H1N1 Vaccination Beacon of Hope to SARS-COV-2 Infection
Introduction: COVID-19 pandemic has been a nightmare for entire medical community with no definitive treatment available after nearly half a year. The impact on human civilization is farfetched. Aim: Amidst rising number of patients, countries facing second wave with no immediate respite in terms of protective vaccine the present study is a novel research that establishes link between cross protection by H1N1 Influenza vaccine and occurrence of COVID -19 infection. Material and Method: A prospective study was conducted in the department of Pathology and transfusion medicine at a tertiary care hospital in west India for time period of two months from mid-May to mid-July, 2020. The study group consisted of all adults tested for COVID-19 RTPCR for a period of two months. H1N1 vaccination with one year was considered as vaccinated and details of vaccination were collected at the time of sampling. Results: The study population consisted of 2482 patients, 186 (7049%) were positive for RTPCR and remaining 92.50% (2296) were negative. Among the total positive patients 128 (68.82%) were males and 58 (31.18%) were females. There were 2 (1.08%) COVID positive patients who had H1N1 vaccination within one year whereas 184 (99.92%) did not receive HINI vaccine at all. Among the 2296 (92.505) who tested negative for COVID PCR -124(5.40%) had H1N1 vaccination while remaining 2172 (94.60%) were not vaccinated. The chi square test revealed that H1N1 vaccinated individuals had lesser incidence of COVID PCR positivity, the finding was statistically significant (p value- 0.009748, p value <0.01) Conclusion: This is the first study to establish cross protection afforded by H1N1 vaccination against SARS COV-2 .The findings of the study will be a stimulus for more research and a ray of hope to medical fraternity globally that has been shaken by this ongoing pandemic.
Introduction
Novel Human Corona Virus (HCOV) was identified as causative agent of pneumonia in Wuhan province of China in December, 2019 and was declared as a pandemic by the World Health Organization (WHO) on March 11, 2020. The nomenclature of virus is derived from phylogenetic and taxonomic similarity with SARS –COV 1. The clinical presentation varies from totally asymptomatic individuals to mild fever, cough and dyspnoea to cytokine storm, respiratory failure, coagulopathy and unfortunate death [1]. SARS –COV 2 is closely related to SARS (named SARS –COV 1) and Middle Eastern respiratory syndrome (MERS) COVs causing zoonotic epidemic in 2003 and 2012 respectively [2]. The underlying mechanism of COVID 19 infection, its severity is not completely understood. Comorbid conditions like cardiovascular disease and respiratory diseases contribute to severity of COVID-19 infection resulting in higher morbidity and mortality [3].
The rapid spread of SARS –COV 2 and it’s unprecedented nature has made situation worse with medical facilities in research, diagnostics and therapeutics all stretched beyond limits globally [4]. There is significant generation of scientific knowledge, rapid publications within months which suggest that immune response to SARS –COV 2 are different from those seen with other Corona virus family. A spectrum of infection which is largely asymptomatic to increased fatality strongly suggest that there are differences in host immune response to SARS –COV 2 infection and hence it is imperative that immune responses against SARS –COV 2 are better elucidated for understanding entire spectrum from asymptomatic to cytokine storm.
Immunological memory is the ability of immune system to quickly and specifically recognize an antigen that the body has previously encountered and thereafter initiate an immune response. This immune memory is the basis of vaccination and involves memory T and B cells. Literature studies have shown that immune memory by related pathogens is helpful in reducing severity of disease. It has been speculated that preexisting immunity through cross reactivity to other Corona virus strains could protect against severe SARS –COV 2 infection [5, 6, 7].
The corona virus outbreak poses ongoing global threat and prompted us for looking at any possible cross protection with prior H1N1 vaccination [8, 9, 10]. The present study was aimed to search any possible link between prior H1N1 vaccination and COVID 19 infection. The H1N1 vaccination within past one year was considered to be vaccinated.
Material and Methods
A prospective study was conducted in the department of Pathology and transfusion medicine at a tertiary care hospital in west India for time period of two months from mid May to mid July, 2020. The laboratory is NABL accredited and conforms to strict protocols and standard operating procedures. The study group consisted of all adults who were tested for RT PCR COVID-19 assay. The detailed history pertaining to symptoms and H1N1 vaccination within a year was collected at the time of sampling. The data was collected from laboratory software after maintaining confidentiality of data and descriptive statistics were applied using online free statistical calculator.
Results
The study population consisted of 2482 patients who were tested for RTPCR COVID (Two step E gene and Rdrp gene detection). 186 (7049%) were positive for RTPCR and remaining 92.50% (2296) were negative. (Figure 1). Among the total positive patients 128 (68.82%) were males and 58 (31.18%) were females (Table 1). 82(44.09%) had low hemoglobin for their respective age and sex while 104 (55.91%) had normal hemoglobin values (Table 2). The absolute neutrophil count (ANC) was within normal range for 127 (68.28%) of COVID positives while neutrophilia was seen in 52 (27.96%) and neutropenia noted in 7(3.76%) (Table 3). 149 (80.11%) had lymphocyte count within normal reference range for age and 37 (19.89%) had lymphopenia (Table 4). There were 2 (1.08%) COVID positive patients who had H1N1 vaccination within one year whereas 184 (99.92%) did not receive HINI vaccine at all (Table 5). Among the 2296 (92.505) who tested negative for COVID PCR -124(5.40%) had H1N1 vaccination while remaining 2172 (94.60%) were not vaccinated (Figure 1).
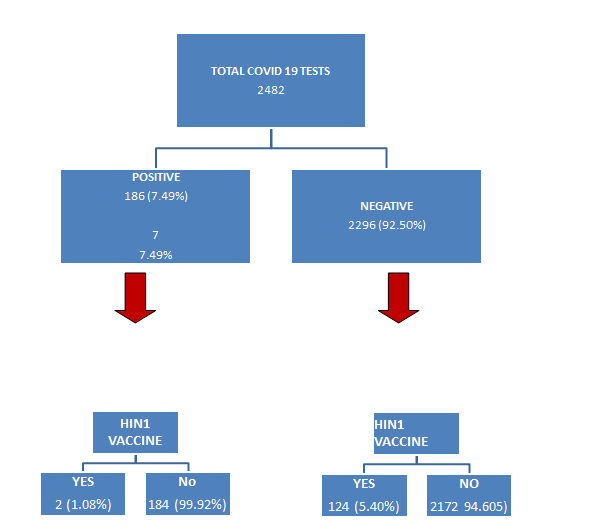
The chi square test revealed that H1N1 vaccinated individuals had lesser incidence of COVID PCR positivity, the finding was statistically significant (p value- 0.009748, p value <0.01) (Table 6).
| Female | 58 (31.18%) |
| Male | 128(68.82%) |
Table 1: Sex distribution.
| Low ( Male-<13mg/dl, Female<12mg/dl) | 82 (44.09%) |
|---|---|
| Normal | 104 (55.91%) |
| High | 52 (27.96%) |
| Low | 7 (3.76%) |
| Normal | 127 (68.28%) |
Table 2: Hemoglobin levels.
| Low | 37 (19.89%) |
| Normal | 149 (80.11%) |
Table 3: Absolute lymphocyte count (Normal 1000-3000/ cu mm).
| No | 184 (99.92%) |
| Yes | 2(1.08%) |
Table 4: H1N1 vaccination YES / NO.
| H1 N1 vaccinated | H1N1 NOT vaccinated | Row Totals | |
|---|---|---|---|
| Positive COVID | 2 (9.44) [5.87] | 184 (176.56) [0.31] | 186 |
| Negative COVID | 124 (116.56) [0.48] | 2172 (2179.44) [0.03] | 2296 |
| Column Totals | 126 | 2356 | 2482 (Grand Total) |
| The chi-square statistic is 6.6803. The p-value is .009748. The result is significant at p<.01. |
Table 5: Chi square 2 x 2.
Discussion
The Corona virus strains infecting humans belong to alpha or beta genera of corona virus. Human corona virus (HCOV-229E) and (HCOV-NL-63) belong to alpha corona virus while (HCOV-OC43) and (HCOV-HKU1) belong to beta genera of corona virus. MERS-COV, SARS COV and SARS COV- 2 also belong to beta corona virus.
The present study shows that H1N1 vaccinated individuals have less occurrence of positive PCR COVID test positivity than those who did not receive vaccine [11].
This finding is statistically significant. The subjects were all those who came or were admitted in hospital for COVID testing eliminating any bias that may arise due to application of inclusion or exclusion criteria. The only caveat in the study was lack of history of other co morbid conditions like cardiovascular disease, diabetes, immune disorders or therapies medication taken. Another limitation was the lack of complete blood count findings in COVID 19 tested negative patients as practically it was not possible to incorporate the same in given stretch of time where all efforts are directed towards evaluation of cross protectivity by H1N1 vaccine.
