Plasmodium's Secret: How a Complex Endomembrane System Drives Malaria's Deadly Efficiency
Plasmodium, the parasite responsible for malaria, has a highly specialized endomembrane system to support its life cycle. Beyond typical organelles like the nucleus, endoplasmic reticulum (ER), and Golgi apparatus, it features unique compartments such as micronemes, rhoptries, dense granules, and the parasitophorous vacuole, crucial for invading host cells and evading the immune system
Abbreviations
ER: Endoplasmic Reticulum; GRASP: Golgi Reassembly- Stacking Proteins; IMC: Internal Membrane Complex.
Editorial
Plasmodium, the parasite responsible for malaria, has a highly specialized endomembrane system to support its life cycle. Beyond typical organelles like the nucleus, endoplasmic reticulum (ER), and Golgi apparatus, it features unique compartments such as micronemes, rhoptries, dense granules, and the parasitophorous vacuole, crucial for invading host cells and evading the immune system. As the parasite progresses through its stages, it undergoes significant cellular changes, particularly in its ER and Golgi, to meet increasing metabolic demands and export proteins for host cell modification. Specialized organelles like rhoptries and micronemes facilitate host cell entry, while the actomyosin motor system drives its mobility. These adaptations allow Plasmodium to survive within red blood cells, making these mechanisms promising targets for antimalarial therapies to disrupt the parasite’s life cycle and reduce malaria transmission.
The endomembrane system in eukaryotic cells coordinates essential functions through a network of organelles, including the nucleus, ER, Golgi apparatus, lysosomes, vacuoles, vesicles, and endosomes. In Plasmodium, the parasite responsible for malaria, this system is highly specialized to adapt to its life cycle. Additional compartments- micronemes, rhoptries, dense granules, and the parasitophorous vacuole- aid in host cell invasion, survival, and immune evasion. The parasite’s apicoplast and mitochondria also support crucial metabolic functions. As Plasmodium transitions between mosquito and human hosts, these organelles undergo structural adaptations to meet changing metabolic needs, illustrating the tight link between cellular structure and parasitic adaptability (Figure 1) [1, 2, 3, 4].
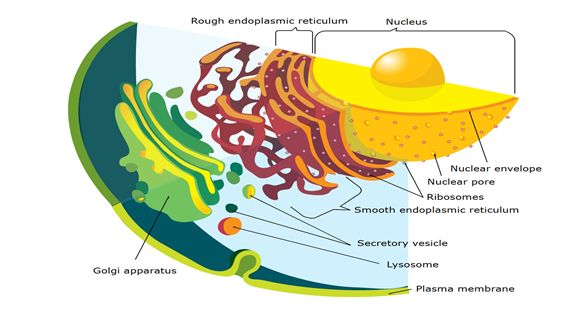
Figure 1: Endomembrane system on a Eukaryot cell. This image is licensed under creative commons attribution.
The parasite undergoes significant cellular changes throughout its life cycle to adapt to and survive within host erythrocytes. Initially, in the merozoite phase, its ER is underdeveloped, reflecting low protein synthesis needs as it focuses on invading host cells. As it progresses to the ring stage, the ER forms, supporting increased metabolic demands. By the trophozoite stage, the developed ER enables the export of proteins that remodel the host erythrocyte’s structure, enhance nutrient uptake, and help the parasite evade immune detection. These exported proteins use targeting sequences to reach specific destinations, ensuring effective host cell modification. The parasite’s nucleus, surrounded by a dual-layered nuclear envelope with ribosome-studded and fibrous membranes, uses nuclear pores to regulate molecular exchange and control gene expression. This gene regulation allows the parasite to adapt to changes within the host cell environment. Overall, the parasite’s ability to develop complex organelle structures, remodel host cells, and precisely control protein targeting and gene expression showcases its advanced strategies for survival and replication. These mechanisms highlight potential therapeutic targets for disrupting the parasite’s life cycle and reducing its impact on the host (Figure 2) [5, 6, 7, 8].
The Golgi apparatus in Plasmodium, the malaria parasite, is vital for its survival and ability to infect human hosts. It is essential in lipid synthesis, protein modifications, and sorting for the parasite’s development and red blood cell invasion. During its life cycle, Plasmodium relies on the Golgi to produce glycoproteins, especially in the early stages like the ring and trophozoite, but they are absent in the merozoite stage. The Golgi’s structure changes across stages, from tubular in the ring stage to vesicular in the early trophozoite stage and distinct compartments in the schizont stage. These three compartments (Cis, Intermediate, and Trans) receive process and sort proteins for delivery to key destinations, such as the plasma membrane or secretory organelles that help in red blood cell invasion [9, 10, 11].
