Adipose Derived Mesenchymal Stem Cells Origin, Characteristics and Promises
Stem cell biology is currently one of the most leading areas of biomedical research, as enthusiasm for their application toward regenerative medicine continues to expand. Mesenchymal stem cells (MSCs), a well characterized population of fibroblastlike cells, exist in almost all tissues and represent a key cell source for tissue repair and regeneration. Adipose tissue derived mesenchymal stem cells (AT-MSCs) represent one of the most promising and advantageous populations of mesenchymal stem cells for use in cell-based therapy and tissue engineering. In the current mini-review we present the origin of AT-MSCs, their general and functional characteristics, and AT-MSCs-based preclinical and clinical approaches.
Introduction
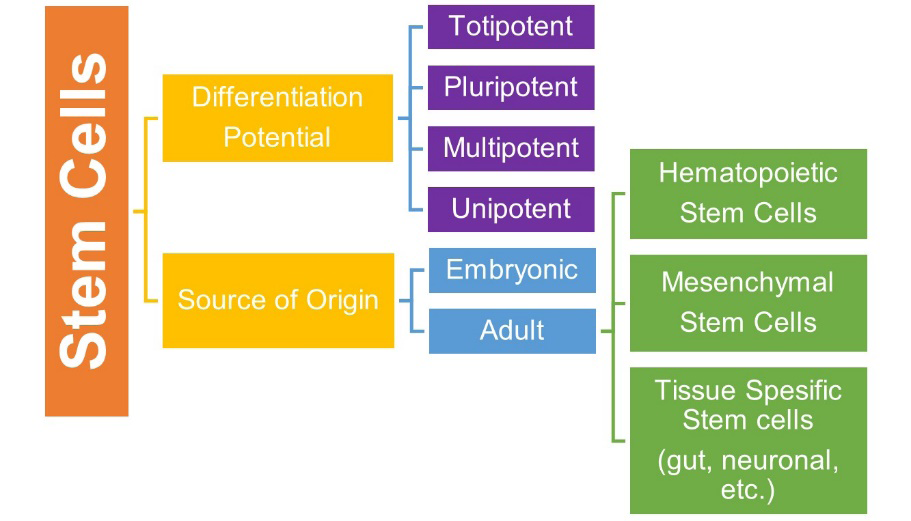
Stem cells are undifferentiated cells that can transform into specialized mature cells under various physical and experimental conditions and have the capacity for self- renewal [1]. Based on their differentiation capacity, stem cells are characterized as totipotent, pluripotent, multipotent, or unipotent. Another classification based on the source of origin categorizes the stem cells into two major groups:
embryonic and adult stem cells [2]. All three germ layers can be differentiated from embryonic stem cells produced from blastocyst cells. Even though they have a greater potential for tissue regeneration than adult stem cells, therapeutic use of such cells is not legally and ethically appropriate. However, adult stem cells reside in differentiated organs and maintain tissue homeostasis with their self-renewing, clonogenic, and multipotent properties [3]. Having many types of cell groups, adult stem cells contain hematopoietic stem cells (HSCs), MSCs, and other tissue-specific stem cells such as gut stem cells, neuronal stem cells, etc. is shown in (Figure 1). Among these cell types, MSCs have been an attractive option to develop clinical applications not only because of their multipotent properties, but also their angiogenic, neurogenic, and immunomodulatory features [4].
Mesenchymal Stem Cells
MSCs are the progenitors of connective tissues that have increasingly been used in regenerative medicine by reconstructing damaged mesenchymal tissues due to their potential to develop into diverse cell types like osteoblasts, adipocytes, chondrocytes, and neuroblasts [4, 5]. Mesenchymal stem cells are spindle shaped, and fibroblast- like adherent cells that proliferate into undifferentiated cells [6]. Although bone marrow stem cells are considered a significant source for mesenchymal stem cells, MSCs have also been derived from different tissues including placenta, umbilical cord blood, adipose tissue, dermis, and orofacial regions tissues and extensively evaluated in various research fields such as tissue repair, immune modulation [7, 8]. Numerous developmental and environmental signals mediate the balance of division of mesenchymal stem cells to generate the suitable number of stem and differentiated cells [9]. The proliferation capacity of differentiated cells is limited due to resulting in a shorter telomere after each division. However, since MSCs express telomerase enzyme, they have longer telomere than other somatic cells, and therefore their proliferation capacity is quite high [10].
According to the literature, there is a close relationship between stem cell differentiation’s direction and mitochondrial functions and dynamics. The level of mitochondria activity in MSCs is relatively low, following stimulation, mitochondrial biogenesis, such as protein levels of respiratory enzymes, oxygen consumption rate, mtDNA gene number, mRNA levels, and intracellular ATP content, increases. To benefit from regenerative medicine, it is important to have knowledge of the roles of mitochondrial dynamics such as energy metabolism and redox state related to MSC differentiation [7, 11]. In addition, when the metabolic activity conditions in MSCs are examined, while the energy requirement in MSCs is provided by glycolysis, differentiated cells are more dependent on oxidative metabolism [12, 13]. In the osteogenic differentiation process, with the activation of mitochondria in MSCs, ATP production is provided by oxidative phosphorylation [7, 14]. In both adipogenesis and osteogenic differentiation, oxygen consumption rate and respiratory enzyme activities goes up and mitochondrial membrane potential decreases significantly [7, 15]. In mammalian cells, oxygen is used in aerobic energy synthesis reactions as a substrate for cytochrome oxidase, the mitochondrial respiratory chain’s terminal enzyme [16]. Mitochondria, one of the main sources of O2 production, affect redox potential, ion balance and energy output of the cells. With the reduction of mitochondrial membrane potential, electron leaking from the electron transport chain increases the mitochondrial ROS production, which can generate super-radicals and cause tissue breakdown. Due to the oxidation of intracellular components, high amounts of ROS are considered to produce cellular dysfunction and destruction [17, 18].
Oxidative Stress
With the contribution of metabolic activities and environmental stressors, the body produces a variety of reactive species as free radicals or non-radicals. These compounds called pro-oxidants, which may be oxygen- derived or nitrogen-derived, are highly reactive and lead to cell and tissue damage by attacking macromolecules such as protein, lipids, and DNA [19]. In a healthy state, the human body has sufficient mechanisms to neutralize the effects of oxidant molecules through antioxidant systems including endogenous or exogenous antioxidant molecules and enzyme systems. However, if the balance between pro-oxidants and antioxidants shifts in favor of pro-oxidants, it causes the phenomenon called oxidative stress. This imbalance can develop due to either deficiency of antioxidants or accumulation of ROS [20]. Excessive ROS production in the body may change DNA structure, lead to protein modification and lipid peroxidation, as well as activate several transcription factors and produce pro/anti- inflammatory cytokines [21]. On that account, oxidative stress causes the initiation and progression of many diseases and pathological processes, including chronic inflammation, aging, carcinogenesis, cardiovascular diseases, diabetes, and neurological disorders [22]. Therefore, understanding the mechanisms involved in the regulation of oxidative stress is so critical.
Free Radicals and Their Neutralization
Free radicals, with an uneven number of electrons, are highly reactive species as they tend to bind with another electron to stabilize themselves. Although having a very short half-life, overproduction of these oxygen, nitrogen, or sulfur-derived species can result in damage to biomolecules and cells through a large chain of chemical reactions called oxidation [23].