The recent literature studies pertaining to cross protection afforded by H1N1 vaccine against SARS COV -2 are sparse. Gunther Fink et al. have analyzed data from 92,664 clinically and molecular confirmed COVID 19 patients in Brazil and have found that recent influenza vaccine confers an average 8% lower odds of intensive care treatment in COVID 19 patients, 18% lower odds of requiring invasive respiratory support and 17% lower odds of death [12]. They have advised for promotion of influenza vaccine especially in population at high risk of severe COVID 19 infection. It is an off target protective effect of influenza vaccine driven by altered state innate immunity.
The immune response to SARS COV-2 is still not clearly understood however a brief account of immune response is discussed. SARS COV -2 genome is 29903 nucleotides having 14 open reading frames (ORF) [13]. The ORF encode for polyproteins which are cleaved by viral protease into 16 nonstructural proteins (nsp) [14]. The 4 ORF code for structural proteins as spike (S), envelope (E), membrane (M) and nucelocapsid (N) genes. There are accessory genes in between which regulate infection but don’t get incorporated into the viron. The life cycle of SARS –COV 2 starts with interaction between S protein (on viron) and virus receptors – HCOV-NL 63, SARS COV and SARS –COV 2 using Angiotensin converting enzyme -2 (ACE-2) as receptor [15, 16]. After attachment to receptor there is endocytosis, S protein cleavage is aided by low pH that brings a conformational change and helps in attachment of virus to plasma membrane releasing viral RNA into cytoplasm for translation [17]. The S protein with two domains - S1- a receptor binding domain (RBD) and S2- for fusion of virus with host membrane having fusion peptide [18]. Protective viral response is conferred by S protein as antibodies against S protein block ACE-2 receptors by binding to RBD, this RBD shows cross reactivity [19, 20].
The neutralizing antibody against SARS COVs can prevent viral entry and the use of convalescent plasma from recovered patients give passive transfer of antibodies which is currently tried as treatment for COVID 19 [21, 22, 23]. In a recent study, Pinto et al. elucidated that naturally isolated SARS COV 1 anti S IgG cross reacts with SARS COV 2 S glycoprotein when trans infected into Chinese hamster ovary (CHO) cells [24, 25, 26].
Lymphocyte antigen 6 complex locus E (LY6E) in studies by Pfaender ar al have shown to interfere with SARS COV-2 spike S protein mediated membrane fusion. At the same time COVs have also evolved mechanisms to inhibit IFN 1 induction and signaling [27]. COV evades pattern recognition receptors (PRR) sensing by avoiding recognition or antagonizing their action [28, 29, 30, 31, 32]. IFN is protective early in disease but later becomes pathologic probably due to interferon induced ACE2 upregulation in airway epithelium [33]. The NSP 9 and NSP 10 might induce IL-6 and IL-8 production, by potential inhibition of NKFF an endogenous NF-kB receptor [34]. The mucosal immune responses in any infection are regulated by conventional dendritic cells (CDCs), monocyte derived DC (mo DC), plasmacytoid DC and macrophages [35]. COVID-19 infection causes dysregulation of these responses but there is inadvertent collusions and need further studies [36]. The innate immune lymphoid cells comprising of cytotoxic (Natural killer, NK) and non cytotoxic helper cells. The recent studies have shown decreased NK cells in peripheral blood of COVID 19 infected patients. NK cell regulate cytotoxicity, induce lysis of virus infevted cells that up regulate virus derived proteins which are recognized by NK cell activation receptors [37, 38, 39, 40].
Lymphopenia was seen in SARS COV-1 infection and also seen in moderate to severe COVID -19 [41]. CD4 T cell for antibody production and CD8 cells for directly killing infected cells are designated cell response against any infection [31, 42]. The extent of lymphopenia is more on CD8 cells so as to the point of exhaustion. Lymphopenia also seem to correlate with serum IL6, IL10and TNF alpha and convalescent patients were found to restore bulk of T cell with overall lowering of pro inflammatory levels [43, 44, 45]. Tocilizumab, an IL6 antagonist was found to increase number of circulating lymphocytes [31, 43, 45]. The immunogenic T cell epitopes are seen across SARS COV1 protein (S,N M) and CD4 T cell response more restricted to S protein [46, 47]. The B cell response is robust and antibody binding SARS COV-2 internal N protein and external S protein are commonly seen, the receptor binding domain of S protein is highly immunogenic and these antibodies can potentially be neutralizing antibodies [48, 49, 50]. The IgG memory cells specific to RBB have been identified in blood of COVID-19 patients and in macaques where it was found to be protective against re infection [50, 51].
The recent study by Chloe H Lee, et al. investigated extent of T cell cross reactivity against SARS COV-2 that could be conferred by other corona and Influenza virus [52]. In their study they generated map of public and private predicted CD8+ T cell epitopes between Corona virus. They found sequence dissimilarity between Influenza virus and Corona virus, although they didn’t find peptide identical match between two strains and all peptides were distinct by at least three amino acids, however those with three amio acid differences, the predicted epitopes were shared across multiple corona virus strains. Accordingly, these epitopes pose potential to be cross protective within corona virus with given modest sequence similarity with epitopes derived from Influenza virus and also pose a potential to cross react against Influenza virus. The findings of the present study are in resonance with predicted cross protection by Chole H Lee, et al.
The findings of the present study emphasize on prevention rather than on cure. It shall be stimulus for more research in this context as the pandemic is ongoing affecting globally all facets of life – medical, social, economical bringing our civilization to a standstill lockdown state. In such a devastating scenario with absence of definitive treatment with no potential vaccine for next few months, the present study could spark a ray of hope and evidence that people at risk of developing COVID 19 disease might benefit significantly from influenza vaccination.
Conclusion
To our knowledge our results represent first evidence of cross protection afforded by H1N1 vaccine against SARS COV-2 infection. More studies are required in this regard as cross protection by H1N1 vaccine in COVID -19 could be a game changer for all including policy makers, medical fraternity and vaccine developers, God knows as the COVID mystery unfolds could our findings be like a drowning man to clutch the straw.
References
-
Coronaviridae Study Group of the International Committee on Taxonomy of Viruses (2020) The species severe acute respiratory syndrome-related corona virus: classifying 2019nCoV and naming it SARS-CoV-2. Nat Microbiol 5: 536-544.
-
Huang C, Wang Y, Li X, Ren L, Zhao J, et al. (2020) Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 395(10223): 497-506.
-
Dowd JB, Andriano L, Brazel DM, Rotondi V, Block P, et al. (2020) Demographic science aids in understanding the spread and fatality rates of COVID-19. PNAS 18: 9696- 9698.
-
Caramelo F, Ferreira N, Oliveiros B (2020) Estimation of risk factors for COVID-19 mortality - preliminary results. MedRxiv.
-
Epstein SL, Price GE (2010) Cross-protective immunity to influenza A viruses. Expert Rev Vaccines 9(11): 1325- 1341.
-
Frank SA (2002) Immunological Variability of Hosts. Immunology and Evolution of Infectious Disease.
-
Auladell M, Jia X, Hensen L, Chua B, Fox A, et al. (2019) Recalling the Future: Immunological Memory toward Unpredictable Influenza Viruses. Front Immunol 10.
-
Nickbakhsh S, Ho A, Marques DFP, McMenamin J, Gunson RR, et al. (2020) Epidemiology of seasonal coronaviruses: Establishing the context for COVID-19 emergence. Medrxic.
-
Khan S, Nakajima R, Jain A, Ramiro de Assis R, Jasinskas A, et al. (2020) Analysis of Serologic Cross-Reactivity between Common Human Coronaviruses and SARS- CoV-2 Using Coronavirus Antigen Microarray. Biorxiv.
-
Cheng M, Chan CW, Cheung RC, Bikkavilli RK, Zhao Q, et al. (2005) Cross-reactivity of antibody against SARS-coronavirus nucleocapsid protein with IL-11. Biochemical and Biophysical Research Communications 338(3): 1654-1660.
-
Lim YX, Ng YL, Tam JP, Liu DX (2016) Human Coronaviruses: A Review of Virus–Host Interactions. Diseases 4.
-
Fink G, Orlova-Fink N, Schindler T, Grisi S, Paula Ferrer A, et al. (2020) Inactivated trivalent influenza vaccine is associated with lower mortality among Covid 19 patients in Brazil. medRxiv.
-
Ogishi M, Yotsuyanagi H (2019) Quantitative Prediction of the Landscape of T Cell Epitope Immunogenicity in Sequence Space. Front Immunol 10: 827.
-
Guan WJ, Ni Z, Hu Y, Liang W, Ou C, et al. (2020) Clinical characteristics of coronavirus disease 2019 in China. N Engl J Med 382: 1708-1720.
-
Hofmann H, Pyrc K, Hoek L, Geier M, Berkhout B, et al. (2005) Human coronavirus NL63 employs the severe acute respiratory syndrome coronavirus receptor for cellular entry. Proc Natl Acad Sci USA 102: 7988-7993.
-
Li W, Moore MJ, Vasilieva N, Sui J, Wong SK, et al. (2003) Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 426: 450-454.
-
Hoffmann M (2020) SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 181: 271-280.
-
Thiel V, Ivanov KA, Putics A, Hertzig T, Schelle B, et al. (2003) Mechanisms and enzymes involved in SARS coronavirus genome expression. J Gen Virol 84(9): 2305- 2315.
-
Masters PS (2006) The molecular biology of coronaviruses. Adv Virus Res 66: 193-292.
-
Zhu Z, Chakraborti S, He Y, Roberts A, Sheahan T, et al. (2007) Potent cross-reactive neutralization of SARS coronavirus isolates by human monoclonal antibodies. PNSA 104(29): 12123-12128.
-
Yuan M, Wu NC, Zhu X, Lee CCD, So RTY, et al. (2020) A highly conserved cryptic epitope in the receptor- binding domains of SARS-CoV-2 and SARS-CoV. Science 368(6941): 630-633.
-
Wang C, Li W, Drabek D, Okba NMA, Haperen R, et al. (2020) A human monoclonal antibody blocking SARS- CoV-2 infection. Nat Commun 11: 2251.
-
Buchholz UJ, Bukreyev A, Yang L, Lamirande EW, Murphy BR, et al. (2004) Contributions of the structural proteins of severe acute respiratory syndrome coronavirus to protective immunity. PNSA 101(26): 9804-9809.
-
Duan K, Liu B, Li C, Zhang H, Yu T, et al. (2020) Effectiveness of convalescent plasma therapy in severe COVID-19 patients. PNSA 117(17): 9490-9496.
-
Bloch EM, Shoham S, Casadevall A, Sachais BS, Shaz B, et al. (2020) Deployment of convalescent plasma for the prevention and treatment of COVID-19. J Clin Invest 130(6): 2757-2765.
-
Pinto D, Park YJ, Beltramello M, Walls AC, Tortorici MA, et al. (2020) Structural and functional analysis of a potent sarbecovirus neutralizing antibody. bioRxiv.
-
Pfaender S, Mar KB, Michailidis E, Kratzel A, Hirt D, et al. (2020) LY6E impairs coronavirus fusion and confers immune control of viral disease. bioRxiv.
-
Zhao X, Zheng S, Chen D, Zheng M, Li X, et al. (2020) LY6E Restricts the Entry of Human Coronaviruses, including the currently pandemic SARS-CoV-2. bioRxiv.
-
Blanco-Melo D, Nilsson-Payant BE, Liu WC, Uhl S, Hoagland D, et al. (2020) Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19. Cell 181: 1036-1045.
-
Hackbart M, Deng X, Baker SC (2020) Coronavirus endoribonuclease targets viral polyuridine sequences to evade activating host sensors. PNAS 117(14): 8094- 8103.
-
Chen X, Ling J, Mo P, Zhang Y, Jiang Q, et al. (2020) Restoration of leukomonocyte counts is associated with viral clearance in COVID-19 hospitalized patients. medRxiv.
-
Deng X, Hackbart M, Mettelman RC, O’Brien A, Mielech A, et al. (2017) Coronavirus nonstructural protein 15 mediates evasion of dsRNA sensors and limits apoptosis in macrophages. PNAS 114(21): E4251-E4260.
-
Channappanavar R, Fehr AR, Zheng J, Wohlford-Lenane C, Abrahante JE, et al. (2019) IFN-I response timing relative to virus replication determines MERS coronavirus infection outcomes. J Clin Invest 129(9): 3625-3639.
-
Ziegler CJK, Allon SJ, Nyquist SK, Mbano IM, Miao VN, et al. (2020) SARS-CoV-Receptor ACE2 Is an Interferon- Stimulated Gene in Human Airway Epithelial Cells and Is Detected in Specific Cell Subsets Across Tissues. Cell 181(5): 1016-1035.
-
Lei L, Qian H, Yang X, Zhou X, Zhang X, et al. (2020) The phenotypic changes of gd T cells in COVID-19 patients. medRxiv.
-
Guilliams M, Lambrecht BN, Hammad H (2013) Division of labor between lung dendritic cells and macrophages in the defense against pulmonary infections. Mucosal Immunol 6(3): 464-473.
-
Song CY, Xu J, He JQ, Lu YQ (2020) COVID-19 early warning score: a multi-parameter screening tool to identify highly suspected patients. medRxiv.
-
Wang F, Nie J, Wang H, Zhao Q, Xiong Y, et al. (2020) Characteristics of peripheral lymphocyte subset alteration in COVID-19 pneumonia. J Infect Dis 221(11): 1762-1769.
-
Yudanin NA, Schmitz F, Flamar AL, Thome JJC, Tait Wojno E, et al. (2019) Spatial and Temporal Mapping of Human Innate Lymphoid Cells Reveals Elements of Tissue Specificity. Immunity 50(2): 505-519.e4.
-
Zheng HY, Zhang M, Yang CX, Zhang N, Wang XC, et al. (2020) Elevated exhaustion levels and reduced functional diversity of T cells in peripheral blood may predict severe progression in COVID-19 patients. Cell Mol Immunol 17: 541-543.
-
Duev-Cohen A, Bar-On Y, Glasner A, Berhani O, Ophir Y, et al. (2016) The human 2B4 and NTB-A receptors bind the influenza viral hemagglutinin and co-stimulate NK cell cytotoxicity. Oncotarget 7(11): 13093-13105.
-
Ni L, Ye F, Chen ML, Feng Y, Deng YQ, et al. (2020) Detection of SARS-CoV-2-Specific Humoral and Cellular Immunity in COVID-19 Convalescent Individuals. Immunity 52(6): 971-977.e3.
-
Liu J, Li S, Liu J, Liang B, Wang X, et al. (2020) Longitudinal characteristics of lymphocyte responses and cytokine profiles in the peripheral blood of SARS-CoV-2 infected patients. EBio Medicine 55: 102763.
-
Tan L, Wang Q, Zhang D, Ding J, Huang Q, et al. (2020) Lymphopenia predicts disease severity of COVID-19: a descriptive and predictive study. Signal Transduction and Targeted Therapy 5: 33.
-
Diao B, Wang C, Tan Y, Chen X, Liu Y, et al. (2020) Reduction and Functional Exhaustion of T Cells in Patients with Coronavirus Disease 2019 (COVID-19). Front Immunol.
-
Giamarellos-Bourboulis EJ, Netea MG, Rovina N, Akinosoglou K, Antoniadou A, et al. (2020) Complex Immune Dysregulation in COVID-19 Patients With Severe Respiratory Failure. Cell Host Microbe 27(6): 992-1000.e3.
-
Lei C, Fu W, Qian K, Li T, Zhang S, et al. (2020) Neutralization of SARS-CoV-2 spike pseudotyped virus by recombinant ACE2-Ig. Nat Commun 11: 2070.
-
Tian X, Li C, Huang A, Xia S, Lu S, et al. (2020) Potent binding of 2019 novel coronavirus spike protein by a SARS coronavirus-specific human monoclonal antibody. Emerg Microbes Infect 9(1): 382-385.
-
Amanat F, Stadlbauer D, Strohmeier S, Nguyen THO, Chromikova V, et al. (2020) A serological assay to detect SARS-CoV-2 seroconversion in humans. Nat Med 26: 1033-1036.
-
Ju B, Zhang Q, Ge J, Wang R, Sun J, et al. (2020) Human neutralizing antibodies elicited by SARS-CoV-2 infection. Nature 584: 115-119.
-
Ba L, Deng W, Huang B, Gao H, Liu J, et al. (2020) The Pathogenicity of SARS-CoV-2 in hACE2 Transgenic Mice. Nature 583: 830-833.
-
Chloe HL, Mariana Pereira P, Buckley P, Woodhouse I, Ogg G, et al. (2020) CD8+ T cell cross-reactivity against SARS-CoV-2 conferred by other coronavirus strains and influenza virus. bioRxiv.
- Psychogenic Erectile Dysfunction in Late Adulthood: A Case Report on Clinical Intervention and Intimacy Restoration
- Clinical Trials on COVID-19 in 2025: A New Chapter in Global Health Research
- Innovations and Challenges in Contemporary Medical Clinical Trials: An Editorial Perspective
- Innovations and Challenges in Contemporary Medical Clinical Trials: A Critical Perspective
- Reimagining Clinical Trials: The Power of Continuous Feedback from Medical Reports
- Factors Influencing Brain Drain: Perspectives from a Medical School in Turkey