Golgi reassembly-stacking proteins (GRASP) are crucial for maintaining the structure and compartmentalization of Plasmodium’s Golgi. These proteins, anchored to the membrane, ensure efficient protein processing. The Golgi also plays a vital role in secreting effector proteins that help the parasite manipulate host cells, evade immune responses, and create an environment conducive to replication, contributing to malaria severity. Given its importance in the parasite’s lifecycle, the Golgi is a promising target for antimalarial therapies. Disrupting its function or targeting GRASP proteins could impair the parasite’s ability to invade and replicate, offering a potential strategy for malaria treatment [12, 13].
Plasmodium merozoites possess a multi-layered pellicle composed of a plasma membrane and an internal membrane complex (IMC) linked to microtubules. This structure enables gliding mobility for invading red blood cells. The apical complex at the merozoite’s tip includes secretory organelles like rhoptries, micronemes, and dense granules. Rhoptries secrete enzymes that form a tight junction with the host cells, aiding entry. While micronemes provide adhesion molecules for attachment, and dense granules release proteins to modify the host cell post-invasion. The merozoite’s movement relies on the actomyosin motor system, where interactions between actin filaments and myosin generate the force needed for gliding mobility. This system is anchored to the IMC and powered by ATP, allowing the parasite to navigate host tissues. During the schizont stage, the Golgi apparatus processes vesicles to form rhoptries and micronemes. Rhoptries, characterized by a long neck and bulbous base, play a central role in altering host cell membranes to facilitate parasite entry. The coordinated function of structural elements, mobility mechanisms, and secretory systems enables Plasmodium to invade host cells. Understanding these mechanisms provides potential targets for antimalarial therapies to disrupt the parasite’s life cycle, thereby reducing malaria transmission and infection severity (Figure 3) [14, 15, 16, 17].
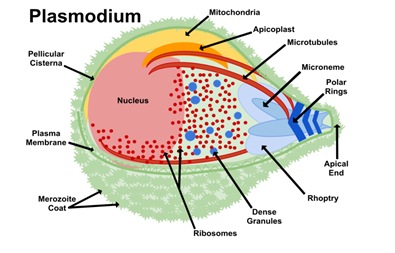
The specialized endomembrane system of Plasmodium supports its ability to adapt, survive, and thrive throughout its life cycle. By leveraging organelles like the ER, Golgi apparatus, apicoplast, and secretory compartments such as micronemes and rhoptries, the parasite efficiently invades host cells, evades immune defenses, and meets its metabolic needs. The dynamic structural adaptations of these organelles tailored to various environmental and host conditions underscore Plasmodium’s remarkable cellular complexity and resilience. Understanding these adaptative mechanisms deepens our knowledge of the parasite’s biology and helps us find new antimalarial therapies. Disrupting key processes like protein export, Golgi function, or host cell remodeling offers promising avenues for reducing malaria’s impact and combating drug resistance. As ongoing research unravels the intricate details of Plasmodium’s cellular machinery, these insights pave the way for innovative strategies to control and eradicate malaria [18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46].
Acknowledgments
The author acknowledges Mrs. Norri Zahra and Mr. Regragui Moumaris. The author thinks Prof. Nisen Abuaf (Sorbonne University and AP-HP) and Tech. Said Youssouf Chanfi (Sorbonne University). The author thinks Ing. Jean- Michel Bretagne (AP-HP). The author thinks Clr. Marie-Hélène Maës and Clr. Monique Abi (Research and Development Company).
References
-
Verhoef JMJ, Boshoven C, Evers F, Akkerman LJ, Gijsbrechts BCA, et al. (2024) Detailing organelle division and segregation in Plasmodium falciparum. J Cell Biol 223(12): e202406064.
-
Schroeder EA, Chirgwin ME, Derbyshire ER (2022) Plasmodium’s fight for survival: escaping elimination while acquiring nutrients. Trends Parasitol 38(7): 544- 557.
-
Wichers JS, Mesén-Ramírez P, Fuchs G, Yu-Strzelczyk J, Stäcker J, et al. (2022) PMRT1, a Plasmodium-Specific Parasite Plasma Membrane Transporter, Is Essential for Asexual and Sexual Blood Stage Development. mBio 13(2): e0062322.
-
Verhoef JMJ, Meissner M, Kooij TWA (2021)Organelle Dynamics in Apicomplexan Parasites. mBio 12(4): e0140921.
-
Anderson DC, Peterson MS, Lapp SA, Galinski MR (2024) Proteomes of plasmodium knowlesi early and late ring-stage parasites and infected host erythrocytes. J Proteomics 302: 105197.
-
Florentin A, Cobb DW, Kudyba HM, Muralidharan V (2020) Directing traffic: Chaperone-mediated protein transport in malaria parasites. Cell Microbiol 22(7): e13215.
-
Crabb BS, de Koning-Ward TF, Gilson PR (2010) Protein export in Plasmodium parasites: from the endoplasmic reticulum to the vacuolar export machine. Int J Parasitol 40(5): 509-513.
-
Van Dooren GG, Marti M, Tonkin CJ, Stimmler LM, Cowman AF, et al. (2005) Development of the endoplasmic reticulum, mitochondrion and apicoplast during the asexual life cycle of Plasmodium falciparum. Mol Microbiol 57(2): 405-419.
-
Gabriela M, Matthews KM, Boshoven C, Kouskousis B, Jonsdottir TK, et al. (2022) A revised mechanism for how Plasmodium falciparum recruits and exports proteins into its erythrocytic host cell. PLoS Pathog 18(2): e1009977.
-
Mayer DCG (2021) Protein Sorting in Plasmodium Falciparum. Life (Basel) 11(9): 937.
-
Struck NS, Herrmann S, Schmuck-Barkmann I, de Souza Dias S, Haase S, et al. (2008) Spatial dissection of the cis- and trans-Golgi compartments in the malaria parasite Plasmodium falciparum. Mol Microbiol 67(6): 1320- 1330.
-
Zhang X, Wang Y (2016) GRASPs in Golgi Structure and Function. Front Cell Dev Biol 3: 84.
-
Hallée S, Richard D (2015) Evidence that the Malaria Parasite Plasmodium falciparum Putative Rhoptry Protein 2 Localizes to the Golgi Apparatus throughout the Erythrocytic Cycle. PLoS One 10(9): e0138626.
-
Baum J, Richard D, Healer J, Rug M, Krnajski Z, et al. (2006) A conserved molecular motor drives cell invasion and gliding motility across malaria life cycle stages and other apicomplexan parasites. J Biol Chem 281(8): 5197- 5208.
-
Cowman AF, Tonkin CJ, Tham WH, Duraisingh MT (2017) The Molecular Basis of Erythrocyte Invasion by Malaria Parasites. Cell Host Microbe 22(2): 232-245.
-
Sibley LD (2004) Intracellular parasite invasion strategies. Science 304(5668): 248-453.
-
Kats LM, Black CG, Proellocks NI, Coppel RL (2006) Plasmodium rhoptries: how things went pear-shaped. Trends Parasitol 22(6): 269-276.
-
Moumaris M (2024) Malaria’s Hidden Weapon: How Plasmodium Transforms Red Blood Cells to Evade and Invade. Int J Zoo Animal Biol 7(6): 000629.
-
Moumaris M (2024) Advancements in Magnetic Resonance Imaging: Transforming Non-Invasive Diagnosis and Treatment Monitoring in Radiology. Journal of Life Sciences Research and Reviews. J Life Sci Res and Rev 2(5): 1-3.
-
Moumaris M (2024) Advancements in Diagnosing and Treatments Plasmodium knowlesi: Challenges and Innovations. Int J Zoo Animal Biol 7(5): 000622.
-
Moumaris M (2024) Confronting Plasmodium knowlesi: Challenges and Strategies in Malaria Healthcare. Int J Zoo Animal Biol 7(4): 000607.
-
Moumaris M (2024) Unraveling the Enigma: Tackling Knowlesi Malaria in Southeast Asia. Int J Zoo Animal Biol 7(2): 000585.
-
Moumaris M (2024) Unveiling the Enigmatic Plasmodium knowlesi: Insights, Challenges, and Promises in Malaria Research. Int J Zoo Animal Biol 7(1): 000566.
-
Moumaris M (2024) Unlocking the Potential: Overcoming Challenges in CAR-T Cell Therapy for Cancer Treatment. J Biotechnology and Bioprocessing 5(2): 2766-2314.
