Reduction of Sleep Spindles in Children and Adolescents with Major Depression is Age and Sex-Dependent
<p style="text-align: justify;">Background: Major Depressive Disorder (MDD) is associated to substantial sleep disturbances and reduced neuroplasticity. Sleep spindles, involved in neuroplasticity mechanisms during brain development, are more abundant in children and adolescents than in adults, and as such, might be a sensitive measure of sleep alterations in early-onset MDD. This study investigated the effects of sex and age on sleep spindles in children and adolescents with MDD. Methods: The study included 69 depressed [34 children 8-12 years of age (18 males and 16 females) and 35 adolescents 13-18 years of age (14 males and 21 females)], and 42 healthy [18 children 8-12 years of age (10 males and 8 females) and 24 adolescents 13-18 years of age (12 males and 12 females)] youth. All participants maintained a regular sleep-wake schedule for 5 days followed by two consecutive nights in the laboratory. Spindle density (SD) was analyzed in Stage 2 of non-rapid eye movement sleep. Results: Children and adolescents with MDD showed lower SD than healthy controls (p<.02). SD was lower in depressed female children (p=0.006) and depressed adolescent males (p=0.01) compared to their corresponding healthy controls. Conclusions: Low sleep spindles generation characterized children and adolescents with MDD, indicating that low sleep-dependent neuroplasticity are a feature of early-onset MDD. Young girls and adolescent boys with MDD showed the lowest spindle generation, suggesting that sleep-associated neuroplasticity alterations in MDD are moderated by sex and age. The earlier reduction in sleep-dependent neuroplasticity in depressed females, compared to depressed males, may increase susceptibility for depressive episodes in subsequent years.</p>
Introduction
Major depressive disorder (MDD) affects children and adolescents at a rate of 5 to 11% [1]. It is well known that the increase in incidence of MDD during this critical period of brain development is related to greater expression of the illness in girls, with 2:1 female/male ratio [2, 3]. MDD frequently involves delays in social, emotional, and cognitive development [4, 5, 6] and substantial decreases in neuroplasticity [7].
Nursing & Healthcare International Journal
There is compelling evidence from human and animal studies suggesting that MDD is characterized by reduced neuroplasticity that include structural and functional abnormalities [7, 8]. Neuroimaging research and postmortem studies in humans have shown atrophy in the hippocampus and the prefrontal cortex, demonstrated by reduced neuronal size and volume [9, 10, 11, 12]. These findings have been confirmed in animal models of depression [13, 14]. In addition, neurotrophic factors such as brain-derived neurotrophic factor (BDNF), known to mediate neuroplasticity during brain maturation and memory formation [15], shows reduced expression in postmortem tissue of those diagnosed with MDD [16, 17] and animal models of depression [18, 19]. Moreover, studies utilizing the widely accepted model of synaptic plasticity (long-term potentiation, LTP) have reported decreased LTP in the hippocampus [20] and the thalamocortical network [21] of animal models of MDD. Altogether, this is solid evidence that MDD is associated with impaired neuroplasticity in the cortical-limbic network [7, 8]. It is well documented that MDD is accompanied by sleep disturbances in more than 90% of the patients [22], and could reflect impairment in sleep-dependent neuroplasticity. Numerous studies have recognized the role of sleep in neuroplasticity mechanisms underlying the processes of brain maturation and memory consolidation [23, 24]. In particular, sleep spindles are bursts of activity (frequency: 12-16Hz; duration: 0.5-3 sec) generated in the thalamocortical network and constitute the hallmark of stage 2 non-Rapid Eye Movement (NREM) sleep [25, 26]. Spindles are thought to play a role in neuroplasticity by synchronizing the flow of information from limbic structures to neocortex [27, 28, 29], where they enhance cortical plasticity through an LTP- like mechanism [30]. Spindles developmental time course follows an increase after age 3, peak during adolescence, and decline there after [31, 32], suggesting a role for this sleep event in early-life brain maturation. For example, studies have shown that infants with abnormal spindle generation present with low developmental quotient and mental retardation [33, 34]. Research on the role of sleep on memory consolidation indicates that spindles increase after a learning task [35], an effect observed in young but not older adults [36] further supporting a role for spindles in early-life brain neuroplasticity. Sex differences in spindle generation have been studied in healthy adults, finding more abundant spindles in females than males [32]. To the best of our knowledge, the interaction of age and sex differences in spindles have not been addressed in healthy or MDD children and adolescents. Age and sex provide additional degrees of neuroplasticity that could impact onset and maintenance of MDD. As mentioned above, sex differences in MDD emerge during the adolescent period, with a higher incidence in females than males [2]. There have been few studies of sleep spindles in MDD with contradictory results [37] and our own study [38] showed a decrease in spindles in depressed patients compared to healthy individuals whereas two other studies found no difference in spindles between depressed and healthy individuals [39, 40]. These last two studies did not include children and adolescents in their samples, which considering the changes in spindles during development [31] is a considerable limitation. In our previous study, we showed an influence of sex on spindle generation in early-onset MDD, with prevalence in females. However, we could not investigate the effect of age on early-onset MDD due to the limited number of subjects in the sample. The present study utilized a larger sample to investigate the interaction of age and sex, two important factors contributing to spindle generation and MDD onset, in children and adolescents with MDD and healthy controls. Based on the high incidence of spindles and the higher prevalence of MDD in females, we hypothesize that the largest SPA changes will occur in depressed females compared to healthy females.
Methods
Participants
Initially 114 children (age 8-12) and adolescents (age 13-18), 69 depressed (MDD) and 45 healthy controls (HC) were recruited for the study. Of the 114 participants enrolled in the study, data were excluded from analysis due to technical difficulties such as: electrode loss (one healthy female child), and errors with the automatic spindle detection software caused by artifacts in the EEG (two healthy adolescent males). The final sample included 69 depressed [thirty-four children 8-12 years of age (18 males and 16 females) and thirty-five adolescents 13-18 years of age (14 males and 21 females)], and 42 healthy [eighteen children 8-12 years of age (10 males and 8 females) and twenty-four adolescents 13-18 years of age (12 males and 12 females)] youth. Participants were recruited through published advertisements and posted flyers at community centers, Nursing & Healthcare International Journal
hospitals, outpatient psychiatric clinics, and pediatric clinics. Self-referral or referral from a community clinician was also permitted. Inclusion criteria for all participants consisted of ability to provide informed written consent (parent) and assent (child), and no medication (for at least 4 weeks) or counseling at the time of the clinical interview. Additional inclusion criteria for healthy controls were no personal or family history of psychopathology in first-degree relatives. For the MDD group, inclusion required a current diagnosis of nonpsychotic MDD, single or recurrent, according to Diagnostic and Statistical Manual of Mental Disorders, Fourth Edition (DSM-IV), and symptomatic at the time of enrollment. Exclusion criteria for all participants included significant head injury or unconsciousness for > 5 minutes (lifetime); independent/intrinsic sleep disorder based on history or polysomnography; significant previous or concurrent medical illness by history, physical exam, or clinical lab tests. Additional exclusion criteria included lifetime diagnosis of anorexia or bulimia or substance abuse in the last 6 months.
Clinical Assessment
A brief telephone screen was administered to determine potential eligibility. All eligible participants were then scheduled for a full clinical interview. Prior to the initial interview, the study was explained and written informed consent was obtained from the parent(s) and assent from the participant. All study participants underwent the same initial psychiatric evaluations. At the initial visit, each participant and parent(s) was interviewed separately using the Schedules for Affective Disorders and Schizophrenia for School-Aged Children: Present and Lifetime [41]. Additionally, depressive- symptom severity was assessed using the Children’s Depression Rating Scale-Revised (CDRS-R) [42]. A minimum score of > 40 on the CDRS-R was required for entry into the study, indexing moderate depressive- symptom severity and matching the criterion of our previous work [38, 43, 44, 45]. The Children’s Global Assessment Scale (CGAS) [46] and the Family Global Assessment Scale (FGAS) [47] assessed overall functioning of the child and the family, respectively. Tanner maturation (1-5 scores) was self-assessed by participants using the “Typical Progression of Pubertal Development Chart” adapted from Tanner [48]. Breast and pubic hair development were assessed for girls, and genital development was assessed for boys.
Procedures
Sleep recording and scoring: All participants agreed to follow their usual school-week’s bed- and rise-times schedule established by sleep history, throughout the study. Actigraphs (Actiwatch-LTM, Mini-Mitter) were worn throughout the week and sleep/wake diaries were collected daily during the home recording period. Data from the actigraphs were downloaded prior to their first night in the laboratory to ensure that participants adhered to their regularized rise- and bed-times. This is a very simple experimental design that resulted in no protocol violations. Each participant spent 2 consecutive nights in the Sleep Laboratory. Night 1 served as laboratory adaptation and as an additional screen for the presence of independent sleep disorders, and night 2 as baseline or study night. Night 1 recordings also included chest and abdomen respiration bands, nasal-oral thermistors, and leg electrodes. Electrode placement on the baseline night included F3, F4, C3, C4, O1, O2, P3, P4, left and right EOG, recorded from the upper and lower canthi, and a bipolar, chin-cheek EMG. EEG electrodes were referenced to the ear lobes linked through a 10 KΩ resistor to minimize non-homogeneous current flow and potential artifactual hemispheric asymmetries, as is standard in our laboratory. EEG was transduced by Grass™ P511 A/C amplifiers set at sensitivity of 5 (50μV, 0.5 s calibration), corresponding to a gain of 50,000. The half-amp low- and high-bandpass filters were set at 0.3 and 30 Hz, respectively. A 60-Hz notch filter attenuated electrical noise. Visual stage scoring of 30 second epochs was conducted according to standard sleep staging criteria, described in Rechtschaffen and Kales [49], by research personnel trained at better than 90 % agreement on an epoch-by-epoch basis. Sleep latency was defined as the first consecutive 10 minute block of any sleep stage (except REM) with no more than 2 minutes of waking time, reflecting persistent sleep onset. Total sleep period was defined as the time from lights out to lights on. REM latency was defined as the minutes from sleep onset to the first epoch of REM sleep with no minimum duration criterion. Sleep efficiency is calculated as the total amount of sleep time divided by the total sleep period. The number of arousals was defined as the total number of waking episodes of at least 30 sec duration. The personnel Nursing & Healthcare International Journal
who scored the records were blind to the diagnostic group, age, and sex. Spindle detection and analysis: Sleep spindles were analyzed in the left-frontal (F3) electrode on stage 2 segments of NREM sleep. Spindles were automatically detected utilizing the software HypnoLab 1.2 (SWS Soft, Italy). The detection criterion in the HypnoLab program was set to identify spindles with frequency of 11–16Hz, amplitude of 14µV or higher, and duration of 0.5–3sec. Following automatic detection of spindles by the software, a researcher, blind to the clinical condition, age, and sex visually confirmed each of the spindles selected by the program. Spindle density (SD) was defined as the number of 0.5 sec spindles per number of stage 2 epochs, and was the dependent variable in all analyses. Statistical analysis: The Statistical Analysis Software (SAS) version 9.1 for Windows was utilized for all statistical analyses. We conducted two sets of analyses to investigate the effect of early-onset MDD and sex on age- related changes in sleep spindles based on 1) chronological age using a cutpoint of 12 years to contrast children and adolescents, and 2) maturational age using a Tanner developmental score cutpoint of 3 to contrast prepubertal (Tanner Stages 1&2) and pubertal (Tanner Stages; 3-5) youth. A 3-way analysis of variance (ANOVA) assessed the diagnostic group by sex by age interaction on spindle density (SD). Least-squares multiple comparisons tested differences between individual means at an experiment-wise p<0.05, only if a significant overall ANOVA effect was obtained. The experiment-wise probability is equivalent to a Bonferroni adjustment and provides protection against Type 1 errors. Data are shown as the mean ± standard deviation, unless otherwise specified.
Results
Demographic and Clinical Characteristics of the Sample
Demographic and clinical information are shown in (Table 1). Tanner scores were missing for one MDD female child and analyses were based on n=15. Healthy control males and females did not differ on age (F3,110=0.5 p=.40) or Tanner scores (F3,109=1.0 p=.50). As expected, there were differences between the MDD and the HC groups for the Family Global Assessment Scale (FGAS: F3,110=104.7, p<.0001), the Children’s Global Assessment Scale (CGAS: F3,110=408.5, p<.0001), and the Children’s Depression Rating Scale (CDRS: F3,110=262.1, p<.0001).
| HC | MDD | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Males (n=22) | Females (n=20) | Males (n=32) | Females (n=37) | |||||||||
| Age | 12.5 ± 2.4 | 12.9 ± 3.0 | 11.9 ± 3.0 | 12.4 ± 2.8 | ||||||||
| ≤ 12 years | 10.3 ± 1.4 | 9.8 ± 1.5 | 9.4 ± 1.0 | 9.6 ± 1.5 | ||||||||
| ≥ 13 years | 14.5 ± 1.0 | 14.9 ± 1.7 | 15.0 ± 1.4 | 14.6 ± 1.3 | ||||||||
| Tanner score | 2.9 ± 1.4 | 3.6 ± 1.3 | 2.8 ± 1.6 | 3.1 ± 1.6 | ||||||||
| FGAS | 91.6 ± 5.8 | 91.2 ± 2.4 | 67.3 ± 9.0 | 61.6 ± 9.6 | ||||||||
| CGAS | 90.7 ± 5.2 | 89.9 ± 3.7 | 52.2 ± 5.9 | 52.4 ± 6.1 | ||||||||
| CDRS-R | 18.2 ± 1.6 | 19.3 ± 2.2 | 56.7 ± 5.7 | 58.9 ± 10.9 | ||||||||
| Age of onset | — | — | 10.8 ± 2.7 | 11.0 ± 3.0 | ||||||||
| Length current depressive episode* | — | — | 12.6 ± 17.1 | 15.3 ± 18.3 | ||||||||
| Suicide attempts, (n) | — | — | 0 | 0 | ||||||||
| Suicidal Ideation | — | — | 2.3 ± 0.7 | 2.3 ± 0.8 | ||||||||
| Family history of MDD (n) | — | — | 17 | 22 | ||||||||
| Comorbid psychiatric diagnosis (n) | — | — | 17 | 21 | ||||||||
| ADHD (n) | — | — | 9 | 10 | ||||||||
| Generalized Anxiety (n) | — | — | 5 | 4 |
Table 1: Demographic and clinical features of the sample by diagnosis and sex. Data are shown as means ± standard deviation. Bold
Nursing & Healthcare International Journal
| Dysthymic Disorder (n) | — | — | 8 | 9 |
|---|---|---|---|---|
| Phobia (n) | — | — | 1 | 2 |
| Oppositional Disorder | — | — | 0 | 4 |
Table 2: Demographic and clinical features of the sample by diagnosis and sex. Data are shown as means ± standard deviation. Bold
Table 1: Demographic and clinical features of the sample by diagnosis and sex. Data are shown as means ± standard deviation. Bold font indicates significant differences between diagnostic groups. * in months FGAS = Family Global Assessment Scale; CGAS = Clinical Global Assessment Scale; CDRS-R = Children’s Depression Rating Scale – Revised; ADHD=attention deficit hyperactivity disorder
Sleep Macroarchitecture
Chronological age: The means and standard deviations for select sleep macroarchitectural variables are presented in Table 2. A significant diagnosis by sex by age interaction was evident for total sleep time (F7,110=6.2, p<.0001), REM latency (F7,110=5.2, p<.0001), percentage of Stage 1 (F7,110=2.4 p<.03), percentage of SWS (F7,110=2.8 p<.01), and the number of arousals (F7,110=2.3 P<.04). Least-square multiple comparisons indicated that children slept longer (p range: .0002 – .001), had longer REM latency (p range: .0001 – .05), and spent more time in SWS (p range: .0001 – .05) than adolescents, independent of sex or diagnosis. No between-group difference were found for the time spent in stage 2 of NREM sleep (F7,110=1.8, p=.10), eliminating the possibility that changes in spindle activity was due to group differences in the time spent in Stage 2 sleep.
| Sleep variable | Children | Adolescents | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HC | MDD | HC | MDD | |||||||||||
| Total sleep time, min | 503.2 ± 47.2 | 500.8 ± 37.5 | 447.4 ± 40.3 | 451.6 ± 41.5 | ||||||||||
| Sleep latency, min | 10.8 ± 7.1 | 18.6 ± 15.3 | 14.8 ± 13.3 | 16.7 ± 11.3 | ||||||||||
| REM latency, min | 127.5 ± 47.5 | 130.4 ± 45.7 | 89.0 ± 37.0 | 88.1 ± 43.3 | ||||||||||
| Sleep Stage, % ‡ | ||||||||||||||
| Stage 1 | 7.8 ± 6.2 | 8.5 ± 4.3 | 8.6 ± 3.7 | 10.7 ± 6.2 | ||||||||||
| Stage 2 | 47.2 ± 7.1 | 48.9 ± 6.4 | 52.0 ± 5.8 | 50.2 ± 8.2 | ||||||||||
| SW (Stage 3 + 4) | 21.8 ± 4.4 | 19.8 ± 6.9 | 16.1 ± 5.5 | 15.0 ± 7.5 | ||||||||||
| REM | 18.9 ± 4.0 | 17.5 ± 5.1 | 18.4 ± 3.7 | 20.3 ± 4.7 | ||||||||||
| Awake and Movement | 4.1 ± 1.2 | 5.1 ± 3.4 | 4.7 ± 3.3 | 3.7 ± 1.6 | ||||||||||
| Sleep efficiency, % | 93.5 ± 2.1 | 91.5 ± 4.7 | 92.3 ± 4.4 | 92.9 ± 2.7 | ||||||||||
| Number of arousals | 27.8 ± 8.6 | 25.7 ± 7.9 | 24.1 ± 7.2 | 20.8 ± 6.8 |
Table 2: Means and standard deviations of sleep macroarchitecture by Diagnosis and Age. Bold font indicates significant differences between age groups. ‡ Relative to total sleep time Effect of MDD, sex, and age on spindle density Chronologic age: A 3-way ANOVA identified a significant diagnosis by sex by age interaction (F7,110=2.8, p<.009). Multiple comparisons revealed lower SD in MDD adolescent males compared to HC adolescent males (p<.02), while MDD female children expressed lower SD than HC female children (p<.007). HC female children also generated higher SD than HC male children (p<.009), suggesting an earlier developmental increase in spindle generation in females than males. This developmental difference in SD was not evident between HC adolescent males and females (p=.65). These group differences are depicted in detail in (Figure 1).
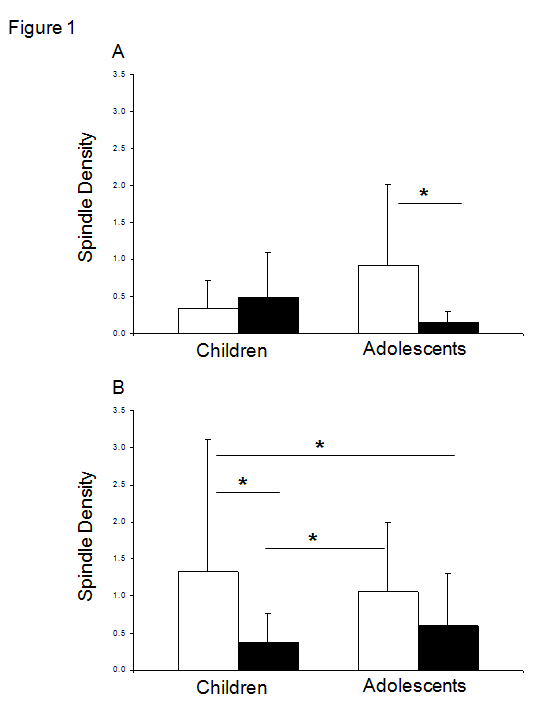
Figure 1: Spindle density in health and depressed males (A) and females (B). Open bars represent healthy children and adolescents and solid bars depressed children and adolescents. MDD adolescent males had lower spindle density than HC adolescent males. Spindle density was lower in both MDD female groups compared to HC female children. MDD female children also had lower spindle density than HC adolescent females. HC = Healthy Controls; MDD = Depressed * p<.05. Maturational age: Analysis of the group by sex by maturity effects on SD were similar to that obtained with chronological age, with a significant 3-way interaction from ANOVA (F7,110=2.6 p<.02). The lowest SD was observed in the MDD pubescent males in comparison to HC pubescent males (p<.03), and pubescent (p<.0007) and prepubescent (p<.03) HC females. All of the other MDD groups also showed significantly lower SD than HC pubescent females (p range: .02 – .03). Data are shown in (Table 3).
| HC | MDD | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PP | P | PP | P | |||||||||||||
| M | F | M | F | M | F | M | F | |||||||||
| D*Tan* S | 0.3±0. 4 | 1.3±0. 9 | 0.8±1. 0 | 1.1±1. 3 | 0.5±0. 6 | 0.3±0. 4 | 0.1±0. 1 | 0.5±0. 6 |
Table 4: Means and standard deviations for spindle density by diagnose, Tanner, and sex. HC = Healthy Controls; MDD = Major Depre
Discussion
The major finding of this study was that MDD in children and adolescents was characterized by lower spindle density than healthy controls, implying reduced brain neuroplasticity in early onset MDD. The large reduction in sleep spindles generation in MDD female children and MDD adolescent males suggests that the impact of MDD on neuroplasticity is age-and sex- dependent. The present results highlight the need to develop sex and age specific therapies to improve sleep quality rather than time spent sleep in children and adolescents with MDD, and particular emphasis should be given to the enhancement and restoration of sleep-dependent neuroplasticity mechanisms. Interpretation of our previous findings of abnormal sleep spindle generation in at-risk and depressed youth [38] and the current work raise the possibility that 1) sleep spindle abnormalities antecede the onset of MDD, underscoring the need to systematically assess sleep-dependent neuroplasticity in children and adolescents at-risk for MDD in a longitudinal study; and 2) sleep-dependent neuroplasticity abnormalities in depressed female children may increase susceptibility for future depressive episodes. The present data confirmed our previous results of decreased sleep spindle generation in early-onset MDD, but it adds the caveat that it is age specific. Our secondary analysis of sleep spindles based on pubertal stage produced identical results to those obtained contrasting the sample according to chronological age. However, males in the healthy and depressed group showed slightly lower Tanner scores than females, and the largest reduction in SD occurred in the depressed female children and depressed adolescent males. These results coincide with an earlier onset of puberty in females than in males, and the proposed role of gonadal hormones on the Nursing & Healthcare International Journal
precipitation and course of MDD [50, 51]. The earlier reduction in sleep-dependent neuroplasticity observed in MDD female children may coincide with the earlier sexual maturation that occurs in females and contribute to decrease the brain ability to cope with adverse events, increasing vulnerability for expression of subsequent depressive episodes. The lower sleep spindles generation in children and adolescents with MDD found in this study is congruent with the human and animal literature on the neurobiology of MDD showing reduced neuroplasticity such as: smaller neuronal size, fewer glial cells, shorter dendrites, lower levels of trophic factors, and reduced synaptic plasticity in the hippocampus and cortex, which indicates impairment in the cortical-limbic network [52, 8]. Additionally, a recent study utilizing an animal model of MDD showed a reduced in information flow from thalamus to cortex concomitantly with decreased LTP in this network [21]. Sleep spindles, generated in the thalamus, are thought to mediate the transfer of information from hippocampus to cortex during sleep [27, 53] and enhance synaptic transmission in cortex through an LTP-like mechanism [30]. It is possible that lower spindles generation in MDD limits the transfer of information from limbic structures to cortex, reducing/hindering cortical stimulation. Studies examining the relationship between sleep spindles generation and the aforementioned abnormalities in the cortical-limbic and thalamocortical networks in MDD, particularly during critical stages of brain development, may provide insight into the neurobiology of and risks for early-onset MDD. The current results also demonstrated higher sleep spindles generation in healthy female children than in healthy children males, a difference that was resolved in the adolescents. A previous study on the developmental time course of spindles generation showed a peak of this sleep event in adolescence [31]. However, our data suggest that spindles generation reach an earlier peak and decline in development for females than males, similar to that observed in SWA [54]. This natural decrease in neuroplasticity may have strong implications for the onset of depression and may help to elucidate the reasons for the 2:1 female to male ratio in MDD incidence that appears in adolescence. Early-life alterations in sleep spindles could cause enduring abnormalities in cortical network connectivity and reduced neuroplasticity, which may account for the higher severity and recurrence of early-onset depression compared to adult-onset depression [55]. Moreover, the decrease in spindle activity associated with MDD may also account for some of the cognitive deficit observed in people with depression [56] given the role of spindles in sleep-dependent memory consolidation [35, 57], particularly in younger but no older adults [36]. An interesting finding of this study is that spindle activity was a very accurate and sensitive measure of depression-associated changes at these early ages, even in the absence of sleep macroarchitectural alterations. Changes in sleep variables related to neuroplasticity processes such as spindle activity, but not sleep macroarchitecture variables might explain why depressed children and adolescents report significant subjective sleep complaints, but no differences in macroarchitectural sleep EEG variables compared to healthy controls [58]. Perhaps, high levels of brain neuroplasticity and homeostatic capacity in youth prevent or hinder the expression of detectable sleep macroarchitecture abnormalities. Later in life, as brain plasticity is naturally reduced and depressive symptoms continue to exert pressure, brain alterations become more established and start to appear in the sleep macroarchitecture of the patients. There are several limitations in this study. For instance, the cross-sectional approach severely limits our ability to reliably establish causality between reduced spindle activity and onset of MDD. Likewise, relevance of reduced sleep spindles to course and severity of MDD needs to be demonstrated. To date there is no evidence that depressed youth with the lowest spindle activity will have a more severe and protracted course of illness. A longitudinal design with adequate follow-up, particularly of youth at risk for MDD based on family history, will be the ideal strategy to properly address these important questions.
Key points
- MDD incidence peaks during adolescence, with higher prevalence in females, and is associated to sleep disturbances and reduced neuroplasticity
- Sleep spindles participate in neuroplasticity and reach a peak in adolescence
- MDD female children and adolescent males showed the lowest sleep spindle generation, indicating that sleep- associated neuroplasticity in MDD is moderated by sex and age
- Clinical assessment and treatment of sleep disturbances related to early-onset depression should consider sex and maturational changes to achieve better outcomes Nursing & Healthcare International Journal
• Reduced sleep-dependent neuroplasticity in depressed female children may exacerbate depressive symptomatology in their subsequent years
Acknowledgements
The authors would like to thank Raffaele Ferri, M.D. and Francesco Rundo, B.E. for technical support, the research team of the Sleep and Chronophysiology Laboratory for running subjects and Holli Bertram M.S.W. for conducting clinical interviews. This work was supported by R01-MH077744 (RA), R01-MH056953 (RA), UL-1RR024986 (JL).
References
-
Fleming JE, Offord DR (1990) Epidemiology of childhood depressive disorders: a critical review. J Am Acad Child Adolesc Psychiatry 29(4): 571-580.
-
Hankin BL, Abramson LY (1999) Development of gender differences in depression: description and possible explanations. Ann Med 31(6): 372-379.
-
Hankin BL, Abramson LY, Moffitt TE, Silva PA, McGee R, et al. (1998) Development of depression from preadolescence to young adulthood: emerging gender differences in a 10-year longitudinal study. J Abnorm Psychol 107(1): 128-140.
-
Rao U, Chen LA (2009) Characteristics, correlates, and outcomes of childhood and adolescent depressive disorders. Dialogues Clin Neurosci 11(1): 45-62.
-
Brumback RA, Weinberg WA (1990) Pediatric behavioral neurology: an update on the neurologic aspects of depression, hyperactivity, and learning disabilities. Neurol Clin 8(3): 677-703.
-
Kovacs M, Goldston D (1991) Cognitive and social cognitive development of depressed children and adolescents. J Am Acad Child Adolesc Psychiatry 30(3): 388-392.
-
Fossati P, Radtchenko A, Boyer P (2004) Neuroplasticity: from MRI to depressive symptoms. Eur Neuropsychopharmacol Suppl 5: S503-510.
-
Pittenger C, Duman RS (2008) Stress, depression, and neuroplasticity: a convergence of mechanisms. Neuropsychopharmacology 33(1): 88-109.
-
Stockmeier CA, Mahajan GJ, Konick LC, Overholser JC, Jurjus GJ (2004) Cellular changes in the postmortem hippocampus in major depression. Biol Psychiatry 56(9): 640-650.
-
Rajkowska G, Miguel-Hidalgo JJ, Wei J, Dilley G, Pittman SD et al. (1999) Morphometric evidence for neuronal and glial prefrontal cell pathology in major depression. Biol Psychiatry 45(9): 1085-1098.
-
Campbell S, Marriott M, Nahmias C, MacQueen GM (2004) Lower hippocampal volume in patients suffering from depression: a meta-analysis. Am J Psychiatry 16(4): 598-607.
-
MacQueen GM, Campbell S, McEwen BS, Macdonald K, Amano S (2003) Course of illness, hippocampal function, and hippocampal volume in major depression. Proc Natl Acad Sci U S A, 100(3): 1387- 1392.
-
Czéh B, Michaelis T, Watanabe T, Frahm J, de Biurrun G (2001) Stress-induced changes in cerebral metabolites, hippocampal volume, and cell proliferation are prevented by antidepressant treatment with tianeptine. Proc Natl Acad Sci U S A, 98(22): 12796-12801.
-
Czéh B, Perez Cruz C, Fuchs E, Flügge G (2008) Chronic stress-induced cellular changes in the medial prefrontal cortex and their potential clinical implications: does hemisphere location matter? Behav Brain Res 190(1): 1-13.
-
Citri A, Malenka RC (2008) Synaptic plasticity: multiple forms, functions, and mechanisms. Neuropsychopharmacology 33(1): 18-41.
-
Chen B, Dowlatshahi D, MacQueen GM, Wang JF, Young LT (2001) Increased hippocampal BDNF immunoreactivity in subjects treated with antidepressant medication. Biol Psychiatry 50(4): 260-265.
-
Dowlatshahi D, MacQueen GM, Wang JF, Young LT (1998) Increased temporal cortex CREB concentrations and antidepressant treatment in major depression. Lancet 352(9142): 1754-1755.
-
Smith MA, Makino S, Kvetnanský R, Post RM (1995a) Effects of stress on neurotrophic factor expression in the rat brain. Ann N Y AcadSci 771: 234-239. Nursing & Healthcare International Journal
-
Smith MA, Makino S, Kvetnansky R, Post RM (1995b) Stress and glucocorticoids affect the expression of brain-derived neurotrophic factor and neurotrophin- 3 mRNAs in the hippocampus. J Neurosci 15(3): 1768-1777.
-
Kim JJ, Diamond DM (2002) The stressed hippocampus, synaptic plasticity and lost memories. Nat Rev Neurosci 3(6): 453-462.
-
Quan M, Zheng C, Zhang N, Han D, Tian Y (2011) Impairments of behavior, information flow between thalamus and cortex, and prefrontal cortical synaptic plasticity in an animal model of depression. Brain Res Bull 85(3-4): 109-116.
-
Peterson MJ, Benca RM (2006) Sleep in mood disorders. Psychiatr Clin North Am 29(4): 1009-1032.
-
Poe GR, Walsh CM, Bjorness TE (2010) Cognitive neuroscience of sleep. Prog Brain Res 185: 1-19.
-
Peirano PD, Algarín CR (2007) Sleep in brain development. Biol Res 40(4): 471-478.
-
Steriade M (1995) Thalamic origin of sleep spindles: Morison and Bassett (1945) J Neurophysiol 73(3): 921-922.
-
Loomis AL, Harvey EN, Hobart G (1935) Potential Rhythms of the Cerebral Cortex during Sleep. Science 81(2111): 597-598.
-
Buzsáki G (1996) The hippocampo-neocortical dialogue. Cereb Cortex 6(2): 81-92.
-
Siapas AG, Wilson MA (1998) Coordinated interactions between hippocampal ripples and cortical spindles during slow-wave sleep. Neuron 21(5): 1123-1128.
-
Clemens Z, Molle M, Eross L, Barsi P, Halasz, et al. (2007) Temporal coupling of parahippocampal ripples, sleep spindles and slow oscillations in humans. Brain 130(11): 2868-2878.
-
Rosanova M, Ulrich D (2005) Pattern-specific associative long-term potentiation induced by a sleep spindle-related spike train. J Neurosci 25(41): 9398- 9405.
-
Scholle S, Zwacka G, Scholle H (2007) Sleep spindle evolution from infancy to adolescence. Clin Neurophysiol 118(7): 1525-1531.
-
Huupponen E, Himanen L, Varri A, Hasan J, Lehtokangas, et, al. (2002) A study on gender and age differences in sleep spindles. Neuropsychobiology 45: 99-105.
-
Shibagaki M, (2005) Delta and spindle components in the integrated EEG during nocturnal sleep for infants with developmental disabilities. Percept Mot Skills 101(3): 811-818.
-
Shibagaki M, Kiyono S, Watanabe K (1982) Spindle evolution in normal and mentally retarded children: a review. Sleep 5(1): 47-57.
-
Gais S, Molle M, Helms K, Born J (2002) Learning- dependent increases in sleep spindle density. J Neurosci 22(15): 6830-6834.
-
Peters KR, Ray L, Smith V, Smith C (2008) Changes in the density of stage 2 sleep spindles following motor learning in young and older adults. J Sleep Res 17(1): 23-33.
-
Maertelaer V, Hoffman G, Lemaire M, Mendlewicz J (1987) Sleep spindle activity changes in patients with affective disorders. Sleep 10(5): 443-451.
-
Lopez J, Hoffmann R, Armitage R, (2010) Reduced sleep spindle activity in early-onset and elevated risk for depression. J Am Acad ChildAdolesc Psychiatry 49(9): 934-943.
-
Goetz R, Goetz DM, Hanlon C, Davies M, Weitzman ED, et al. (1983) Spindle characteristics in prepubertal major depressives during an episode and after sustained recovery: a controlled study. Sleep 6(4): 369-375.
-
Reynolds CF, Kupfer DJ, Taska LS, Hoch CC, Spiker DG, etal. (1985) EEG sleep in elderly depressed, demented, and healthy subjects. Biol Psychiatry 20(4): 431-442.
-
Kaufman J, Birmaher B, Brent DA, Ryan ND, Rao U (2000). K-SADS-PL. J Am Acad Child Adolesc Psychiatry 39(10): 1208.
-
Poznanski EO, Freeman LN, Mokros HB (1985) Children's Depression Rating Scale - Revised (September 1984). Psychopharmacol Bull 21(4): 979- 989.
-
Armitage R, Emslie GJ, Hoffmann RF, Weinberg WA, Kowatch RA, et al. (2000) Ultradian rhythms and Nursing & Healthcare International Journal temporal coherence in sleep EEG in depressed children and adolescents. Biol Psychiatry 47(4): 338- 350.
-
Emslie GJ, Armitage R, Weinberg WA, Rush AJ, Mayes TL, et al. (2001) Sleep polysomnography as a predictor of recurrence in children and adolescents with major depressive disorder. Int J Neuropsychopharmacol 4(2): 159-168.
-
Robert JJ, Hoffmann RF, Emslie GJ, Hughes C, Rintelmann J, et al. (2006) Sex and age differences in sleep macroarchitecture in childhood and adolescent depression. Sleep 29(3): 351-358.
-
Shaffer D, Gould MS, Brasic J, Ambrosini P, Fisher P, et al. (1983) A children's global assessment scale (CGAS) Arch Gen Psychiatry 40(11): 1228-1231.
-
Mrazek DA, Mrazek P, Klinnert M (1995) Clinical assessment of parenting. J Am Acad Child Adolesc Psychiatry 34(3): 272-282.
-
Tanner JM (1962) Growth at Adolescence: Springfield: Charles C Thomas.
-
Rechtschaffen A, Kales A (1968) A Manual of Standarized Terminology, Techniques and Scoring Systems for Sleep Stages of Human Subjects. National Institute of Healthy Publication No. 204. US Goverment Printing Office, Washington DC.
-
Halbreich U, Lumley LA (1993) The multiple interactional biological processes that might lead to depression and gender differences in its appearance. J Affect Disord 29(2-3): 159-173.
-
Parry BL, Haynes P (2000) Mood disorders and the reproductive cycle. J Gend Specif Med 3(5): 53-58.
-
Krishnan V, Nestler EJ (2010) Linking molecules to mood: new insight into the biology of depression. Am J Psychiatry 167(11): 1305-1320.
-
Clemens Z, Molle M, Eross L, Jakus R, Rasonyi G, et al. (2011) Fine-tuned coupling between human parahippocampal ripples and sleep spindles. Eur J Neurosci 33(3): 511-520.
-
Campbell IG, Darchia N, Khaw WY, Higgins LM, Feinberg I (2005) Sleep EEG evidence of sex differences in adolescent brain maturation. Sleep 28(5): 637-643.
-
Zisook S, Rush AJ, Albala A, Alpert J, Balasubramani GK, et al. (2004) Factors that differentiate early vs. later onset of major depression disorder. Psychiatry Res 129(5): 127-140.
-
Dresler M, Kluge M, Genzel L, Schussler P, Steiger A (2010) Impaired off-line memory consolidation in depression. Eur Neuropsychopharmacol 20(8): 553- 561.
-
Fogel SM, Smith CT (2006) Learning-dependent changes in sleep spindles and Stage 2 sleep. J Sleep Res 15(3): 250-255.
-
Bertocci MA, Dahl RE, Williamson DE, Iosif AM, Birmaher B, et al. (2005) Subjective sleep complaints in pediatric depression: a controlled study and comparison with EEG measures of sleep and waking. J Am Acad Child Adolesc Psychiatry 44(11): 1158- 1166.
- Teaching Cognitive Behavioral Therapy to Graduate Psychiatric Mental Health Nurse Practitioner Students: Utilizing A Mixed Methods Course Evaluation
- The Role of Pharmacogenetic Testing in Clinical Practice: A Path toward more Effective, Personalized and Cost-effective Care
- Implementing Screening with the GAD-7 in an Outpatient Mental Health Setting: A Quality Improvement Project
- Social Media and Health Promotion
- Impact of Covid-19 Pandemic on the Academic Performance and Attitude of Nursing Students towards E-Learning
- A Study to Assess the Knowledge Regarding Ill Effects of Tobacco Consumption on Health among Adolescents in Selected Higher Secondary School in Nadiad City