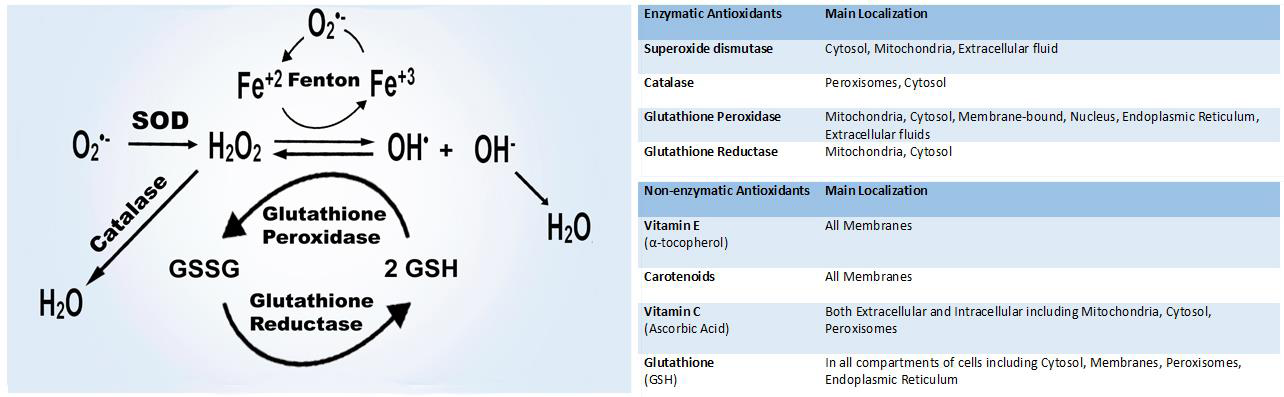
The main members of the ROS family are superoxide anion (02 -•), hydrogen peroxide (H2O2), hydroxyl radical (OH•), hypochlorous acid (HOCl), peroxyl radical (ROO•), and hydroperoxyl radical (HOO•) which are produced over the normal oxygen metabolism in the body, mainly in the mitochondria, both depending on enzymatic and nonenzymatic reactions [24]. Respiratory chain, phagocytosis, prostaglandin synthesis, and cytochrome P450 system are the main enzymatic processes resulting in ROS production [25]. In addition to mitochondrial respiration, nonenzymatic production occurs due to ionizing radiation, heavy metals, xenobiotics, and when oxygen reacts with organic compounds [26].
Including enzymatic and nonenzymatic antioxidant molecules, multiple mechanisms in the human body compose the antioxidant system to counteract the disruptive effects of ROS and oxidative stress [24, 27, 28]. Major compounds of the antioxidant system are shown in (Figure 2).
ROS and MSCs Differentiation
Due to the increasing potential applications of MSC in regenerative medicine, studies on MSC differentiation have shown that ROS can play an influential role in determining the direction of differentiation. In fact, undifferentiated cells are less resistant to exogenous ROS than differentiated cells [29]. While uncontrolled ROS can be harmful, a regulated basal level of ROS can perform as a signaling molecule to keep cell processes including proliferation, differentiation, and survival [17, 30]. According to Rodrigues et al., increased ROS stimulates the stress-induced MAPK pathways JNK, p38MAPK, and ERK, as well as apoptotic proteins and ant apoptotic pathways [31]. Up regulation of ROS has been reported to suppress osteogenic differentiation by reducing the activity of alkaline phosphatase (ALP) and Hedgehog (Hh) signaling pathway, which is fundamental for bone growth and maintenance [29, 32]. In fact, for osteogenic differentiation, the interaction of hormones (glucocorticoid and parathyroid hormones) and various extracellular signals such as wingless/patient1 ligands (Wnt), bone morphogenetic protein (BMP), fibroblast growth factor (FGF), transforming growth factor β (TGF-β), and Hh signaling pathway is constitutive [33, 34]. Chen, et al. reported a substantial drop in intracellular H2O2 and O2 levels on the second day following osteogenic induction, as well as a notable overexpression of the antioxidant enzymes SOD2 and Catalase after 14 days of induction. Furthermore, throughout osteogenic differentiation of hMSCs, the number of copies of mtDNA, protein levels of respiratory enzymes, oxygen consumption rate, mRNA levels of mitochondrial biogenesis genes, and intracellular ATP content increased, while intracellular ROS levels decreased dramatically, and antioxidant enzymes were up regulated. These findings imply that in osteogenic induction, mitochondrial respiratory activities increase in response to the growing energy demand [35]. As a result, it is suggested that Excess ROS suppresses osteoblastic differentiation and antioxidants possibly restore this process. Moreover, considering age-related bone loss and bone strength, it should be noted that elevation of intracellular ROS in the MSC of the elderly patient may result in decreased osteoblastic differentiation [36].
The generation of ROS in MSCs was found to be higher during adipogenic development. Differentiation of human MSCs into adipocytes begins with an elevation in mitochondrial oxygen consumption in addition to increasing of intracellular ROS generated from mitochondrial complex III due to mTORC1 signaling. The addition of exogenous H2O2 has been observed to stimulate adipogenesis in human adipose-derived MSCs depending on applied concentration.
In addition, in a study examining the stimulating role of endogenous nitric oxide (NO) in adipogenesis, there was a 50% elevation in basal NO levels in the first two days after adipogenic differentiation [37]. These findings showed that enhanced ROS generation is not just a result of adipocyte development [38, 39]. In another study, differentiation of human bone marrow-derived MSCs into adipocytes resulted in increases in SOD2, catalase, SOD3 mRNA and protein expression [40, 41]. Moreover, Yasunari et al. revealed that N-acetyl-cysteine (NAC) treatment reduced increased ROS formation, which in turn blocked adipogenic differentiation and mitochondrial targeted antioxidants can inhibit adipogenic differentiation [38]. Similar to adipogenic differentiation of MSCs, in chondrogenic differentiation, the level of ROS is elevated with an advanced decline in catalase activity [42]. ROS produced by NADPH oxidase 2 and 4 is required for differentiation of murine primary chondrocytes and the ATDC5 cell line. It was observed that Superoxide dismutase (SOD3) contributed for the reduction ROS in the extracellular matrix, whereas NAC inhibited chondrogenic differentiation. Consistent with this, SOD3 enzyme was diminished upon chondrogenesis [41, 43]. Furthermore, the addition of H2O2 increased chondrogenic differentiation, while administration of NAC inhibited it, implying that ROS is important in chondrogenesis [7, 29].
Because free radicals modulate the activity of transcription factors, cell proliferation, and certain levels of differentiation, studies on neurogenic differentiation have shown associations between low amounts of ROS and certain signaling pathways [44, 45]. The reduced level of ROS suppressed the Notch1 pathway and Hes1 expression, caused progressive elevation in the expression of Nestin, a neural stem cell-specific protein, and the antioxidant b-ME induced the differentiation of BMSCs in neural cells. Considering future MSC-based therapeutic approaches, these results suggest that ROS-based regulation of the Notch1 signaling pathway may be associated with neurogenic differentiation of MScs [46]. The overall regulatory role of ROS on stem cells is shown in Figure 3.
However, it has been shown that in addition to acting as second messengers in intracellular signaling that regulates cell functions including cell proliferation, differentiation, survival, and apoptosis, ROS can act as a antimicrobial agent when overproduced. There is strong evidence that excessive ROS production can contribute to cell and tissue damage as well as chronic inflammation, as they are involved in inflammatory events such as increased vascular permeability, leukocyte extravasation, phagocytosis, respiratory burst, and angiogenesis [47].
![Figure 3: However, it has been shown that in addition to acting as second messengers in intracellular signaling that regulates cell functions including cell proliferation, differentiation, survival, and apoptosis, ROS can act as a antimicrobial agent when overproduced. There is strong evidence that excessive ROS production can contribute to cell and tissue damage as well as chronic inflammation, as they are involved in inflammatory events such as increased vascular permeability, leukocyte extravasation, phagocytosis, respiratory burst, and angiogenesis [47].](/fulltextimages/8386/fig_3.png)
Inflammation
Inflammation is a protective immune response to invading pathogens, and it is operated by both the innate and adaptive immune systems. The acute inflammatory response with the events of systemic vasodilation, vascular permeability, and leukocyte emigration takes place when the innate immune system confronts the tissue damage by pathogens or stress [48]. Pathogen-associated molecular patterns (PAMPs) and danger-associated molecular patterns (DAMPs), which are released by injured cells due to trauma, ROS, bacterial inflammation, etc., can be recognized by the family of pattern-recognition receptors (PRRs). Macrophages, monocytes, dendritic cells, and neutrophils are among the immune cells that express PRRs [49]. As the NF-κB transcription factor is activated by stresses including oxidative stress, several cytokines and chemokines are released. Further, immune cells such as neutrophils recruit and migrate through the vascular wall into the infection site (Figure 4). Although this process is considered advantageous for the organism at the beginning, if it is not controlled, it can lead to excessive inflammation and many related inflammatory diseases [48]. The excessive generation of ROS and RNS can directly induce the secretion of inflammatory mediators and the stimulation of inflammation [49].
With the above-mentioned cellular processes which are interconnected especially with tissue regeneration, both ROS and inflammation take a significant regulatory role in signaling mechanisms and the metabolism of the stem cells.
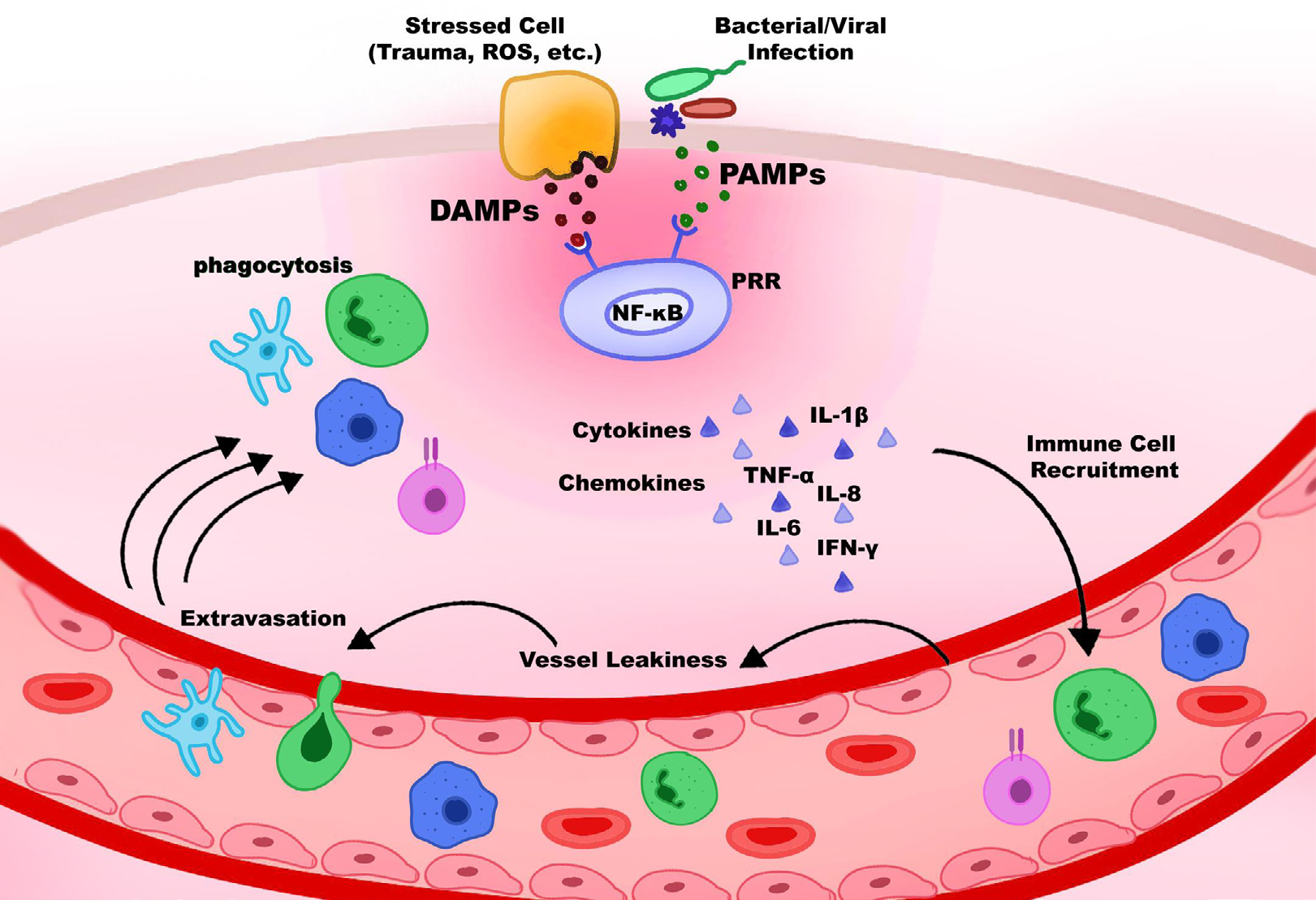
Inflammation and ROS Connection
Many authors have recognized the correlation between oxidative stress and inflammation. ROS can initiate intracellular signal transductions and mediate the stimulation of various transcription factors e.g., NF-қB [47]. These transcription factors in turn increase the expression of pro-inflammatory genes and induce the chronic inflammatory state [50]. At the same time, inflammatory cells promote oxidative stress by releasing multiple reactive species at sites of inflammation [51]. Most of the chronic disorders associated with high levels of ROS cause oxidative stress, which results in protein oxidation, glycosylated products, DNA damage, and lipid peroxidation [52]. It was reported that protein oxidations activate the release of signal molecules such as the pyridoxine 2 (PRDX2) molecules, which is considered an inflammatory signal. Once released, it functions as a redox-dependent inflammatory mediator that causes macrophages to secrete tumor necrosis factor-α (TNF-α) [53]. Macrophages play a key role in the regulation of inflammatory. In the inflammatory process, monocytes are recruited by neutrophils and move to the site of injury, where they transform into macrophages [54]. The link between macrophage population shift, also known as polarization, and tissue remodeling has been studied extensively. Based on their physiological properties, macrophages are classified as either pro-inflammatory conventionally activated (M1- like) or anti-inflammatory alternatively activated (M2- like). Once recruited and differentiated, macrophages are polarized to the M1-like state when stimulated by interferon- y (IFN-y), TNF-α, and Toll-like receptor-4 (TLR4) activation. Production of IL-10 and IL-4 causes the population to shift towards M2-polarization that signals tissue remodeling and repair [55, 56, 57].
The fact that macrophage blockade regulates Wnt recipient stem cell differentiation in intestinal crypts indicates a direct relationship between macrophages and stem cells [58]. MSCs activate macrophages toward an immunosuppressive M2 phenotype. As a result, MSCs create a more anti-inflammatory state by reducing TNF-𝛼, IFN-y secretion, and NK killer proliferation and increasing IL-10 and IL-4 secretion [56, 57, 59, 60]. Furthermore, MSCs secrete low levels of MHC class I and co-stimulatory molecules such as CD40, CD80, and CD86 to prevent all reactive antibody production and degradation [61, 62]. Because of their multimodal properties, MSCs are thought to have potential uses in different therapy modes such as new tissue formation, healing of tissue damage, grafting of other cells and tissues, and treatment of immune-based pathologies [63, 64].
The significance of ROS in macrophage-mediated immunity unquestionable. ROS has direct antibacterial effect against bacteria and parasites, in addition to redox control of immunological signaling and inflammatory activation [65]. Macrophages are frequently the first immune cells that come into contact with invading pathogens. Recognition of bacteria by macrophages results in the generation of ROS in many cellular compartments, which perform several antibacterial roles, including destruction of bacteria phagocytized by the oxidative burst produced by Nox2 [65, 66]. The most important point is at what stage ROS production tends to repair or destruction in tissue damage.
Dental Pulp Stem Cells
Among the MSCs, the ability of orofacial MSCs to self- renew and differentiate into many cell types has made them an ideal option for tissue regeneration. Since dental stem cells are ecto-mesenchymal origin, they can have both mesoderm and ectoderm properties and are easily accessible sources for multipotent postnatal stem cells [67]. Mesenchymal stem cells obtained from orofacial tissues are not only recommended for remodeling of hard tissue on account of their superior properties compared to bone marrow MSC cells but also suitable for regeneration of craniofacial tissues originating from the neural crest [10, 67]. Among the rich MSCs source identified in oral tissues are DPSCs, stem cells from human exfoliated deciduous teeth (SHED), periodontal ligament stem cells (PDLSCs), stem cells from apical papilla (SCAP), gingival fibroblastic stem cells (GFSCs), and dental follicle stem cells (DFPCs). These stem cell populations are obtained from various oral and maxillofacial tissues at different stages of development. DPSCs, for example, are isolated from the pulp tissues of permanent teeth, whereas SHED is isolated from the pulp of deciduous teeth. However, it has been noted that depending on the environmental conditions, there are multiple stem cell populations in the pulp tissue [68, 69].
Dental pulp, which includes both mesenchymal and ectodermal tissue and neural crest cells, is composed of blood vessels, nerve fibers, and connective tissue [70]. DPSCs originating from migrating neural crest cells are ectoderm- derived stem cells. Numerous In vitro and In vivo studies on cell growth, differentiation capacity, and potential to lead stem cell functions have revealed that DPSCs have impressive application potential for regenerative medicine [11, 71]. The multilineage differentiation potential of DPSCs is shown in Figure 5. Similar to that of MSCs, DSPCs exhibit a fibroblast- like morphology, adhesive quality to plastic surfaces, surface marker expression, proliferation, and colony forming behavior. In addition to mesenchymal stem cell markers such as CD73, CD90, and CD105, The immunophenotype of DPSCs has been found to include another mesenchymal stem cell marker, STRO-1, which is co-expressed with CD146 and pericyte antigen, [68, 72, 73] DPSCs also express neural lineage markers such nestin, B III tubulin, and glial fibrillary acidic protein (GFAP), which are seen on neural stem cells [74, 75].
![Figure 5: Similar to that of MSCs, DSPCs exhibit a fibroblast- like morphology, adhesive quality to plastic surfaces, surface marker expression, proliferation, and colony forming behavior. In addition to mesenchymal stem cell markers such as CD73, CD90, and CD105, The immunophenotype of DPSCs has been found to include another mesenchymal stem cell marker, STRO-1, which is co-expressed with CD146 and pericyte antigen, [68,72,73] DPSCs also express neural lineage markers such nestin, B III tubulin, and glial fibrillary acidic protein (GFAP), which are seen on neural stem cells [74,75].](/fulltextimages/8386/fig_5.png)
Gronthos, et al. published the first study on DPSCs, demonstrating that their stem cell capabilities are comparable to those of bone marrow stromal cells (BMSCs), with similar immunophenotype and calcified nodule formation under differentiation medium treatment [71]. DPSCs have shown a high differentiation ability, such as neurogenesis, adipogenesis, osteogenesis, chondrogenesis, angiogenesis, and dentinogenesis, and are tightly controlled by a variety of growth factors, including tumor growth factor-β (TGF-β), basic fibroblast growth factor (FGF), platelet-derived growth factor (PDGF), epidermal growth factor (EGF), TNF-, and insulin-like growth factors (IGF) I and II [76].
ROS and Dental Pulp Stem Cell Differentiation
According to the literature, as in DPSCs, mitochondrial functions and dynamics determine the pathway of stem cell development [11]. In cardiomyocyte coculture, DPSCs showed greater mitochondrial respiratory capacities and lower mtROS than BM-MSCs and AD-MSCs. In addition, depletion of mitofilin has induced osteogenic/dentinogenic differentiation in DPSCs. Mitofilin regulates osteogenesis and is a transmembrane protein found in the inner mitochondrial membrane [77]. It was demonstrated that odontogenic differentiation in DSPCs, initiated by mitochondrial elongation with developed crista, increasing mitochondrial oxygen consumption rate, ATP production, and upregulation of mitochondrial glycolytic enzyme activity [78]. Long-term H2O2 treatment of DPSCs increased ROS production, which reduced cell survival and overexpression of antioxidant molecules Cu/Zn and Mn SOD as well as odontogenic/ osteogenic markers like dentin sialophosphoprotein, dentin matrix protein-1, osteopontin, bone sialoprotein, Runx-2, and bone morphogenetic protein 2 and 7. However, PPAR- overexpressed cells enhanced dentin mineralization despite being exposed to oxidative stress [79]. In a recent study, a high O2 environment compared to physiological oxygen levels activated p38 / p21 phosphorylation and /NRF-2 signaling pathway in DPSCS and decreased DPSC proliferation due to increased oxidative stress [80]. Moreover, SHED differentiated into neuronal cells with higher mitochondrial membrane potential and mt DNA in addition to longer mitochondria [81].
However, it was revealed that DPSCs are heterogeneously located in different niches in dental pulp tissues including subodontoblast layer, pulpal vasculature, and central pulp. Therefore, their proliferative capacity may be different [82]. Alaidaroos, et al. showed that when DPSC has a high proliferative ability, they resist H2O2-induced senescence and increase SOD2/glutathione S-transferase Z1 (GSTZ1) expression and SOD activities [83]. Whereas, low proliferative subpopulations exhibited accelerated senescence with low SOD, catalase and glutathione-associated antioxidant activities. Thus, this increases the complexity of individual DPSC subpopulations regarding their origin and regenerative properties and may cause significant limitations in mesenchymal stem cell (MSC)-based therapies.
Inflammation, Regeneration and Dental Pulp
DPSCs have been proven in numerous studies to develop into odontoblast-like cells and create a dentin/ pulp-like complex [84, 85, 86]. To form new pulp-like tissue, two tissue engineering approach can be applied in the root canal, the first is cell transplantation and the second is cell homing. Based on cell transplantation technique, stem cells together with growth factors such as FGF, PDGF, TGF-β, IGF, vascular endothelial growth factor (VEGF), nerve growth factor (NGF) and bone morphogenetic proteins (BMPs) are placed on a proper scaffold and applied directly to the root canal. In the cell homing approach, recombinant signaling molecules or endogenous, dentin-derived growth factors attract the local stem cells to migrate from periapical tissues into the root canal. This approach can also be performed to repair damaged dentin-pulp complexes caused by local inflammatory disorders [87, 88, 89].
Dental inflammation has been recognized as an adverse factor leading to pulp destruction, mostly through necrosis or apoptosis. If dental pulp inflammation is in a reversible phase, to generate a reparative dentine, stem precursors and pericytes can be mobilized for odontoblastic differentiation [90, 91, 92]. Wnt receiving stem cell activation that can differentiate into odontoblast-like cells requires the presence of macrophages in the dental pulp. It was reported that elevation of Wnt cells significantly stimulated the up regulation of TGF-β1 in the dental pulp and produced acceleration in the polarization of the Wnt/β-catenin pathway from the pro-inflammatory phase to the anti-inflammatory phase (M1-M2 polarization) via GSK-3 antagonist small molecules, thus enhanced the reparative capacity [93, 94, 95]. Moreover, Neves, et al. showed that when macrophages are depleted, they significantly impair the reparative dentin- forming capacity, causing an accumulation of neutrophils at the injury site that leads to excessive inflammation. On the other hand, depletion of neutrophils suppressed inflammation and increased reparative dentin secretion in the pulp [96]. Regulation of inflammation with macrophages in dental pulp stem cells is shown in (Figure 6).
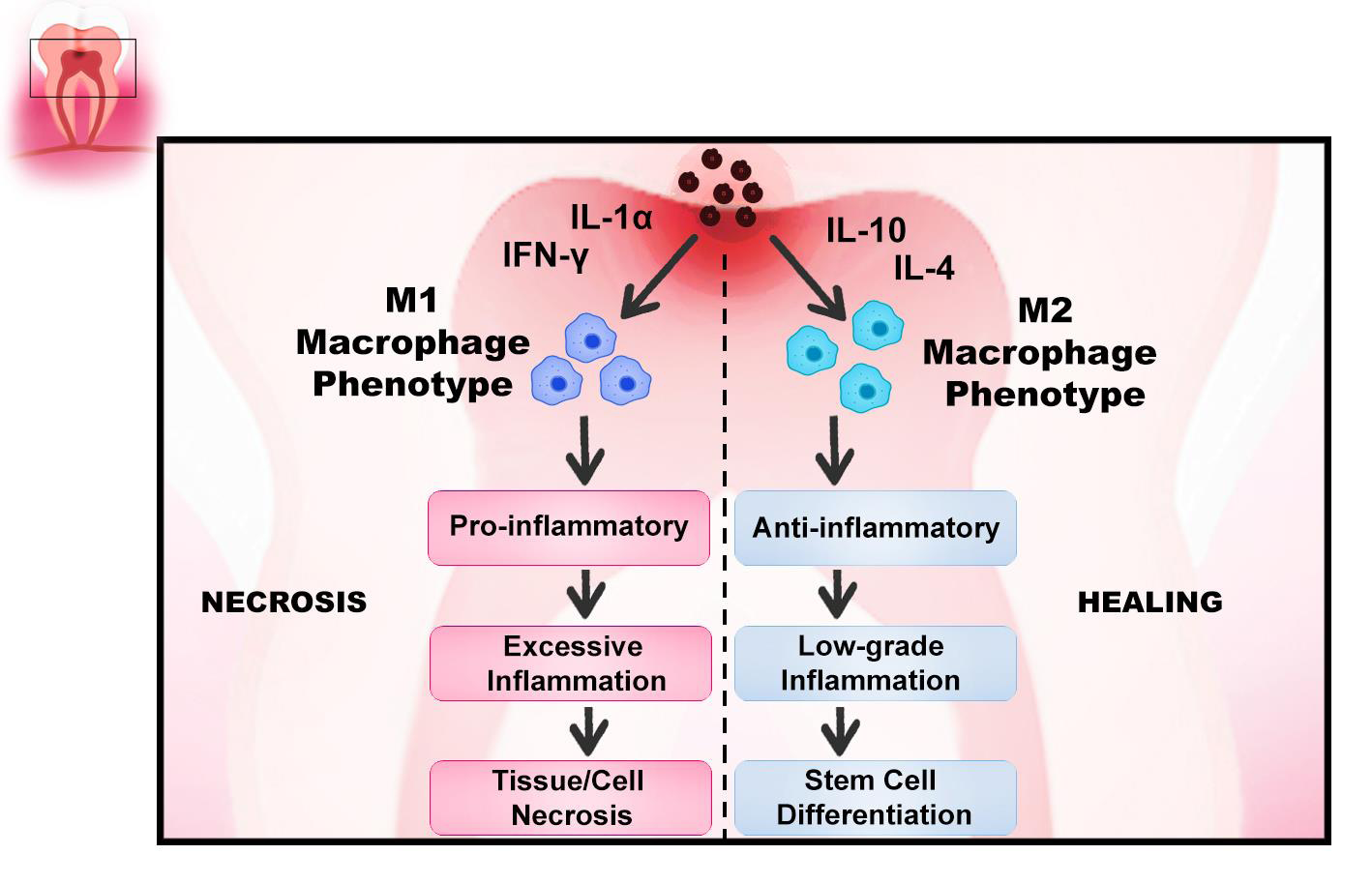
However, the development of reparative dentin is a complex mechanism that needs a mild inflammatory reaction within the dental pulp [92, 96, 97]. Odontoblasts, immune-competent cells (monocytes, macrophages, and stem/progenitor cells), and subsequently pulp fibroblasts generate a large number of signaling molecules to regulate the immune response following recognition of the pathogen. Cells slide along the root and migrate to the wound side, towards the crown. Inflammatory processes cause the proliferation of pulp progenitor stem cells and increase in number and size. Their phenotype changes and they become odontoblast-like cells that produce collagen, ALPase, and SPARC/osteonectin [90].
Considering pulp inflammation caused by deep caries, it is well recognized that bacterial lipid byproducts increase intercellular ROS, as well as inhibition of cell growth, cell cycle kinetics, and protein synthesis [90]. The control of the cytotoxicity of generated intercellular ROS, in turn, can stimulate the recruitment of certain pulp cells such as progenitor stem cells, involved in reparative and regenerative processes [98, 99]. Since glutathione effectively removes the excessive ROS, in a recent study, MTA agent was used with soluble N-acetyl cysteine antioxidant molecule and enhanced the differentiation capacity of DPSCs [100].
Therefore, in the design of reparative dentin formation and treatment models in regenerative medicine, it is important to know the inflammatory processes that occur in the tooth pulp as a result of dentin injury, as well as their impact on local stem cell activation [90, 96]. On the other hand, it has been shown that ROS accumulation leads to premature senescence of dental pulp cells. In addition, the vitality of senescent cells can be damaged by exogenous ROS caused by infection, trauma, and chemicals. Therefore, such inflammatory process contributes to the impairment of pulp homeostasis and reduces the regenerative ability of dental stem cells [101]. Considering the interactions mentioned above, the future of dental pulp therapies involves the selective elimination of senescent cells, also known as senolysis, which prevents various age- related diseases. Most oral pathologies are implicated on the effects of ageing without exerting undesirable side effects. Senolytic drugs are increasing the resistance to ageing. They may delay or treat the geriatric syndromes and extend the life of the organisms. Studies have shown that several senolytics, including dasatinib plus quercetin, 17-DMAG, and navitoclax, are effective in reducing senescent cell burden in mice with decreases in senescent cell indicators such as cellular senescence-associated β-galactosidase activity. Therefore, 1st generation senolytics such as dasatinib, quercetin, fisetin and curcumin, and 2nd generation senolytic therapies such as immunomodulators are considered as future therapeutic drugs of senolysis [102, 103, 104, 105, 106].
Conclusion
ROS and inflammation play a significant regulatory role in cell signaling mechanism which controls the balance between regeneration and tissue damage, and these are essential processes for stem cell metabolism and stem cell- based therapies. Indeed, there is interplay between both phenomena as the accumulated ROS can directly induce the secretion of inflammatory mediators and stimulation of inflammation, the other way around, also inflammatory cells promote oxidative stress by releasing multiple reactive species at sites of inflammation. Key molecules in transcription, protein modulation, and protein stability can be modified for regulation of oxidative stress and inflammation which is fundamental for future studies to improve stem cell proliferation, differentiation, and survival as well as to prevent cell senescence and ageing process. Since dental pulp stem cell-based therapies have gained attention in this field lately, future therapies are also necessary to focus on eliminating senescent cells and improvement of senolytic drugs in terms of resisting ageing.
References
-
Brignier AC, Gewirtz AM (2010) Embryonic and Adult Stem Cell Therapy. J Allergy Clin Immunol 125(2): 336- 344.
-
Baharvand H (2009) Trends in Stem Cell Biology and Technology. N.Y.: Humana Press.
-
Gurusamy N, Alsayari A, Rajasingh S, Rajasingh J (2018) Adult Stem Cells for Regenerative Therapy. Prog Mol Biol Transl Sci 160: 1-22.
-
Han Y, Li X, Zhang Y, Han Y, Chang F, Ding J (2019) Mesenchymal Stem Cells for Regenerative Medicine. Cells 8(8): 886.
-
Parekkadan B, Milwid JM (2010) Mesenchymal Stem Cells as Therapeutics. Annu Rev Biomed Eng 12: 87-117.
-
Friedenstein AJ, Petrakova KV, Kurolesova AI, Frolova GP (1968) Heterotopic of Bone Marrow. Analysis of Precursor Cells for Osteogenic and Hematopoietic Tissues. Transplantation 6(2): 230-247.
-
Li Q, Gao Z, Chen Y, Guan MX (2017) The Role of Mitochondria in Osteogenic, Adipogenic and Chondrogenic Differentiation of Mesenchymal Stem Cells. Protein Cell 8(6): 439-445.
-
Marquez-Curtis LA, Janowska Wieczorek A, McGann LE, Elliott JA (2015) Mesenchymal Stromal Cells Derived from Various Tissues: Biological, Clinical and Cryopreservation Aspects. Cryobiology 71(2):181-197.
-
Hocking DC (2015) Therapeutic Applications of Extracellular Matrix. Adv Wound Care (New Rochelle) 4(8): 441-443.
-
Mao X, Liu Y, Chen C, Shi S (2017) Mesenchymal Stem Cells and Their Role in Dental Medicine. Dent Clin North Am 61(1): 161-172.
-
Tsutsui TW (2020) Dental Pulp Stem Cells: Advances to Applications. Stem Cells Cloning 13: 33-42.
-
Hofmann AD, Beyer M, Krause-Buchholz U, Wobus M, Bornhauser M, et al. (2012) OXPHOS Supercomplexes as a Hallmark of the Mitochondrial Phenotype of Adipogenic Differentiated Human Mscs. PLoS One 7(4): e35160.
-
Hsu YC, Wu YT, Yu TH, Wei YH (2016) Mitochondria in Mesenchymal Stem Cell Biology and Cell Therapy: from Cellular Differentiation to Mitochondrial Transfer. Semin Cell Dev Biol 52: 119-131.
-
Tahara EB, Navarete FD, Kowaltowski AJ (2009) Tissue-, Substrate-, and Site-Specific Characteristics of Mitochondrial Reactive Oxygen Species Generation. Free Radic Biol Med 46(9): 1283-1297.
-
Pietila M, Palomaki S, Lehtonen S, Ritamo I, Valmu L, et al. (2012) Mitochondrial Function and Energy Metabolism in Umbilical Cord Blood- and Bone Marrow-Derived Mesenchymal Stem Cells. Stem Cells Dev 21(4): 575-588.
-
Varela-Rey M, Embade N, Ariz U, Lu SC, Mato JM, et al. (2009) Non-Alcoholic Steatohepatitis and Animal Models: Understanding the Human Disease. Int J Biochem Cell Biol 41(5): 969-976.
-
Sart S, Song L, Li Y (2015) Controlling Redox Status for Stem Cell Survival, Expansion, and Differentiation. Oxid Med Cell Longev 2015: 1-14.
-
Wang W, Zhang Y, Lu W, Liu K (2015) Mitochondrial Reactive Oxygen Species Regulate Adipocyte Differentiation of Mesenchymal Stem Cells in Hematopoietic Stress Induced by Arabinosylcytosine. PLoS One 10(3): e0120629.
-
Young IS, Woodside JV (2001) Antioxidants in Health and Disease. J Clin Pathol 54(3): 176-186.
-
Birben E, Sahiner UM, Sackesen C, Erzurum S, Kalayci O (2012) Oxidative Stress and Antioxidant Defense. World Allergy Organ J 5(1): 9-19.
-
Kohlgruber S, Upadhye A, Dyballa-Rukes N, McNamara CA, Altschmied J (2017) Regulation of Transcription Factors by Reactive Oxygen Species and Nitric Oxide in Vascular Physiology and Pathology. Antioxid Redox Signal 26(13): 679-699.
-
Liguori I, Russo G, Curcio F, Bulli G, Aran L, et al. (2018) Oxidative Stress, Aging, and Diseases. Clin Interv Aging 13: 757-772.
-
Phaniendra A, Jestadi DB, Periyasamy L (2015) Free Radicals: Properties, Sources, Targets, and their Implication in Various Diseases. Indian J Clin Biochem 30(1): 11-26.
-
Lu JM, Lin PH, Yao Q, Chen C (2010) Chemical and Molecular Mechanisms of Antioxidants: Experimental Approaches and Model Systems. J Cell Mol Med 14(4): 840-860.
-
Pizzino G, Irrera N, Cucinotta M, Pallio G, Mannino F, et al. (2017) Oxidative Stress: Harms and Benefits for Human Health. Oxid Med Cell Longev 2017: 8416763.
-
Ping Z, Peng Y, Lang H, Xinyong C, Zhiyi Z, et al. (2020) Oxidative Stress in Radiation-Induced Cardiotoxicity. Oxid Med Cell Longev 2020: 3579143.
-
Ozcan M, Aydemir D, Bacanli M, Anlar HG, Ulusu NN, et al. (2021) Protective Effects of Antioxidant Chlorophyllin in Chemically Induced Breast Cancer Model In vivo. Biol Trace Elem Res 199(12): 4475-4488.
-
Ozcan M, Esendagli G, Musdal Y, Canpinar H, Bacanli M, et al. (2018) Dual actions of the antioxidant chlorophyllin, a glutathione transferase P1-1 inhibitor, in tumorigenesis and tumor progression. J Cell Biochem.
-
Kim M, Kim C, Choi YS, Kim M, Park C, et al. (2012) Age- Related Alterations in Mesenchymal Stem Cells Related to Shift in Differentiation from Osteogenic to Adipogenic Potential: Implication to Age-Associated Bone Diseases and Defects. Mech Ageing Dev 133(5): 215-225.
-
Atashi F, Modarressi A, Pepper MS (2015) The Role of Reactive Oxygen Species in Mesenchymal Stem Cell Adipogenic and Osteogenic Differentiation: A Review. Stem Cells Dev 24(10): 1150-1163.
-
Rodrigues M, Turner O, Stolz D, Griffith LG, Wells A (2012) Production of Reactive Oxygen Species by Multipotent Stromal Cells/Mesenchymal Stem Cells upon Exposure to Fas Ligand. Cell Transplant 21(10): 2171-2187.
-
Lee DH, Lim BS, Lee YK, Yang HC (2006) Effects of Hydrogen Peroxide (H2O2) on Alkaline Phosphatase Activity and Matrix Mineralization of Odontoblast and Osteoblast Cell Lines. Cell Biol Toxicol 22(1): 39-46.
-
Marie PJ (2008) Transcription Factors controlling osteoblastogenesis. Arch Biochem Biophys 473(2): 98- 105.
-
Komori T (2011) Signaling Networks in RUNX2- Dependent Bone Development. J Cell Biochem 112(3): 750-755.
-
Chen CT, Shih YR, Kuo TK, Lee OK, Wei YH (2008) Coordinated Changes Of Mitochondrial Biogenesis and Antioxidant Enzymes During Osteogenic Differentiation of Human Mesenchymal Stem Cells. Stem Cells 26(4): 960-968.
-
Tan J, Xu X, Tong Z, Lin J, Yu Q, et al. (2015) Decreased Osteogenesis of Adult Mesenchymal Stem Cells by Reactive Oxygen Species Under Cyclic Stretch: A Possible Mechanism of Age Related Osteoporosis. Bone Res 3: 15003.
-
Yan H, Aziz E, Shillabeer G, Wong A, Shanghavi D, et al. (2002) Nitric Oxide Promotes Differentiation of Rat White Preadipocytes in Culture. J Lipid Res 43(12): 2123-2129.
-
Kanda Y, Hinata T, Kang SW, Watanabe Y (2011) Reactive Oxygen Species Mediate Adipocyte Differentiation in Mesenchymal Stem Cells. Life Sci 89(7-8): 250-258.
-
Tormos KV, Anso E, Hamanaka RB, Eisenbart J, Joseph J, et al. (2011) Mitochondrial Complex III ROS Regulate Adipocyte Differentiation. Cell Metab 14(4): 537-544.
-
Higuchi M, Dusting GJ, Peshavariya H, Jiang F, Hsiao ST, et al. (2013) Differentiation of human adipose-derived stem cells into fat involves reactive oxygen species and Forkhead box O1 mediated upregulation of antioxidant enzymes. Stem Cells Dev 22(6): 878-888.
-
Nightingale H, Kemp K, Gray E, Hares K, Mallam E, et al. (2012) Changes in Expression of the Antioxidant Enzyme SOD3 Occur upon Differentiation of Human Bone Marrow-Derived Mesenchymal Stem Cells In Vitro. Stem Cells Dev 21(11): 2026-2035.
-
Heywood HK, Lee DA (2017) Bioenergetic Reprogramming of Articular Chondrocytes by Exposure to Exogenous and Endogenous Reactive Oxygen Species and its Role in the Anabolic Response to Low Oxygen. J Tissue Eng Regen Med 11(8): 2286-2294.
-
Kim KS, Choi HW, Yoon HE, Kim IY (2010) Reactive Oxygen Species Generated by NADPH Oxidase 2 and 4 are Required for Chondrogenic Differentiation. J Biol Chem 285(51): 40294-40302.
-
Dayem AA, Choi HY, Kim JH, Cho SG (2010) Role of Oxidative Stress in Stem, Cancer, and Cancer Stem Cells. Cancers (Basel) 2(2): 859-884.
-
Kim TH, Woo JS, Kim YK, Kim KH (2014) Silibinin Induces Cell Death Through Reactive Oxygen Species- Dependent Downregulation of Notch-1/ERK/Akt Signaling in Human Breast Cancer Cells. J Pharmacol Exp Ther 349(2): 268-278.
-
Shi Y, Hu Y, Lv C, Tu G (2016) Effects of Reactive Oxygen Species on Differentiation of Bone Marrow Mesenchymal Stem Cells. Ann Transplant 21: 695-700.
-
Yuan T, Yang T, Chen H, Fu D, Hu Y, et al. (2019) New Insights into Oxidative Stress and Inflammation During Diabetes Mellitus-Accelerated Atherosclerosis. Redox Biol 20: 247-260.
-
Clark R, Kupper T (2005) Old Meets New: The Interaction between Innate and Adaptive Immunity. J Invest Dermatol 125(4): 629-637.
-
Takeuchi O, Akira S (2010) Pattern Recognition Receptors and Inflammation. Cell 140(6): 805-820.
-
Biswas SK (2016) Does the Interdependence between Oxidative Stress and Inflammation Explain the Antioxidant Paradox? Oxid Med Cell Longev 2016: 5698931.
-
Droge W (2002) Free Radicals in the Physiological Control of Cell Function. Physiol Rev 82(1): 47-95.
-
Popa-Wagner A, Mitran S, Sivanesan S, Chang E, Buga AM (2013) ROS and Brain Diseases: The Good, The Bad, and The Ugly. Oxid Med Cell Longev 2013: 963520.
-
Salzano S, Checconi P, Hanschmann EM, Lillig CH, Bowler LD, et al. (2014) Linkage of Inflammation and Oxidative Stress Via Release of Glutathionylated Peroxiredoxin-2, Which Acts as a Danger Signal. Proc Natl Acad Sci U S A 111(33): 12157-12162.
-
Swirski FK, Nahrendorf M, Etzrodt M, Wildgruber M, Cortez Retamozo V, et al. (2009) Identification of Splenic Reservoir Monocytes and their Deployment to Inflammatory Sites. Science 325(5940): 612-616.
-
Xiong XY, Liu L, Yang QW (2016) Functions and Mechanisms of Microglia/Macrophages in Neuroinflammation and Neurogenesis after Stroke. Prog Neurobiol 142: 23-44.
-
Kim J, Hematti P (2009) Mesenchymal Stem Cell- Educated Macrophages: A Novel Type of Alternatively Activated Macrophages. Exp Hematol 37(12): 1445- 1453.
-
Villalta SA, Nguyen HX, Deng B, Gotoh T, Tidball JG (2009) Shifts in Macrophage Phenotypes and Macrophage Competition for Arginine Metabolism Affect the Severity of Muscle Pathology in Muscular Dystrophy. Hum Mol Genet 18(3): 482-496.
-
Sehgal A, Donaldson DS, Pridans C, Sauter KA, Hume DA, et al. (2018) The Role of CSF1R-Dependent Macrophages in Control of the Intestinal Stem-Cell Niche. Nat Commun 9(1): 1272.
-
Tidball JG (2005) Inflammatory Processes in Muscle Injury and Repair. Am J Physiol Regul Integr Comp Physiol 288(2): 345-353.
-
Spaggiari GM, Capobianco A, Becchetti S, Mingari MC, Moretta L (2006) Mesenchymal Stem Cell-Natural Killer Cell Interactions: Evidence that Activated NK Cells are Capable of Killing Mscs, Whereas Mscs can Inhibit IL-2- Induced NK-Cell Proliferation. Blood 107(4): 1484-1490.
-
Aggarwal S, Pittenger MF (2005) Human mesenchymal stem cells modulate allogeneic immune cell responses. Blood 105(4): 1815-1822.
-
Tse WT, Pendleton JD, Beyer WM, Egalka MC, Guinan EC (2003) Suppression of allogeneic T-cell proliferation by human marrow stromal cells: implications in transplantation. Transplantation 75(3): 389-397.
-
Wyles CC, Houdek MT, Behfar A, Sierra RJ (2015) Mesenchymal stem cell therapy for osteoarthritis: current perspectives. Stem Cells Cloning 8: 117-124.
-
Denu RA, Hematti P (2016) Effects of Oxidative Stress on Mesenchymal Stem Cell Biology. Oxid Med Cell Longev 2016: 2989076.
-
Herb M, Schramm M (2021) Functions of ROS in Macrophages and Antimicrobial Immunity. Antioxidants (Basel) 10(2): 313.
-
Slauch JM (2011) How does the oxidative burst of macrophages kill bacteria? Still an open question. Mol Microbiol 80(3): 580-583.
-
Huang GT, Gronthos S, Shi S (2009) Mesenchymal stem cells derived from dental tissues vs. those from other sources: their biology and role in regenerative medicine. J Dent Res 88(9): 792-806.
-
Martens W, Bronckaers A, Politis C, Jacobs R, Lambrichts I (2013) Dental stem cells and their promising role in neural regeneration: an update. Clin Oral Investig 17(9): 1969-1983.
-
Feng J, Mantesso A, De Bari C, Nishiyama A, Sharpe PT (2011) Dual origin of mesenchymal stem cells contributing to organ growth and repair. Proc Natl Acad Sci U S A 108(16): 6503-6508.
-
Kerkis I, Kerkis A, Dozortsev D, Stukart Parsons GC, Sílvia Maria Gomes Massironi, et al. (2006) Isolation and characterization of a population of immature dental pulp stem cells expressing OCT-4 and other embryonic stem cell markers. Cells Tissues Organs 184(3-4): 105-116.
-
Gronthos S, Mankani M, Brahim J, Robey PG, Shi S (2000) Postnatal human dental pulp stem cells (DPSCs) In vitro and In vivo. Proc Natl Acad Sci U S A 97(25): 13625- 13630.
-
Shi S, Gronthos S (2003) Perivascular niche of postnatal mesenchymal stem cells in human bone marrow and dental pulp. J Bone Miner Res 18(4): 696-704.
-
Dominici M, Le Blanc K, Mueller I, Slaper Cortenbach I, Marini F, et al. (2006) Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 8(4): 315-317.
-
Karbanova J, Soukup T, Suchanek J, Pytlik R, Corbeil D, et al. (2011) Characterization of dental pulp stem cells from impacted third molars cultured in low serum-containing medium. Cells Tissues Organs 193(6): 344-365.
-
Sakai K, Yamamoto A, Matsubara K, Nakamura S, Naruse M, et al. (2012) Human dental pulp-derived stem cells promote locomotor recovery after complete transection of the rat spinal cord by multiple neuro-regenerative mechanisms. J Clin Invest 122(1): 80-90.
-
Shiba H, Fujita T, Doi N, Nakamura S, Nakanishi K, et al. (1998) Differential effects of various growth factors and cytokines on the syntheses of DNA, type I collagen, laminin, fibronectin, osteonectin/secreted protein, acidic and rich in cysteine (SPARC), and alkaline phosphatase by human pulp cells in culture. J Cell Physiol 174(2): 194-205.
-
Hwang HI, Lee TH, Jang YJ (2015) Cell proliferation- inducing protein 52/mitofilin is a surface antigen on undifferentiated human dental pulp stem cells. Stem Cells Dev 24(11): 1309-1319.
-
Wang L, Cheng L, Wang H, Pan H, Yang H, et al. (2016) Glycometabolic reprogramming associated with the initiation of human dental pulp stem cell differentiation. Cell Biol Int 40(3): 308-317.
-
Lee YH, Kang YM, Heo MJ, Kim GE, Bhattarai G, et al. (2013) The survival role of peroxisome proliferator- activated receptor gamma induces odontoblast differentiation against oxidative stress in human dental pulp cells. J Endod 39(2): 236-241.
-
El Alami M, Vina Almunia J, Gambini J, Mas Bargues C, Siow RC, et al. (2014) Activation of p38, p21, and NRF-2 mediates decreased proliferation of human dental pulp stem cells cultured under 21% O2. Stem Cell Reports 3(4): 566-573.
-
Lv YJ, Yang Y, Sui BD, Hu CH, Zhao P, et al. (2018) Resveratrol counteracts bone loss via mitofilin-mediated osteogenic improvement of mesenchymal stem cells in senescence-accelerated mice. Theranostics 8(9): 2387- 2406.
-
Sloan AJ, Waddington RJ (2009) Dental pulp stem cells: what, where, how? Int J Paediatr Dent 19(1): 61-70.
-
Alaidaroos NYA, Alraies A, Waddington RJ, Sloan AJ, Moseley R (2021) Differential SOD2 and GSTZ1 profiles contribute to contrasting dental pulp stem cell susceptibilities to oxidative damage and premature senescence. Stem Cell Res Ther 12(1): 142.
-
Batouli S, Miura M, Brahim J, Tsutsui TW, Fisher LW, et al. (2003) Comparison of stem-cell-mediated osteogenesis and dentinogenesis. J Dent Res 82(12): 976-981.
-
Matsui M, Kobayashi T, Tsutsui TW (2018) CD146 positive human dental pulp stem cells promote regeneration of dentin/pulp-like structures. Hum Cell 31(2): 127-138.
-
Prescott RS, Alsanea R, Fayad MI, Johnson BR, Wenckus CS, et al. (2008) In vivo generation of dental pulp- like tissue by using dental pulp stem cells, a collagen scaffold, and dentin matrix protein 1 after subcutaneous transplantation in mice. J Endod 34(4): 421-426.
-
Nakashima M, Akamine A (2005) The application of tissue engineering to regeneration of pulp and dentin in endodontics. J Endod 31(10): 711-718.
-
Laird DJ, von Andrian UH, Wagers AJ (2008) Stem cell trafficking in tissue development, growth, and disease. Cell 132(4): 612-630.
-
Galler KM, Widbiller M (2017) Perspectives for Cell- homing Approaches to Engineer Dental Pulp. J Endod 43(9S): 40-45.
-
Galvão I, Sugimoto MA, Vago JP, Machado MG, Sousa LP (2018) Mediators of Inflammation. In: Riccardi C, et al. (Eds.), Immunopharmacology and Inflammation, Cham: Springer International Publishing, pp: 3-32.
-
Mosser DM, Edwards JP (2008) Exploring the full spectrum of macrophage activation. Nature Reviews Immunology 8(12): 958-969.
-
Cooper PR, Takahashi Y, Graham LW, Simon S, Imazato S, et al. (2010) Inflammation-regeneration interplay in the dentine-pulp complex. J Dent 38(9): 687-697.
-
Babb R, Chandrasekaran D, Carvalho Moreno Neves V, Sharpe PT (2017) Axin2-expressing cells differentiate into reparative odontoblasts via autocrine Wnt/beta- catenin signaling in response to tooth damage. Sci Rep 7(1): 3102.
-
Neves VC, Babb R, Chandrasekaran D, Sharpe PT (2017) Promotion of natural tooth repair by small molecule GSK3 antagonists. Sci Rep 7: 39654.
-
Tsunawaki S, Sporn M, Ding A, Nathan C (1988) Deactivation of macrophages by transforming growth factor-beta. Nature 334(6179): 260-262.
-
Neves VCM, Yianni V, Sharpe PT (2020) Macrophage modulation of dental pulp stem cell activity during tertiary dentinogenesis. Sci Rep 10(1): 20216.
-
Goldberg M, Farges JC, Lacerda-Pinheiro S, Six N, Jegat N, et al. (2008) Inflammatory and immunological aspects of dental pulp repair. Pharmacol Res 58(2): 137-147.
-
Galler KM, Weber M, Korkmaz Y, Widbiller M, Feuerer M (2021) Inflammatory Response Mechanisms of the Dentine-Pulp Complex and the Periapical Tissues. Int J Mol Sci 22(3): 1480.
-
Zhang F, Lau SS, Monks TJ (2011) The cytoprotective effect of N-acetyl-L-cysteine against ROS-induced cytotoxicity is independent of its ability to enhance glutathione synthesis. Toxicol Sci 120(1): 87-97.
-
Jun SK, Yoon JY, Mahapatra C, Park JH, Kim HW, et al. (2019) Ceria-incorporated MTA for accelerating odontoblastic differentiation via ROS downregulation. Dental Materials 35(9): 1291-1299.
-
Lee YH, Kim GE, Cho HJ, Yu MK, Bhattarai G, et al. (2013) Aging of In vitro pulp illustrates change of inflammation and dentinogenesis. J Endod 39(3): 340- 345.
-
Kirkland JL, Tchkonia T (2015) Clinical strategies and animal models for developing senolytic agents. Exp Gerontol 68: 19-25.
-
Baar MP, Brandt RMC, Putavet DA, Klein JDD, Derks KWJ, et al. (2017) Targeted Apoptosis of Senescent Cells Restores Tissue Homeostasis in Response to Chemotoxicity and Aging. Cell 169(1): 132-147.
-
van Deursen JM (2019) Senolytic therapies for healthy longevity. Science 364(6441): 636-637.
-
Kirkland JL, Tchkonia T (2020) Senolytic drugs: from discovery to translation. J Intern Med 288(5): 518-536.
-
Kirkland JL, Tchkonia T, Zhu Y, Niedernhofer LJ, Robbins PD (2017) The Clinical Potential of Senolytic Drugs. J Am Geriatr Soc 65(10): 2297-2301.
- Research Progress of Induced Pluripotent Stem Cells and Their Clinical Application Prospects
- Nishan Al-Kamal is the Starting Point of A Feminist Scientist
- Current Concepts and Future Perspectives of Stem Cell Therapy in Peripheral Arterial Disease
- Stem Cell and Oxidative Stress-Inflammation Cycle
- Mitochondria Targeted Antioxidants can Improve In Vitro Embryo Production in Buffalo
- Effects of Complex Treatment by Use of Fetal Stem Cells on Bone Mineral Density in Postmenopausal Women