-
Moumaris M (2023) Revolutionizing Malaria Research: CRISPR unveils New Frontiers. J Biotechnology and Bioprocessing 4(5): 2766-2314.
-
Moumaris M (2024) Lyme Disease: A Zoonosis Tick- Borne Borrelia Bacterium [4/4]. Int J Zoo Animal Biol 7(1): 000549.
-
Moumaris M (2023) Lyme Disease: A Zoonosis Tick- Borne Borrelia Bacterium [3/4]. Int J Zoo Animal Biol 6(4): 000500.
-
Moumaris M (2023) Lyme Disease: A Zoonosis Tick- Borne Borrelia Bacterium [2/4]. Int J Zoo Animal Biol 6(2): 000465.
-
Moumaris M (2022) Lyme Disease: A Zoonosis Tick- Borne Borrelia Bacterium [1/4]. Int J Zoo Animal Biol 5(6): 000425.
-
Moumaris M, Bretagne JM, Abuaf N (2020) Nanomedical Devices and Cancer Theranostics. The Open Nanomedicine and Nanotechnology Journal 6: 1-11.
-
Moumaris M, Bretagne JM, Abuaf N (2019) Biological Membranes and Malaria-Parasites. The Open Parasitology Journal 7: 1-18.
-
Moumaris M, Bretagne JM, Abuaf N (2018) Hospital Engineering of Medical Devices in France. The Open Medical Devices Journal 6: 10-20.
-
Moumaris M, Rajoely B, Abuaf N (2015) Fluorescein Isothiocyanate-Dextran can track Apoptosis and Necrosis induced by heat shock of Peripheral Blood Mononuclear Cells and HeLa Cells. Open Biological Sciences Journal 1: 7-15.
-
Moumaris M, Rajoely B, Abuaf N (2012) The Naïve B Cells are the Lymphocytes with the Highest Anionic Phospholipid Binding Ratios. The Open Immunology Journal 5: 27-35.
-
Moumaris M (2007) Magnetic resonance imaging at the Hôtel-Dieu of Paris. Paris-Descartes University, Unités de formation et de recherche (UFR) biomédicale des Saints- Pères, France.
-
Moumaris M (2005) Identification of a new molecule to monitor apoptosis. Sorbonne-Paris-Nord University, France.
-
Moumaris M (2003) Biomedical research, the law of bioethics relating to the donation and use of elements and products of the human body. Paris-Descartes University, Faculté de Médecine, France.
-
Moumaris M, Abuaf N (2002) Use of labeled dextran for in-vitro assessment of increased cell permeability, cell death and apoptosis. Bulletin officiel de la propriété industrielle (Brevet n°00/09235) 2811682: A3.
-
Moumaris M, Benoliel S, Rouquette AM, Rajoely B, Abuaf N (2000) Phospholipid binding proteins on the plasma membrane of lymphocytes. J Autoimmun 15(2): 81-271.
-
Moumaris M, Ignoti S, Benoliel S, Oghina G, Rajoely B, et al. (1999) Characterization of B-cell adhering to the lamellar phospholipids. French Congress of Antiphospholipid Antibody, Paris, France.
-
Moumaris M (1996) Membranes érythrocytaires dans le paludisme: modèle d’étude: Souris- Plasmodium berghei anka. Université Pierre et Marie Curie, Faculté de Médecine, Paris, France.
-
Moumaris M, Sestier C, Miltgen F, Halbreitch A, Gentilini M, et al. (1995) Effect of Fatty Acid Treatment in Cerebral Malaria-Susceptible and Nonsusceptible Strains of Mice. The Journal of Parasitology 81(6): 997-999.
-
Sabolovic D, Moumaris M, Miltgen F, Sestier C, Halbreich A (1995) A subpopulation of red blood cells induced by bleeding or mosquito sucking. Chinese National Congress of Medical Biophysics, Shanghai, China.
-
Sabolovic D, Moumaris M, Miltgen F, Sestier C, Halbreich A (1995) Characterisation of subpopulation of red blood cells as a preferential target for malaria invasion. French Congress of Electrophoresis, Cell Electrophoresis, Pastor Institute, Paris 19(7): 1215-1219.
-
Moumaris M (1994) Effet des acides gras sur la malaria cérébrale chez des souris susceptible et non susceptible. Université Paris XII, Faculté de Médecine, Créteil, France.
-
Moumaris M (1992) Lyme disease: Serological study. University of Orleans, France.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind