slender and the anterior trunk has one field of spines …(with) two or four cement glands. These are Arhythmorhynchus. In the second form, the anterior trunk has two fields of trunk spines in at least one sex, the posterior trunk is much shorter, and there are four cement glands. This group…. represents Southwellina” (Schmidt [33], p. 300). This species was originally described as A. brevis based on already prepared museum specimens by Van Cleave [5]. Travassos [34] assigned the species to genus Polymorphus and Meyer [35] accepted this generic assignment. Van Cleave [36] re-examined the status of the species from additional collections and re-assigned it back to Arhythmorhynchus based on his “belief that … the species is in agreement with the accepted concepts of the genus Arhythmorhynchus” and that Meyer’s [35] relegation “apparently” based on his interpretation of the proboscis as “either weakly ovoidal or possibly approaching cylindrical form” was in error (Fig. 15). He did not refer to the reason for the re-assignment by Travassos [34]. For more on the turbulent nomenclature history of A. brevis, see Amin [37].
MGS gave us two whole-mounted darkly stained specimens, a male and a female that were collected from a great egret Casmerodius albus (Linn.) in Florida. Exact date and locality were not noted but other collections from the same source were from Franklin County in June, 1983. Measurements of our specimens are in Table 4 compared to those from the descriptions of Van Cleave [5, 36].
• Specimens: Deposited at the Harold W. Manter Laboratory (HWML) Parasitology Collection no. 217074, Lincoln, Nebraska.
• Remarks: Van Cleave’s [5, 36] accounts were incomplete, not including measurements of lemnisci, receptacle, testes, cement glands, or cement gland ducts. Van Cleave did not count the number of cement glands but he (Van Cleave [36], Fig. 1) showed what appears to be 2 glands which is in agreement with Schmidt [33] of 2 or four glands in Arhythmorhynchus. However, our specimen appears to have more than 2 glands. Van Cleave’s [36] (Fig. 1) also appears to show a lemniscus reaching to anterior end of posterior testis as it does in our male specimen. We could not delineate the whole female reproductive system (nor did Van Cleave) but the gonopore was between subterminal to near terminal with a simple vagina adjoining an unremarkable sphincter. Van Cleave (2016) described eggs with rounded swellings at the polar ends of “middle” membrane. Our female specimen had only large elongate ovarian balls.
| Specimens | Our material | Material from Van Cleave (1916, 1945) in Japan | ||
|---|---|---|---|---|
| Locality | Florida, USA | Our specimens from Maryland, Minnesota, Illinois | ||
| Host | Casmerodius albus (Linn.), great egret | Botaurus lentiginosus (Rackett), American bittern Pandion haliaetus carolinensis (Linn.), osprey Ardia herodius herodius Linn., great blue heron Nycticorax nycticorax (Linn.), black crowned night heron | ||
| Worm sex | Female | Male | Females | Males |
| Sample size | 1 | 1 | 7 | 6 |
| Trunk L X W (mm) | 9.07 X 1.25 (ant.) & 0.62 (post.) | 6.12 X 1.15 (ant.) & 0.55 (post.) | 6.0-12.0 X 3.0 (ant,) & ca. 1.5 (post.) | 5.0-6.0 X 1.0-1.5 (ant.) & 0.5-0.75 (post.) |
| Ant. trunk spine L | 12-17 | 10-14 | 12 | 12 |
| Proboscis L X W at center | 738 X 302 | 634 X 239 | 665 X 340 | 665 X 340 |
| Hook rows | 15 | 15 | 13-16 | 13-16 |
| Hooks/row | 19 | 19 | 18 | 18 |
| Largest hook | 52 X 25 wide at base | 42 X 15 wide at base | Slightly larger ventrally, 41-47 X --- | |
| Posterior 7 hooks | Spiniform, rootless | Spiniform, rootless | Only 5 rootless posterior spiniform hooks (Figs. 2 & 3) | |
| Smallest basal hooks | 45 X 13 wide at base | 42 X 10 at base | 47 long | 47 long |
| Receptacle L X W (mm) | 1.67 X 0.26 | 1.17 X 0.22 | --- | --- |
| Lemnisci L X W (mm) | 1.92 X 0.31 | 1.70 X 0.25 Reaching post. testis | --- | Reaching post. testis (Fig. 1) |
| Ant. testis length X width | XXX | 600 X 275 | XXX | --- |
| Post. testis length X width (mm) | XXX | 525 X 300 | XXX | --- |
| No. cement glands | XXX | 2 or more | XXX | 2 (Fig. 1) |
| Cement gland length X width (mm) | XXX | 2.57 X 0.12 | XXX | --- |
| Cement gland ducts length X width | XXX | 884 X 93 | XXX | 0.57-1.12 X 0.23-0.34 |
| Egg length X width | Ovarian balls | XXX | 76-100 X 24-30 | XXX |
Table 4: ** Comparative morphometrics between specimens in the original description of _Arhythmorhynchus brevis_ from Japan and o
*All measurements in micrometers unless otherwise noted. Extreme measurements are bolded. Table 4:** Comparative morphometrics between specimens in the original description of Arhythmorhynchus brevis from Japan and our specimens from Florida.
Southwellina hispida (Van Cleave, 1925) Witenberg, 1932
Polymorphida (Polymorphidae) (Figures 16-20) Schmidt [33] resurrected the genus Southwellina Witenberg, 1932 and redefined it. Van Cleave [38] described S. hispida as Arhythmorhynchus hispidus from 4-5 mm long immatures in Illinois, USA and Fukui [39] redescribed it under the same name from 6 males and 13 females that infected Nycticorax n. nycticorax Linn. in Japan.
One of the synonyms of S. hispida is Polymorphus ardeae Belopolskaia [40] seven specimens of which were sent to us in February, 2008 for evaluation. Konstantinos Oyreilidis and Basil Ragias from Kavala, Greece sent 6 whole mounted males and 1 female collected from Ardea cinerea Linn. collected at the lagoons Vassova and Eratino of the River Nestos Delta in East Macedonia, Greece. They, among others [41] posted a report on these specimens as P. ardeae, the name under which their specimens were sent to us in 2008. Despite this universally accepted synonymy, it should be noted that, irrespective of measurements, these two synonyms differ markedly in proboscis shape (ovoid in P. ardeae but expanded medially in S. hispida) as well as the morphology of hook roots especially the posterior hooks having long anterior manubria in S. hispida (Amin, et al. [42], Fig. 2) and being without roots posteriorly and anteriorly in P. ardeae (see Belopolskaia [40], Fig. 1).
Only 2 male specimens from Macedonia, Greece were informative, one partially. The other specimens were totally corrupted with processing plaques. The anteriorly swollen trunk of our male specimens measured 3.37-8.75 mm long by 1.25-2.25 mm wide. Anterior trunk spines 22 long. Spindle-shaped proboscis 775-875 long by 325-350 wide at middle swelling (Figs. 16,18) with 24 rows of 14- 16 hooks each. Largest hook 50-57 long by 17-20 wide at base. Posterior hooks with prominent and straight anterior manubria. Receptacle 1.50-1.52 mm long by 0.42-0.45 mm wide. Lemnisci inflated, superficially similar to receptacle in shape (Figs. 16,17) and about as long, 1.50 long by 0.28-0.32 wide. Testes round, almost equal, post-equatorial, oblique to nearly side by side (Fig. 17), anterior testis 235 by 235, posterior testis 225 by 225. Bursa round, 550-650 long by 550-675 in diameter.
Comparatively, our specimens from Macedonia were somewhat smaller than those described by Fukui [39] from Japan in size of trunk and testes. Fukui [39], however, described the testes in his specimens as tandem or very slightly oblique (his Fig. 34) and the lemnisci were club- shaped and very long, about 2-3 times as large as the receptacle (his Fig. 27b). The receptacle and lemnisci in our specimens were about equally long (Fig. 17). The proboscis and receptacle were larger in our specimens and the armature was comparable. Van Cleave [38] described the species from 5 juveniles in Japan that are best described in detail by Amin, et al. [42] who described juveniles from California.
Our single gravid female specimen from Florida provided by DJF from the yellow-crowned night heron Nycticorax violaceus (Linn.) was larger in all common measurements (10.25 mm long by 3.25 wide) than the male specimens with anterior trunk spines 30 long, proboscis 936 long by 350 at middle swelling with 16 hooks per row, largest hook 63 long by 27 wide at base. The gravid female specimen from Florida was slightly smaller than the female from Macedonia but provided measurements of eggs of 80-95 by 22-28 that had the rounded polar prolongation of the fertilization membrane characteristic of this species (Figs. 19, 20). Other measurements and counts were comparable to those in Van Cleave [38] and Fukui [39]. • Specimens: Deposited at the Lincoln, Nebraska Harold W. Manter Laboratory (HWML) Parasitology Collections nos. 217075 and 217076 from Macedonia and Florida, respectively.
Acknowledgments
This project was partially supported by an ongoing institutional grant from the Parasitology Center, Inc., Scottsdale, Arizona, USA. We are immensely grateful to the many collaborators and former students who contributed the study materials that made the execution of this work possible. Students’ involvements were part of joint research projects mostly under independent study programs at the University of Wisconsin-Parkside, Kenosha Wisconsin. Collaborators simply shared research material as part of joint research understandings. We also gratefully recognize the help of Dr. Gabor Racz., Collection Manager, Harold W. Manter Laboratory of Parasitology, University of Nebraska- Lincoln for kindly accessing and cataloging our specimens.
References
-
Amin OM, Rubtsova NY (2023) New Annotated Records of Helminth Parasites, Mostly from North America, with Light Microscopy. I. Trematoda (Digenea), Cestoda, Nematoda. Int J Zoo Animal Biol 6(5): 000507.
-
Van Cleave HJ (1940) The Acanthocephala collected by the Allan Hancock Pacific Expedition, 1934, Allan Hancock Pacific Expeditions 2(15): 501-527.
-
Van Cleave HJ (1916a) Acanthocephala of the genera _Centrorhynchus_ and _Mediorhynchus_ (new genus) from North American birds, Transactions of the American Microscopical Society 35(4): 221-232.
-
Yamaguti S (1963) Systema Helminthum, Vol V Acanthocephala, New York and London, Interscience, pp: 423.
-
Van Cleave (1916b) A revision of the genus _Arhythmorhynchus_ with descriptions of two new species from North American birds. J Parasitol 2: 167-174.
-
Van Cleave HJ (1924) A critical study of the Acanthocephala described and identified by Joseph Leidy, Proceedings of the Academy of Natural Sciences, Philadelphia 76: 279-334.
-
Pujatti D (1950) _Centrorhynchus spinosus_ Kaiser, 1893 nel Sud India _Ospiti intermedi_ (Acanthocephala). Doriana 1(10): 1-4.
-
Golvan Y J (1956) Le genre _Centrorhynchus_, Lühe 1911 (Acanthocephala-Polymorphidae). Révision des espèces européennes et description d’une nouvelle espèce africaine, parasite de rapace diurne. Bull Inst Franc Afrique Noire A18(3): 732-791.
-
Schmidt GD, Neiland KA (1966) Helminth fauna of Nicaragua. III Some Acanthocephala of birds, including three new species of _Centrorhynchus_. J Parasitol 54(4): 739-745.
-
Nickol BB (1966) Acanthocephala of Louisiana birds. PhD Thes, Louisiana State University, pp: 118.
-
Richardson DJ, Nickol BB (1995) The Genus _Centrorhynchus_ (Acanthocephala) in North America with Description of _Centrorhynchus robustus_ n sp, redescription of _Centrorhynchus conspectus_, and a Key to Species. J Parasitol 81(5): 767-772.
-
Khokhlova IG (1986) Akantotsefaly nazemnykh pozvonochnykh fauny SSSR (Acanthocephala of terrestrial vertebrates of the USSR). Moscow USSR Nauka, pp: 275.
-
Lisitsyna OI (2019) Fauna of Ukraine. Acanthocephala. Publisher: NVP “Naukova Dumka Ukrainy” 31: 223.
-
Amin OM, Heckmann RA, Halajian A, Eslami A (2010) Redescription of _Sphaerirostris picae_ (Acanthocephala: Centrorhynchidae) from magpie, _Pica pica_, in northern Iran, with a special reference to unusual receptacle structures and notes on histopathology. J Parasitol 96(3): 561-568.
-
Schmidt GD (1985) Development and Life cycles. In: Crompton DWT, Nickol BB (Eds.), Biology of the Acanthocephala. Cambridge Univ Press, London, New York, USA, pp: 519.
-
Golvan YJ (1994) Nomenclature of the Acanthocephala. Research and Reviews in Parasitology 54(3): 135-205.
-
Petrochenko VI (1958) Acanthocephala of Domestic and Wild Animals, II, Moscow, Academy of Science USSR, pp: 478.
-
Skryabin KI (1913) Zur Acanthocephalen-Fauna Russisch Turkestans. Acanthocephalen der Sumpf- und Wasservögel, Zoologische Jahrbücher, Abteilung für Anatomie und Ontogenie der Tiere 35(4): 403-414.
-
Holloway HL Jr, Gee RJ (1990) Vesicles in the proboscis of Acanthocephala. J Parasitol 76(4): 585-587.
-
Nickol BB, Holloway HL (1968) Morphology of the presoma of _Corynosoma hamanni_ (Acanthocephala: Polymorphidae). J Morphol 124 (2): 217-226.
-
Radwan NA, Amin OM, Heckmann RA, Abd El Monsef MM (2012) An epidemiological study of _Sphaerirostris picae_ (Acanthocephala: Centrorhynchidae) from hooded crow (_Corvus corone cornix_) (Aves: Corvidae) in north Delta of Egypt. Sci Parasitol 13(2): 65-72.
-
Amin OM, Dailey MD (1998) Description of _Mediorhynchus_ _papillosus_ (Acanthocephala: Gigantorhynchidae) from a Colorado, USA population, with a discussion of morphology and geographical variability. J Helminthol Soc Wash 65(2): 189-200.
-
Schmidt GD, Kuntz RE (1966) New and little known plagiorhynchid Acanthocephala from Taiwan and Pescadores Islands. J Parasitol 52(3): 520-527.
-
Amin OM, Canaris AG, Kinsella JM (1999) A taxonomic reconsideration of the genus _Plagiorhynchus_ s lat (Acanthocephala: Plagiorhynchidae), with descriptions of South African _Plagiorhjynchus_ (_Prosthorhynchus_) _malayensis_, and a key to the species of the subgenus _Prosthorhynchus_. J Helminthol Soc Wash 66(2): 123-132.
-
Soota TD, Kansal KC (1972) The helminth fauna of Andaman and Nicobar. Acanthocephala. Rec Zool Surv India 66(1-4): 303-307.
-
Zafar MM, Farooqui HU (1981) Redescription of _Plagiorhynchus nicobarensis_ (Soota and Kansal, 1972) n comb, with comments on _Centrorhynchus spilornae_ Schmidt and Kuntz, 1969. Ind J Parasitol 5(2): 179-182.
-
Rana BD (1970) Winter food of the common babbler (_Turdoides caudatus_) in Rajastan. Ind Forest 96(2): 153- 155.
-
Feare C, Craig A (1999) Starlings and mynas Princeton, Princeton Univ Press pp: 288.
-
Moore J (1983) Responses of an avian predator and its isopod prey to an acanthocephalan parasite. Ecol 64(5): 1000-1015.
-
Nickol BB, Dappen GE (1982) _Armadillidium vulgare_ (Isopoda) as an intermediate host of _Plagiorhynchus_ _cylindraceus_ (Acanthocephala) and isopod response to infection. J Parasitol 68(4): 570-575.
-
Schmidt GD (1975) _Andracantha_, a new genus of Acanthocephala (Polymorphidae) from fish-eating birds, with descriptions of three species. J Parasitol 61(4): 615- 620.
-
Alegret M (1941) Contribución al conocimiento de los vermes parásitos de _Phalacrocorax auritus floridanus_. Molina y Co., Habana pp: 32.
-
Schmidt GD (1973) Resurrection of _Southwellina_ Witenberg, 1932, with a description of _Southwellina_ _dimorph_ sp. n., and a key to genera in Polymorphidae (Acanthocephala). J Parasitol 59(2): 299-305.
-
Travassos L (1926) Contribuições paro conhecimento da fauna helminthologica Brasileira. Revisãô dos acanthocephalos brasileiros. Parte II, Familia Echinorhynchidae Hamann, 1892, sub-fam. Centrorhynchinae Travassos, 1919. Mem Inst Oswaldo Cruz 19: 31-125.
-
Meyer A (1932) Acanthocephala. In Bronn HG (Ed.), Klassen und Ordnungen des Tier-Reichs, Akad Verlag MBH, Leipzig 4: 1-332.
-
Van Cleave HJ (1945) The status of the acanthocephalan genus _Arhythmorhynchus_, with particular reference to the validity of _A. brevis_. Trans Amer Micros Soc 64: 133- 137.
-
Amin OM (1992) Review of the genus _Polymorphus_ Lühe, 1911 (Acanthocephala: Polymorphidae) with the synonymization of _Hexaglandula_, Petrochenko, 1950, and _subcorynosoma_ Hoklova, 1967, and a key to the species. Qatar Univ Sci J 12: 115-123.
-
Van Cleave HJ (1925) Acanthocephala from Japan. Parasitol 17: 149-156.
-
Fukui T (1929) On some Acanthocephala found in Japan. Annot Zool Japon 12:255-270.
-
Belopolskaia MM (1958) The parasite fauna of birds of the Southern Nature Conservancy District (Far East). Acanthocephala. Parasitol Compend Zool Inst Akad Sci USSR 18: 403-420.
-
Oyreilidis K, Ragias B, Vatsos I, Fasseas K, Kolygas MN, Athanassopoulou F (2016) _Polymorphus ardeae_ Belopolskaja, 1958 (Acanthocephala) infection in sea bass (_Dicentrarchus labrax_ Linnaeus, 17858) in Greece. European Aquaculture Soc., Edinburgh, Scotland.
-
Amin OM, Chaudhary A, Singh HS (2022) Morphological and Molecular Description of Immature _Southwellina_ _hispida_ (Van Cleave, 1925) Witenberg, 1932 (Acanthocephala: Polymorphidae) from Cavity of the Paratenic Host _Gillichthys mirabilis_ Cooper (Gobiidae) in California, with Analyses of the Chemical Composition of Hooks and Spines. Acta Parasitol 67: 1107-1125.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind