Aggregation Role of Mandibular Gland Secretions in the Giant Tropical Ant Paraponera clavata (Formicidae: Paraponerinae)
Social insects exhibit sophisticated behavioral repertoires to maintain communication and regulate colony activities. To elucidate the behavioral mechanisms of social insects is essential a thorough understanding of morphophysiological characteristics, such as the functionality of exocrine gland secretions. In this study, it was analyzed whether the mandibular gland secretions influence the aggregation behavior of workers of the ant Paraponera clavata. Associated behavioral acts were also studied. Bioassays indicated that mandibular gland extract triggers aggregation behaviors in P. clavata workers. The observed behavioral acts include mandible opening, raising of the antennae, antennal contact with treated filter paper, selfgrooming, cleaning of the abdominal tip, and allogrooming. These observations demonstrate the potential multifunctionality of compounds secreted by mandibular glands within the behavioral repertoire of P. clavata.
Introduction
Ants are the most diverse group of eusocial Aculeata. There are substantial morphophysiological differences between mechanisms of social interactions of different ant societies. For instance, at least 85 types of exocrine glands have been described in ants; these glands produce compounds that are essential for chemical communication [1, 2, 3, 4]. Exocrine gland secretions are used for interaction between individuals, territory demarcation and defense, and also to recognition of sexual partners [5, 6].
Mandibular gland secretions are also highly diverse [7] and are known to trigger a series of behaviors (e.g., mandible opening, aggression) associated with colony defense. Pheromones produced by mandibular glands are the channel of chemical communication which trigger inter-individual recognition within the colony [1, 8] and induce specific surveillance and alert behaviors in workers [9, 10, 11]. The
response of workers to the pheromone is characterized by an initial alarm behavior, followed by aggregation to defend the colony [12, 13, 14]. Thus, pheromones produced by mandibular glands have two evident functions: (1) they serve as an alarm at high concentrations and (2) lead to recruitment of workers at low concentrations.
For the giant tropical ant, Paraponera clavata (Fabricius), few studies have investigated the role of exocrine glands, whose functionality is not yet fully understood [15, 16, 17]. The species P. clavata is the only living of the subfamily Paraponerinae. It is distributed in the Neotropical region, mainly in the tropical forests of western Brazil [18, 19] and in several other countries in South and Central America [20]. This extremely aggressive species is large (up to 2.5 cm in length) and well known for its painful sting. Nests of P. clavata are generally found at the base of trees [21]. The species is an opportunistic predator but can also exploit other resources, such as extrafloral nectars, acting mainly at night, although it can occasionally be seen foraging during the day. Recruitment of workers is achieved using pheromones for the foraging of both soil and tree canopy [22, 23, 24, 25]. Recently, Andrade, et al. [17] observed morphophysiological variations in the contents of P. clavata mandibular gland reservoirs, which contain solid, high-protein secretions. Such variations may indicate that mandibular gland secretions in P. clavata have a variety of functions in the communication between individuals.
In the present study, we test the hypothesis that mandibular gland secretions in P. clavata trigger behaviors associated with aggregation (include what type of behavior is associated). Ultimately, we aim to evaluate the behavioral role of mandibular secretions in P. clavata workers.
Material and Methods
Ants
Adult workers of P. clavata were manually collected from nests (N=3) located in the Inhamum Environmental Protection Area (04°53′S 43°24′W), Caxias, Maranhão, Brazil, and transferred to the Laboratory of Myrmecology (LAMIR) of the State University of Maranhão (UEMA), Brazil. Workers were maintained in plastic boxes lined with soil and substrate collected from the original nests. Artificial nests were kept under controlled conditions (27±2 °C and a photoperiod of 12 h light and 12 h darkness). Ants were fed a diet of apples, honey, and locust nymphs.
Preparation of Mandibular Gland Extract
Mandibular glands of P. clavata workers (N=20) were dissected in saline (125 mM NaCl). Then, extraction was performed by immersing and grinding the glands in 400 µL of hexane (98.5%). Extracts were used immediately after preparation. In all bioassays, workers had contact with extracts produced by individuals from their own colony.
Study of Aggregation Behaviors
Bioassays were performed to verify the effects of mandibular gland secretions on the aggregation behavior of P. clavata workers. For this, two parallel artificial trails measuring 60 cm in length and 3 cm in width were made in an acrylic arena. One trail was treated with mandibular gland extract and the other with hexane (control). Treatments were applied at predetermined points (10 µL per point) spaced 10 cm apart, as depicted in Figure 1. A release area (20 cm length × 6 cm width) at the base of the arena provided access to both trails, enabling ants to choose their preferred route. The side of treatment and control trails was randomly changed in each trial. At every trial, the arena was cleaned on both sides with hexane.
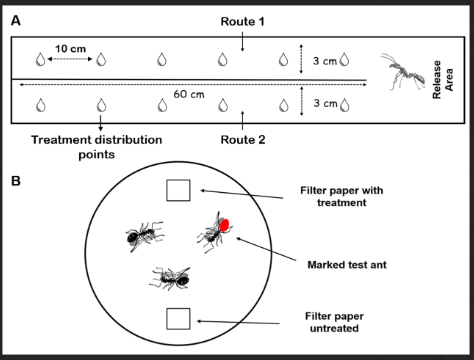
Ants were placed, one at a time, in the release area and observed for 5 min. or until they traveled to the end of one of the routes. Altogether, 30 workers from each nest (N = 3) were tested, totaling 90 bioassays. All ant movements were filmed with a digital camera (Sony Carl Zeiss, 14.1 megapixels).
Study of Behavioral Repertoires
Manipulative bioassays were performed under laboratory conditions to investigate the potential of mandibular gland secretions in triggering other behavioral repertoires in P. clavata workers. Groups of three nestmate workers were placed in arenas consisting of a plastic container (20 cm diameter × 10 cm height) lined with filter paper. Before placing the ants in the arena, one of the workers was marked with water-based paint (liquid, non-toxic) at the gaster level in order to monitor its behavior individually. The behavior of non-marked ants was not analyzed. Two pieces of filter paper (1 cm2) were introduced into the arena, of which one was untreated and one was treated with 10 µL of mandibular gland extract or hexane (control). Each test was started immediately after solvent volatilization (a few seconds). For each test combination (control and mandibular gland extract), 30 repetitions were performed per nest (N=3), totaling 180 bioassays. Bioassay analysis consisted in the observation of behavioral acts directed at treated and nontreated filter papers and nestmates. All ant movements were filmed with a digital camera (Sony Carl Zeiss, 14.1 megapixels).
Statistical Analysis
Data were analyzed in the R statistical software [26]. The effect of mandibular gland extract (x-variable) on the distance traveled by workers (y-variable) was assessed by a Generalized Linear Model (GLM). The effect of extract (x-variable) on ant behaviors (y-variable) was examined by a Generalized Linear Model (GLM) with Binomial distribution. The response variable was the proportion of a specific behavior in relation to the total number of behaviors observed. Analysis for each y-variable was conducted independently for each nest. Residual analysis was performed to assess the adequacy of error distribution and model fitting.
Results
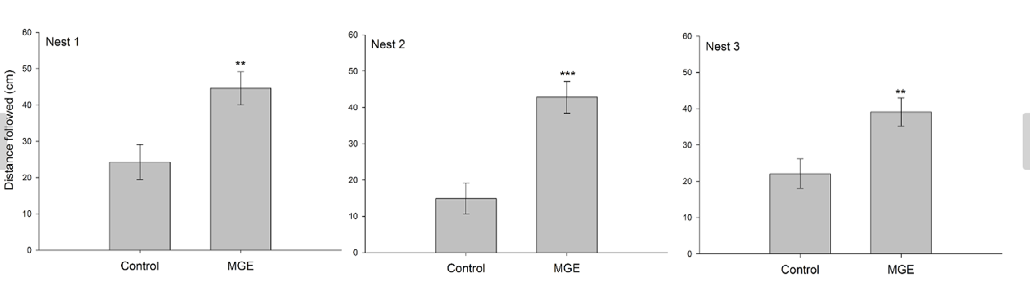
The distance followed by P. clavata workers on artificial trails was significantly affected by the presence of extract in all tested nests (GLM; nest 1: F1,28= 8.68, P= 0.004; nest 2: F1,28 = 17.05, P = 0.0001; nest 3: F1,28 = 8.55, P = 0.004). Ants who followed the trail containing mandibular gland extract traveled a greater distance than ants who followed the trail treated with solvent only (control) (Figure 2).
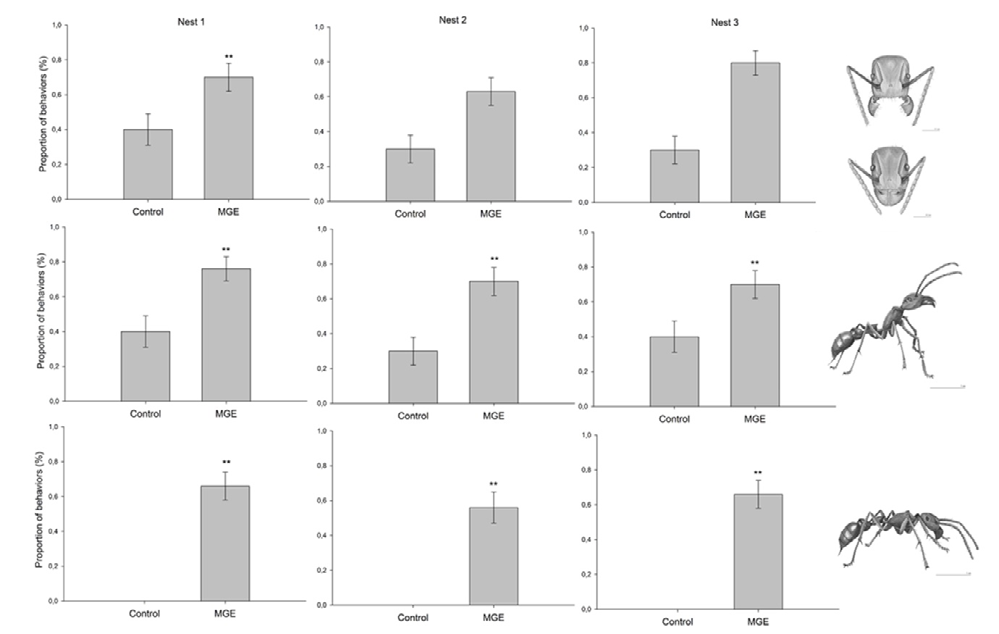
Six behaviors were frequently observed during the alarm response bioassay: mandible opening, raising of the antennae, antennal contact with filter paper, self-grooming, cleaning of the abdominal tip, and allogrooming. In all nests, the proportion of mandible opening, raising of the antennae, and antennal contact with filter paper was significantly higher in workers exposed to mandibular gland extract compared with control (Figure 3). The other behaviors (self- grooming, cleaning of the abdominal tip, and allogrooming) were not significantly affected by the presence of mandibular gland extract.
Discussion
In ants, the mandibular gland is predominantly associated with the production of defense pheromones [13, 27, 28]. Worker recruitment can be considered a secondary function of mandibular gland secretion, as evidenced by the results of bioassays with P. clavata. The mandibular gland extracts triggered aggregation behavior in workers.
Previous studies have shown that ant workers release a small amount of alarm pheromone, usually stored in mandibular glands. This immediately causes an increase in movement rate; nearby workers open their mandibles and then more workers are recruited to defend the colony, leading to aggregation [14, 29]. This behavioral repertoire may explain the results of the bioassay of the present study, in which P. clavata workers were more prone to follow the trail that contained mandibular gland extract than the path treated with only solvent (Figure 2).
In other ant species, pheromones responsible for aggregation behavior are generally secreted by abdominal glands, as in Linepithema humile Mayr [30, 31] and Camponotus pennsylvanicus De Geer [29]. However, in P. clavata, this effect seems to be produced by mandibular gland secretions.
According to Ghosh A, et al. [32], ants detect areas with the highest concentrations of aggregation pheromones and move in that direction. In the present study, we observed that P. clavata workers were in constant search for the origin of the odor, walking toward the path that contained the mandibular gland extract. This was possible because aggregation pheromones of ants are stable, unlike those of other insect groups, which are highly volatile [33]. As a result, the time span in which olfactory cues remain available and can be followed by ants is increased. Andrade TT, et al. [17] demonstrated that P. clavata individuals store mandibular secretions as protein-rich solids. However, for an effective alarm response, secretions generally need to be highly volatile. This characteristic explains the lack of aggressive alarm behavior in the test arena, such as sting exhibition, attack, and agitation. In bioassays conducted by Hermann, et al. [15] under natural conditions, mandibular gland extracts had no significant effects on alarm reactions of P. clavata workers.
The most common defensive responses of ants are mandible opening and sting exhibition [1, 27]. In the bioassays carried out in this study, P. clavata workers did not show their sting, but mandible opening was frequently observed when ants were exposed to mandibular gland extract. Mandible opening has been associated with the triggering of alarm behavior in several social hymenopterans [1, 14, 34, 35, 36].
Raising of the antennae and antennal contact with treated filter paper were also more frequent in ants exposed to mandibular gland extract than in those exposed to the control. These behaviors were reported in other ant species as an initial part of a series of defensive responses [35, 36]. However, only mandible opening was observed in the tested P. clavata workers as a defensive act; other behavioral patterns characteristic of an alarm response was not identified.
Self-grooming is one of the most common behaviors described in ethograms for ants exposed to different extracts [37, 38, 39, 40, 41]. In this study, although self-grooming and allogrooming were observed, these behaviors were not significantly altered by exposure to mandibular gland extract.
According to Napper, et al. [42], defensive behavior triggered by pheromones secreted from mandibular glands is a precursor of other pheromonal responses, such as alarm (high concentration) and aggregation (low concentration).
The responses of P. clavata workers observed here corroborate this affirmation.
Although behavioral patterns were not characteristic of an obvious alarm effect, mandibular gland extract showed potential to attract nestmates [43, 44, 45, 46, 47, 48, 49, 50].
In the present study, we demonstrated that mandibular gland extract triggers behaviors associated with aggregation in P. clavata workers. Other behaviors were also observed, indicating the importance and multifunctional potential of compounds secreted by mandibular glands in P. clavata [51, 52, 53, 54, 55, 56].
Acknowledgments
The authors would like to thank the members of the Laboratory of Myrmecology (LAMIR, Itabuna, Brazil), especially Paula Letycia Nunes de Souza, for their collaboration in the research. This study was partially funded by FAPEMA (Fundação de Amparo à Pesquisa e ao Desenvolvimento Científico e Tecnológico do Maranhão - Solicitation: INFRA-04004/15; UNIVERSAL-01633/16 e UNIVERSAL-00954/18) for financial support and Universidade Estadual do Maranhão, Centro de Estudos Superiores de Caxias (CESC) for logistic support. PFC and JHCD thank the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for the research fellowships provided (PQ 310395/2019-4 and PQ 304629/2018-9, respectively). TTA was supported by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) scholarship.
Author Contributions
Thito Thomston Andrade: Investigation, Formal analysis, Writing - Original draft. Paulo Fellipe Cristaldo: Formal analysis, Writing - Original draft - Review & Editing. Vinícius Albano Araújo: Formal analysis, Writing - Review & Editing. Jacques Hubert Charles Delabie: Conceptualization, Methodology, Writing - Review & Editing. Luiza Carla Barbosa Martins: Conceptualization, Methodology, Investigation, Writing - Review & Editing.
References
-
Caetano FH, Jaffé K, Zara FJ (2002) Ants: Biology and anatomy. UNESP pp: 131.
-
Adams RMM, Jones TH, Jeter AW, Licht HHF, Schultz TR, et al. (2012) A comparative study of exocrine gland chemistry in Trachymyrmex and Sericomyrmex fungus- growing ants. Biochemical Systematics and Ecology 40: 91-97.
-
Billen J, Sobotník J (2015) Insect exocrine glands. Arthropod Structure & Development 44(5): 399-400.
-
Guarda C, Lutinski JA (2020) Glandular secretions of ants (Hymenoptera: Formicidae): a review on extraction, chemical characterization and antibiotic potential. Sociobiology 67(1): 13-25.
-
Guerino AC, Landin CC (2003) Occurrence and morphology of integumentary glands in the abdomen of some bees (Hymenoptera: Apidae): a comparative study. Neotropical Entomology 32(2): 261-267.
-
Billen J (2008) The importance of exocrine glands in insect society. In: Vilela EF (Ed.), Social insects: biology in application. UFV, pp: 87-92.
-
Hölldobler B, Wilson EO (1990) The ants. Harvard University Press, pp: 732.
-
Gama V (1985) Salivary system of Camponotus (Myrmothrix) rufipes (Fabricius 1775), (Hymenoptera: Formicidae). Brazilian Journal of Biology 45: 317-359.
-
Fales HM, Blum MS, Crewe RM, Brand JM (1972) Alarm pheromones in the genus Manica derived from the mandibular gland. Journal of Insect Physiology 18(6): 1077-1088.
-
Cammaerts MC, Evershed RP, Morgan ED (1983) The volaite components of the mandibular gland secretion of the Myrmica lobicornis and Myrmica sulcinodis. Journal of Insect Physiology 29(8): 659-664.
-
Martins LCB, Delabie JHC, Serrao JE (2016) The function of intramandibular glands of the ant Neoponera villosa (Fabricius, 1804) (Hymenoptera: Ponerinae). Tropical Zoology 29(1): 10-15.
-
Moser JC, Brownlee RC, Silverstein R (1968) Alarm pheromones of the Atta texana. Journal Insect Physiology 14(4): 529-535.
-
Crewe RM, Blum MS (1972) Alarm pheromones of the Attini: their phylogenetic significance. Journal of Insect Physiology 18(1): 31-42.
-
Vilela EF, Lucia DTMC (1987) Insect pheromones: biology, chemistry and employment in Prague management. Viçosa-MG: University Press, pp: 155.
-
Hermann HR, Blum MS, Wheeler JW, Overal WL, Schmidt JO, et al. (1984) Comparative anatomy and chemistry of the venom apparatus and mandibular glands in Dinoponera grandis (Guérin) and Paraponera clavata (F.) (Hymenoptera: Formicidae: Ponerinae). Annals of the Entomological Society of America 77(3): 272-279.
-
Orivel J, Malherbe MC, Dejean A (2001) Relationships between pretarsus morphology and arboreal life in Ponerine ants of the genus Pachycondyla (Formicidae: Ponerinae). Annals of the Entomological Society of America 94(3): 449-456.
-
Andrade TT, Gonçalves WG, Serrão JE, Martins LCB (2019) Morphology of the mandibular gland of the ant Paraponera clavata (Hymenoptera: Paraponerinae). Microscopy Research and Technique 82(6): 941-948.
-
Penna ATM (2007) Subfamilia Paraponerinae. In: Jiménez E, et al. (Eds.), Systematics, biogeography and conservation of hunting ants of Colombia. Alexander Von Humboldt Biological Resources Research Institute, pp: 199-122.
-
Bacarro FB, Feitosa RM, Fernandez F, Fernandes IO, Izzo TJ, et al. (2015) Guide to the genres of formigas do Brasil. INPA pp: 388.
-
Kempf WW (1972) Abbreviated catalog of forms of the Neotropical Region (Hym, Formicidae). Entomological Study. 15: 3-344.
-
Moreira IJS, Cruz CDF, Fernandes AKC, Delabie JHC, Meneses CG, et al. (2020) Comparative study of the fauna of three large species of Brazilian mirmecofauna (Hymenoptera: Formicidae). Bulletin of the Paraense Museum Emílio Goeldi, Natural Sciences 15(2): 377-391.
-
Young AM, Hermann HR (1980) Notes on foraging of the giant tropical ant Paraponera clavata (Hymenoptera: Formicidae: Ponerinae). Journal of the Kansas Entomological Society 53(1): 35-55.
-
Janzen DH, Carroll CR (1983) Paraponera clavata. In: Janzen DJ (Ed.), Costa Rican natural history. Chicago: University of Chicago Press, pp: 752-753.
-
Breed MD, Bennett B (1985) Mass recruitment to nectar sources in Paraponera clavata: A field study. Insectes Sociaux 32: 198-208.
-
Fernandes IO, Souza JP, Baccaro FB (2015) State of the art on the phylogeny, taxonomy and biology of Paraponerinae. In: Delabie JHC (Ed.), the Poneromorphic Ants of Brazil. Phylogeny, pp: 43-53.
-
R Development Core Team (2019) R: A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing.
-
Nascimento RR, Sant’Ana AE (2001) Isolation and identification of semiochemicals of social insects. In: Vilela EF & Lucia DTMC (Eds.), Insect pheromones: biology, chemistry and employment in Prague management. Viçosa: Hollos pp: 65-71.
-
Choi MY, Meer RV (2015) Multiple functions of fire ant Solenopsis invicta mandibular gland products. Physiological Entomology 40: 196-204.
-
Ayre GL, Blum MS (1971) Attraction and alarm of ants (Camponotus spp.: Hymenoptera: Formicidae) by pheromones. Physiological Zoology 44(2): 77-83.
-
Cavill GWK, Robertson PL, Davies NW (1979) An Argentine ant aggregation factor. Experientia 35: 989- 990.
-
Cavill GWK, Davies NW, McDonald FJ (1980) Characterization of aggregation factors and associated compounds from the Argentine ant, Iridomyrmex humilis. Journal of Chemical Ecology 6(2): 371-384.
-
Ghosh A, Halder A, Kothari M, Ghosh S (2008) Aggregation pheromone density based data clustering. Information Sciences 178: 2816-2831.
-
Ali MF, Morgan ED (1990) Chemical communication in insect communities: a guide to insect pheromones with special emphasis on social insects. Biological Reviews 65(3): 227-247.
-
Brown WL, Eisner T, Whittaker RH (1970) Allomones and kairomones: transpecific chemical messenger. Bioscience 20(1): 21-22.
-
Lofqvist J (1976) Formic acid and saturated hydrocarbons as alarm pheromones for the ant Formica rufa. Journal of Insect Physiology 22(10): 1331-1346.
-
Marsaro Jr AL, Souza RC, Lucia DTMC, Fernandes JB, Silva MFGF, et al. (2004) Behavioral changes in workes of the leaf-cutting ant Atta sexdens rubropilosa induced by chemical components of Eucalipto maculata leaves. Journal of Chemical Ecology 30(9): 1771-1780.
-
Wilson EO (1976) A social ethogram of the neotropical arboreal ant Zacryptocerus varians (FR. Smith). Animal Behaviour 24: 354-363.
-
Brandão CRF (1983) Sequential ethograms along colony development of Odontomachus affinis Guerin (Hymenoptera, Formicidae, Ponerinae). Insectes Sociaux 30: 193-203.
-
Traniello JFA, Jayasuriya AK (1985) The biology of the primitive ant Aneuretus simoni (Emery) (Formicidae: Aneuretinae) II. The social ethogram and division of labor. Insectes sociaux 32: 375-388.
-
Claro DK, Santos JC, Durães ASJ (2002) Ethogram of the arboreal formiga Cephalotes pusillus (Flug, 1824) (Formicidae: Myrmicinae). Journal of Ethology 4: 31-40.
-
Santos JC, Yamamoto M, Oliveira FR, Claro DK (2005) Behavioral repertory of the weaver ant Camponotus (Myrmobrachys) senex (Hymenoptera: Formicidae). Sociobiology 46: 27-38.
-
Napper E, Pickett JA (2008) Alarm Pheromones of Insects. In: Capinera JL (Ed.), Encyclopedia of Entomology. Springer: Dordrecht pp: 85-95.
-
Bagnères AG, Morgan ED (1991) The postpharyngeal gland and the cuticle of Formicidae contain the same characteristic hydrocarbons. Experientia 47: 106-111.
-
Billen J, Delsinne T (2013) A novel intramandibular gland in the ant Tatuidris tatusia (Hymenoptera: Formicidae). Myrmecological News 19: 61-64.
-
Bolton B (2003) Synopsis and classification of Formicidae. Memoirs of the American Entomological Institute 71: 1-370.
-
Ginzel MD (2010) Olfactory Signals. In: Breed M, Moore J (Eds.), Encyclopedia of Animal Behaviour. Oxford: Academic Press, pp: 584-588.
-
Hölldobler B, Obermayer M, Plowes NJR, Fisher BL (2014) New exocrine glands in ants: the hypostomal gland and basitarsal gland in the genus Melissotarsus (Hymenoptera: Formicidae). Naturwissenschaften 101(7): 527-532.
-
Jandt J, Larson HK, Tellez P, McGlynn TP (2013) To drink or grasp? How bullet ants (Paraponera clavata) differentiate between sugars and proteins in liquids. Naturwissenschaften 100(12): 1109-1114.
-
Jarau S, Schulz CM, Hrncir M, Francke W, Zucchi R, et al. (2006) Hexyl decanoate, the first trail pheromone compound identified in a stingless bee, Trigona recursa. Journal of Chemical Ecology 32(7): 1555-1564.
-
Lattke JE (2003) Subfamily Ponerinae. In: Fernández F (Ed.), Introduction to ants of the neotropical region. Bogotá, Colombia: Alexander von Humboldt Biological Resources Research Institute, pp: 261-276.
-
Longino JT, Hanson P (1995) The ants. In: Hanson P & Gauld I (Eds.), The Hymenoptera of Costa Rica. New York: Oxford University Press, pp: 587-620.
-
McGee KM, Eaton W (2013) The effects of the conversion of a primary to a secondary tropical lowland forest on bullet ant (Paraponera clavata) foraging behavior in Costa Rica: A possible indicator of ecosystem condition. Journal of Insect Behavior 27: 206-216.
-
Morgan ED (2009) Trail pheromones of ants. Physiological entomology 34(1): 1-17.
-
Nieh JC, Contrera FAL, Neto NP (2003) Pulsed mass recruitment by a stingless bee, Trigona hyalinata. Royal Society of London B 270(1529): 2191-2196.
-
Pinheiro J, Bates D (2000) Mixed-effects models in S and S-plus. Springer-Verlag.
-
Stangler ES, Jarau S, Hrncir M, Zucchi R, Ayasse M (2009) Identification of trail pheromone compounds from the labial glands of the stingless bee Geotrigona mombuca. Chemoecol 19: 13-19.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind