Function of 5-Hydroxytryptamine Type 3 Receptors-A Computational Approach
This study investigates the function of 5-HT type 3 (5-HT3) receptors using a computational approach. Antagonists of the 5-HT3 receptor are currently one of the most effective therapeutic agents in treatment of chemotherapy-induced nausea, vomiting, and irritable bowel syndrome. Several experimental studies have shown the effect of pharmacological agents such as 9-tetrahydrocannabinol (THC), the psychoactive component of Cannabis, Cannabidiol (CBD), a non-psychoactive ingredient of Cannabis plant, Menthol, Propofol, and etc. on the functional human 5-HT3 receptors expressed in Xenopus oocyte as well as rat nodose ganglion neurons. 5-HT evoked currents recorded by a two-electrode clamp technique were inhibited by ligands in a concentration dependent manner. Simulations of allosteric inhibition were modeled using Vina docking techniques with the 5-HT3 structure. The 5-HT3 structure was found using homology sequence similarity techniques with the neuronal nicotinic acetylcholine receptor (nACH) and inhibitory neurotransmitter receptor for GABA (A). Results of studies with other members of the super family of ligand gated ion channels signified key residues involved in ligand binding sites within the transmembrane region of 5-HT3. Flexible and rigid docking simulations around key residues resulted in a number of low-energy (high affinity) configurations of ligand binding. The predicted residues TYR and THR may constitute a naturally occurring binding site for 5-HT3.
Introduction
Plants have long been used as a source of natural chemicals for medicinal products on a commercial scale. Menthol, a chemical constituent of the mint plant is used as a very important flavoring agent in products such as toothpaste, mouthwash, foods and cigarettes. Studies have shown that menthol containing products can be used for medicinal purposes ranging from pain management to nausea and vomiting treatment. Until recently, the cellular and molecular targets mediating beneficial effect of menthol containing products was unknown. Antagonists of the 5-HT3 receptor are currently one of the most effective therapeutic agents in treatment of chemotherapy induced nausea, vomiting, and irritable bowel syndrome [1]. The serotonin type three (5-HT3) receptor, a member of the cys-loop family of ligand-gated ion channels, mediates fast depolarizing actions of 5-HT in the central and peripheral nervous system [2]. Several experimental studies have shown the effect of pharmacological agents inhibit the membrane currents of 5-HT. The effects of 9-tetrahydrocannabinol (THC), the psychoactive component of cannabis, on the function of 5- HT type 3 (5-HT3) receptors were investigated using a two-electrode voltage clamp technique in Xenopus oocytes, and a whole-cell patch clamp technique in rat nodose ganglion neurons [3]. The results of this study indicate for the first time that, in Xenopus oocyte expression system, the inhibition by THC, a major psychoactive constituent of Cannabis sativa, of 5-HT3 receptors depends significantly on the amount of functional receptors expressed in this system. Also, the same laboratory investigated the effects of alcohol monoterpene menthol, a major active ingredient of the peppermint plant, on the function of human hydroxytryptamine type 3 (5-HT3) receptors expressed in Xenopus oocytes [4]. They found that 5-HT3 receptor mediated currents in acutely dissociated nodose ganglion neurons were also inhibited by menthol (100 μM). These data demonstrate that menthol, at pharmacologically relevant concentrations, is an allosteric inhibitor of 5-HT3 receptors. However, experimental studies don’t include the 5-HT3 receptor kinetics such as binding sites as well as binding energy in their electrophysiology data. Clearly, more detailed investigations in mammalian neurons or cell lines are needed, where the receptor kinetics in a more detailed and precise manner are included. In this computational study we try to investigate the function of 5-HT type 3 (5-HT3) receptors using same pharmaceutical agents in experimental data [5]. We simulate this experiment using docking of small ligands onto the 5-HT3 chain a receptor (Figure 1).
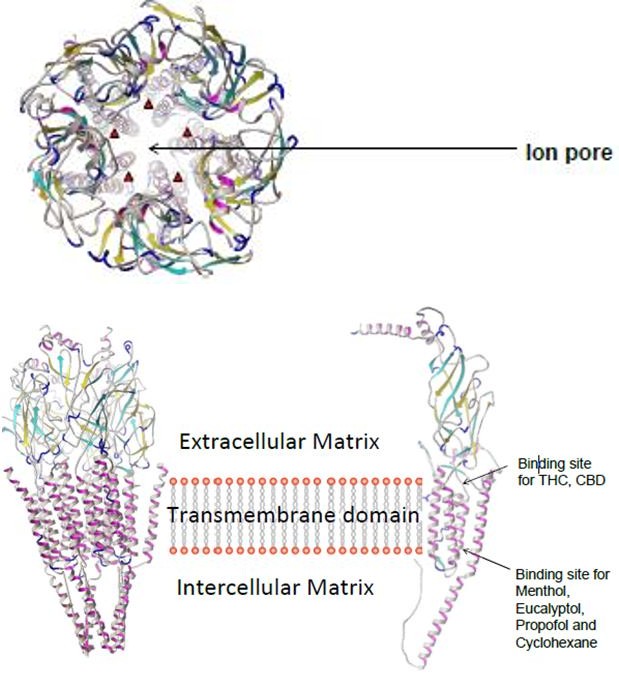
Docking predicts the position and orientation of a ligand when it is bound to a protein receptor or enzyme. A scoring function is used to calculate the energy of interaction based on electrostatics, Van der Waal’s interactions, hydrogen bonds, solvation effects, and loss of entropy.
Methods
Experimental Methodology
Synthesis of cRNA from cDNA clones of the human 5- HT3A, Radioligand Binding Studies, [35S] GTPgS Binding and Whole-Cell Patch-Clamp Recording were performed [4, 6].
Computational methodology
- Obtain a Structural Model of 5-HTR3A. We used Homology modeling techniques utilizing the PHYRE2 6 server to generate a structural model for 5-HTR3A. The result was a model for each chain of the pentameric structure of 5-HT3 [7]. Chain A of the nicotinic acetylcholine receptor (nAChR), under PDB code 2BG9 and the g-aminobutyric acid a receptor (GABAAR), PDB code 4COF was chosen as the modeling template for a Cys-loop receptor. The transmembrane domain of the modeled structure was employed for docking, as described below.
- Docking of Ligands on the 5-HTR3A Structural Model. We use Autodock Vina program to estimate binding energies and binding modes of ligands such as menthol, eucalyptol, propofol, cyclohexane, THC, and CBD. * and thymol and 2isopropylphenol* In order to use this docking program we have to define the region of the receptor related to a binding site. This region is identified based on experimental studies1,2,3. Autodock Vina performed rigid and flexible docking simulations. Choice of the specified grid box area took into account similar binding characteristics believed to be shared by propofol and menthol [8] and the close homology of the g- aminobutyric acid a receptor (GABAAR) to 5-HTR3A [1].
Flexible docking. A more sophisticated model of receptor allows close residues inside of binding site to adjust side chain conformations. This was performed on all large ligands (THC, CBD) Open macromolecule file with Harlem and assign binding site. Find center of bind site and measure a sphere 15A away from center. Color and note each amino acid that lies within the measured area. Assign amino acid residues within site for flexible binding techniques in autodock. • Analyze Interactions. Each ligand was modeled and interactions were visualized using Autodock Tools. First Open ‘Autodock tools’ > click ‘AutoDock vina Result’ Locate receptor and add as macromolecule + select ‘vina result’ Macromolecule – 5HT3.pdbqt, vina result (ligand) – Propofol.pdbqt Click ‘Analyze Results’ and select ‘Single molecule with multiple conformations’ click ‘OK.’ The docked molecule has now displayed its lowest energy conformation. Keyboard left and right to browse other vina docking conformations lowest to highest. (9 total – all zoomed out w/out interactions with amino acid chain of 5HT3) Now click ‘Analyze’ > ‘Dockings’ > ‘Show Interactions’ (This changes the display to a close up image of the first lowest conformation of the docked ligand.) The background color was set to white. (Customizable within display widget) Click ‘Show Molecular Surface’ for ligand and color by atom type to find ligand atoms close to receptor atoms and white for atoms not close to any receptor atoms. Click ‘Create Spheres’ to show wire spheres around receptor atoms close to a ligand. (color gray) Add labels and sticks for residues interacting close to ligand. (optional: choose within 5-10A) Click ‘Hydrogen Bonding’ – choose ‘Activate as Spheres’ (color green – default) and adjust radii + spacing for optimum view. Toggle and zoom to get preferred angle that clearly captures ligand atom bonding with amino acid residues or hydrogen bonds, or both. Capture image by clicking ‘Set Antialiasing’ which will increase quality of viewing window and click save image. Output list of ligand contacts, receptor contacts and hydrogen bonds.
Results
Experimental Results
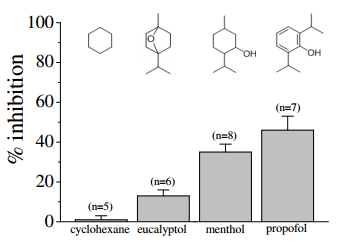
Comparison of the effects of equimolar concentrations (100 µM) of cyclohexane, eucalyptol, propofol, and menthol on 5-HT3 receptors. Bars represent the means ± S.E.M (n=5-8). Chemical structures of the compounds were presented at the top of each bar [4].
Computational Results
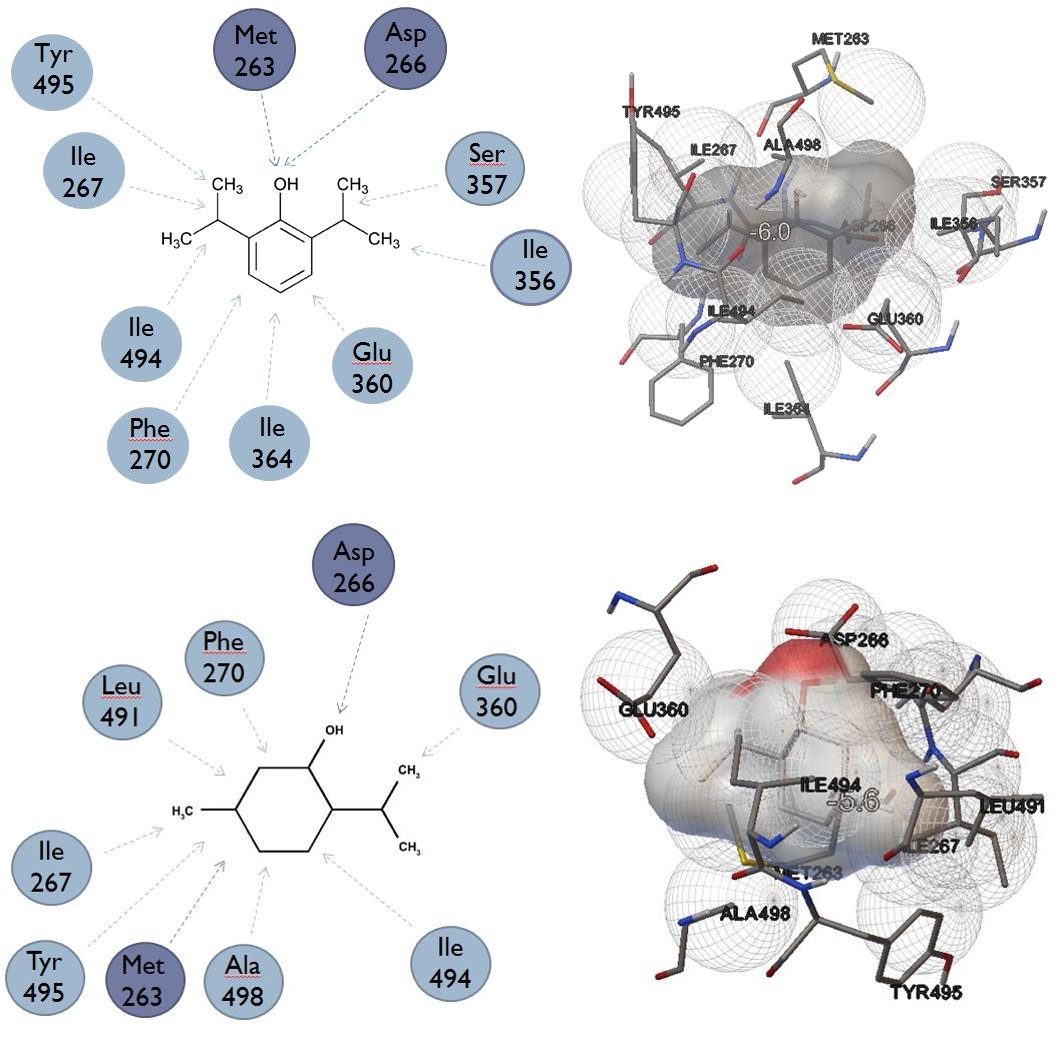
A. Results of rigid docking to lower binding site next to Threonine 361 (menthol binding site, lowest energy conformation) [7].
| Compound | Binding energy (kcal/mol) | ||||
|---|---|---|---|---|---|
| Propofol | -6 | ||||
| Menthol | -5.6 | ||||
| Eucalyptol | -4.5 | ||||
| Cyclohexane | -4.5 | ||||
| Compound | Binding energy (kcal/mol) | ||||
| Thymol | -6.4 | ||||
| 2-isopropylphenol | -6.1 | ||||
| Benzene | -6 |
| Compound | 2-D Ligand interaction diagram | 3-D depiction of interaction |
|---|---|---|
| Propofol | ||
| Menthol | ||
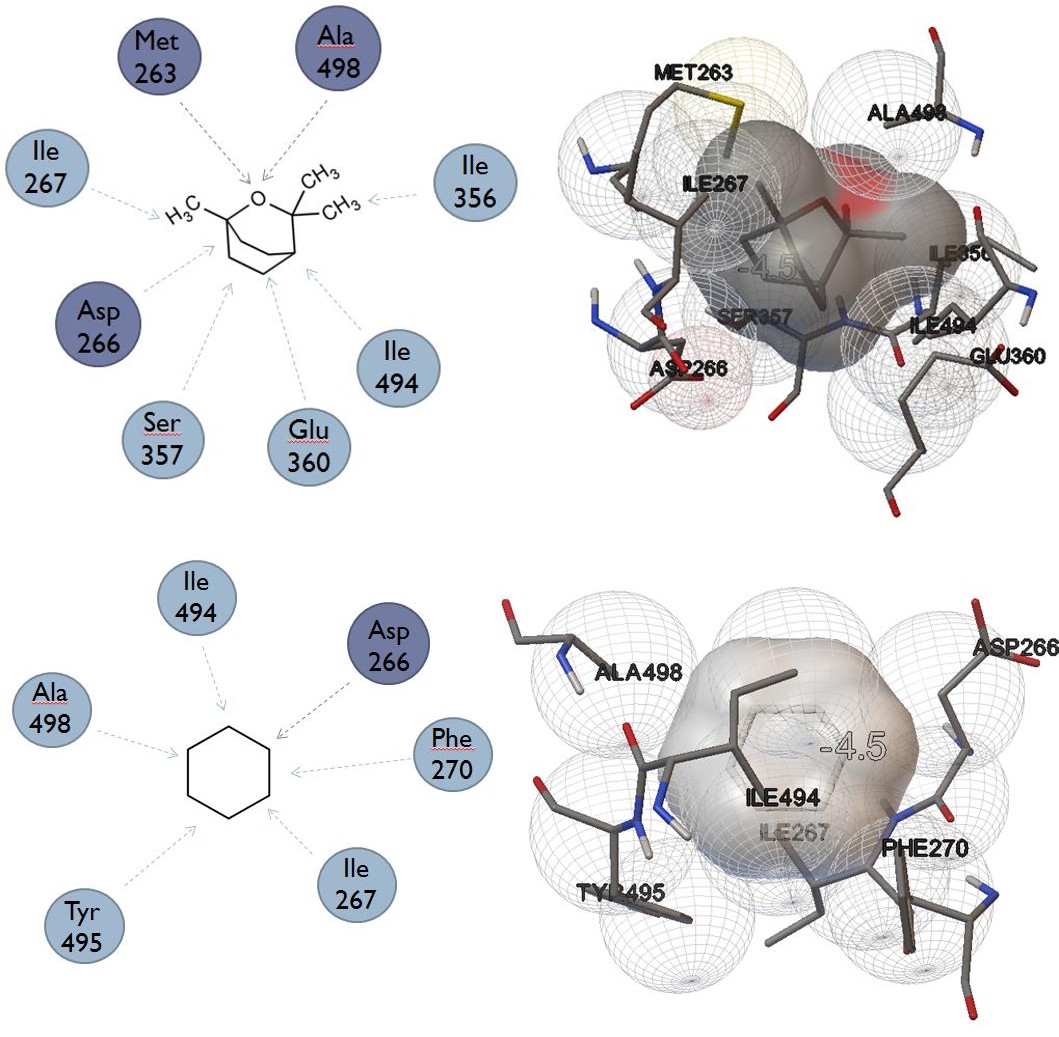
| Eucalyptol | |
|---|---|
| Cyclohexane |
B. Results of rigid and flexible docking to upper binding site near Tyrosine 346 (THC and CBD binding site, lowest energy conformation).
The following are readout files for the rigid docking of THC and CBD. The docked molecule has now displayed its lowest energy conformation.
################################################################# # If you used AutoDock Vina in your work, please cite: # # # # O. Trott, A. J. Olson, # # AutoDock Vina: improving the speed and accuracy of docking # # with a new scoring function, efficient optimization and # # multithreading, Journal of Computational Chemistry 31 (2010) # # 455-461 # # # # DOI 10.1002/jcc.21334 # # # # Please see http://vina.scripps.edu for more information. # ################################################################# Output will be thc_out.pdbqt Detected 4 CPUs Reading input ... done. Setting up the scoring function ... done. Analyzing the binding site ... done. Using random seed: 1209244732 Performing search ... done. Refining results ... done.
mode | affinity | dist from best mode |(kcal/mol) | rmsd l.b.| rmsd u.b. -----+------------+----------+----------
1 -8.4 0.000 0.000 2 -8.4 1.768 6.373 3 -8.3 1.994 2.499 4 -8.2 1.816 6.953 5 -8.0 1.698 2.440 6 -7.7 1.958 7.144 7 -7.7 1.816 6.167 8 -7.6 1.960 6.766 9 -7.6 1.946 2.376 Writing output ... done.
################################################################# # If you used AutoDock Vina in your work, please cite: # # # # O. Trott, A. J. Olson, # # AutoDock Vina: improving the speed and accuracy of docking # # with a new scoring function, efficient optimization and # # multithreading, Journal of Computational Chemistry 31 (2010) # # 455-461 # # # # DOI 10.1002/jcc.21334 # # # # Please see http://vina.scripps.edu for more information. # #################################################################
Output will be cbd_out.pdbqt Detected 4 CPUs Reading input ... done. Setting up the scoring function ... done. Analyzing the binding site ... done. Using random seed: 1410998816 Performing search ... done. Refining results ... done.
mode | affinity | dist from best mode |(kcal/mol) | rmsd l.b.| rmsd u.b. -----+------------+----------+----------
1 -7.7 0.000 0.000 2 -7.5 0.223 1.863 3 -7.2 2.642 5.157 4 -7.2 2.510 5.303 5 -6.2 6.175 12.965 6 -6.1 1.014 2.944 7 -6.0 2.121 5.743 8 -6.0 2.155 5.161 9 -5.5 5.423 11.766 Writing output ... done.
C. Flexible Docking
251, LEU 255, LEU 301, THR 304, VAL 345, CYS 350 and LEU 509. Rigid and flexible docking results for THC and CBD and their corresponding lowest energy conformation displayed in 3D below.
Flexible docking between Delta-9- Tetrahydrocannabinol (THC), Cannabidiol (CBD) and residue receptor (5-HT3) – flexible residues include TYR
| Compound | Rigid docking – Binding energy (kcal/mol) | Flexible docking – Binding Energy (kcal/mol) |
|---|---|---|
| Delta-9-Tetrahydrocannabinol | -8.4 | -9.2 |
| Cannabidiol | -7.0 | -8.8 |
| THC = -9.2 kcal/mol | CBD = -8.8 kcal/mol |
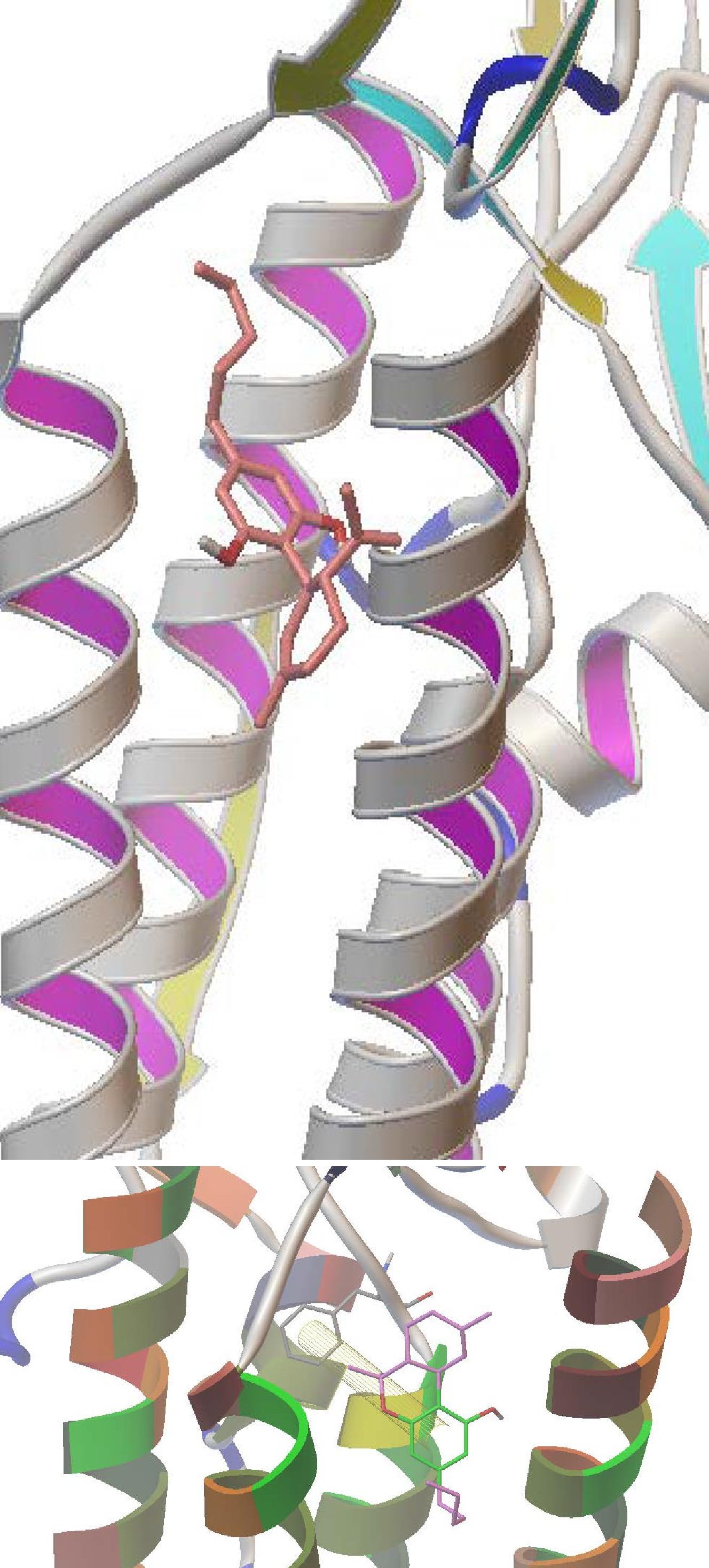
Pi-Pi bonding interactions between Delta-9-Tetrahydrocannabinol (Ligand) and residue PHE 347 receptor (5-HT3) - attractive, noncovalent interactions between aromatic rings. (Shown below)
Discussion
Structural transition causing a conformational change occurs as a result of ligand binding, known as allosteric transition [9]. Antagonist such as THC, CBD, and propofol bind to different sites within the transmembrane domain. Binding of antagonists can create necessary driving forces for conformational transition in such a way that the closed channel state becomes more favorable. A strong binding affinity of antagonists causes more driving force for the receptor to stay in the closed state. Results of the lower binding site docking corresponds to the experimental finding that propofol is the strongest inhibitor of 5-HT3 receptor membrane currents (~50%) while expressing the largest binding energy of -6.0 kcal/mol. The trend of absolute values of calculated binding energies for the other compounds (propofol>menthol> eucalyptol>cyclohexane) provide rational to experimental findings of difference in inhibition of 5HT3 by the same compounds [10]. Both THC and CBD strongly and selectively bind to the upper site (TYR 346). Similar binding energies for THC and CBD explain their similar inhibition of membrane currents in 5HT3 [11]. In order to verify the predicted binding mode, we proposed experimental studies of site-directed mutagenesis of the residue involved in both hydrophilic and hydrophobic interactions at the THR 361 binding site. Experiments using thymol (-6.4 kcal/mol) and 2- isopropylphenol (-6.1 kcal/mol) will be done after docking simulations showed that both interact stronger than propofol and menthol with the THR 361 binding site. Thymol, a monoterpenoid similar to menthol, has an aromatic group which is responsible for its higher affinity to the lower binding site (THR361).
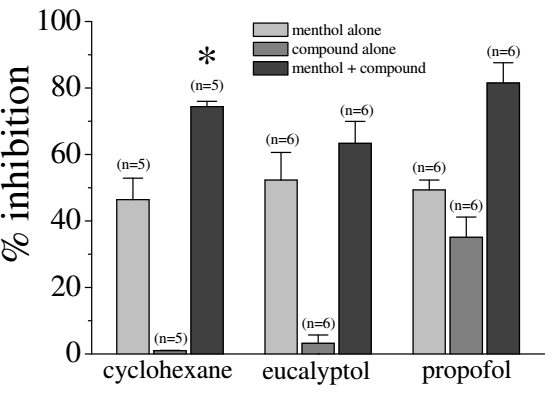
Evaluation of the data in the figure on the left provided evidence that combining menthol with either eucalyptol or propofol enhanced its inhibitory effect on 5HT-induced currents, suggesting that these compounds have additive effects on 5-HT3 receptors [4]. On the other hand, cyclohexane (100 mM) alone displayed undetectable efficacy at inhibiting 5-HT3A receptors [12], but increased significantly the potency of menthol, suggesting that cyclohexane may facilitate the interaction of menthol with its binding site on the receptor. Our computational findings did not back up the observed increase in inhibition of menthol when in the presence of cyclohexane, as shown in the figure above. Experimental error occurred while simultaneously docking two ligands in the lower active site (THR 361) to recreate these results. The initial process of docking menthol into the active site, saving it as a functional (flexible) part of the receptor, and docking the second ligand, cyclohexane, to the system of ligand and receptor, was proven ineffective. Cyclohexane, in the presence of menthol, causes a vast increase in inhibition than the compounds alone. This led to the theory that cyclohexane doesn’t pass into the transmembrane region, but sits on the outside of the channel affecting its membrane potential. The increase shown in the figure above could be attributed to the change in the potential energy across the membrane that cyclohexane could cause.
Conclusion
Experimental inhibition of homomerically expressed human 5-HT3 receptor shows similar trend in computational binding energies to the binding site located near THR 361. Docking calculations provide explanation of molecular basis of difference in inhibition by menthol like compounds. Similar binding energies for THC and CBD corresponds to their similar inhibition of membrane currents measured in experiment [3].These results provide rational for search of new compounds that might be used as antiemetic drugs.
References
-
Thompson AJ, Lester HA, Lummis SC (2010) The structural basis of function in Cys-loop receptors. Q Rev Biophys 43(1): 449-499.
-
Yakel JL, Meyer B Jckson (1988) 5-HT3 receptors mediate rapid responses in cultured hippocampus and a clonal cell line. Neuron 1: 615-621.
-
Yang KH, Isaev D, Morales M, Petroianu G, Galadari S, et al. (2010) The effect of Δ9-tetrahydrocannabinol on 5-HT3 receptors depends on the current density. Neuroscience 171(1): 40-49.
-
Ashoor A, Nordman JC, Veltri D, Yang KH, Shuba Y, et al. (2013) Menthol inhibits 5-HT3 receptor-mediated currents. J Pharmacol Exp Ther jpet 347(2): 398-409.
-
Yip MS, Zi-Wei Chen, Christopher J Edge, Edward H Smith, Robert Dinckinson (2013) A propofol binding site on mammalian GABA-A receptors identified by photolabeling. Nat Chem Biol 9: 715-720.
-
Kelley LA, Sternberg MJ (2009) Protein structure prediction on the web: a case study using the Phyre server. Nature Protocols 4(3): 363-371.
-
Trott O, Olson AJ (2010) Autodock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization and multithreading. Journal of Computational Chemistry 31(2): 455-461.
-
Hall AC, Turcotte CM, Betts BA, Yeung WY, Agyeman AS, et al. (2004) Modulation of human GABAA and glycine receptor currents by menthol and related monoterpenoids. Eur J Pharmacol 506(1): 9-16.
-
Jackson M (2002) Allosteric mechanisms in the activation of ligand-gated channels. In: MD (edn) Biophysics Textbook Online, Vol. Channels, Receptors and Transporters. DE FELICE L, Biophysical Society, Bethesda, PP 1-48.
-
Williams DB, Myles H Akabas (2002) Structural Evidence that Propofol Stabilizes Different GABAA Receptor States at Potentiating and Activating Concentrations. Neuroscience 22(17): 7417-7424.4
-
Barann M, Dilger JP, Bonisch H, Gothert M, Dybek A (2000) Inhibition of 5-HT3 receptors by propofol: equilibrium and kinetic measurements. Neuropharmacology 39(6): 1064-1074.
-
Yang KH, Sehamuddin Galadari, Dmytro Isaev, Georg Petroianu, Toni S Shippenberg (2010b) The nonpsychoactive cannabinoid cannabidiol inhibits 5- hydroxytryptamine3A receptormediated currents in Xenopus laevis oocytes. J Pharmacol Exp Ther 333(2): 547-554.
- A Review of Gene Therapy for Parkinson's Disease to Control Dopaminergic Neurons
- Late-Onset Myasthenia Gravis in a Patient with Recurrent Breast Cancer: A Case Report
- Covid-Induced Dystonia and Opsoclonus: A Case Report
- Generalized Tonic-Clonic Seizure in a Pediatric Patient with Sunflower Syndrome: A Case Report
- Comparison of Doppler Guided Seldinger Technique Versus Classic Palpatory Seldinger Technique for Radial Artery Cannulation-an Open Label Randomized Controlled Trial
- Brown Sequard Syndrome: Understanding the Complexities of Spinal Cord Injury