An update on Telomeres Dynamics in Reproduction; Specifically in Females & Ageing: A Narrative Review
The oocyte portrays a postmitotic cell having a longer life working as locus of reproductive ageing in women. Female germ cells replication occurs in fetal life which age right through reproductive period. Modes behind oocyte ageing are oxidative injury, mitochondrial impairment and disturbed protein inclusive of cohesion. BobEdwards invented the “generation line” at the time of oogonial proliferation in mouse oogenesis, where the initial oocytes to leave the oogonial proliferation while fetal life are the ones which undergo ovulation first at puberty and the last oocytes to leave the oogonial proliferation are the last to undergo ovulation in adult. Hence Kalmbach K group posited ‘’second hit’’ hypothesis of reproductive ageing for incorporating to plethora of characteristics of the oocyte ageing. The 1st hit being oocyte continuing in the older women pass via more cell cycles while fetal oogenesis. The 2nd hit was oocytes which continued had>accrual of environmental & endogenous oxidative injury right through woman’s life. Telomeres (T’s) might modulate both parts of oocyte ageing. T’s offer a mitotic clock, with T debilitation becoming a granted inimical sequelae of cell division due to end replication cell botheration. The guanine enrichment of the telomeres makes them more vulnerable to oxidative injury, in post mitotic cell also. Since telomerase,/ the reverse transcriptase is more adept in sustenance rather than T lengthening. Furthermore telomerase continues to be inactive in maximum oogenesis & early generation. Thus oocytes possess small T’s on the border of viability. To corroborate this posit mice having induced T debilitation and women having natural telomeropathies have diminished ovarian reserve (DOR), aberrant embryonic generation & infertility. Conversely sperms form all through life of male by ALT, have telomerase active progenitor, spermatogonia, longest body T’s. Numerous queries arise i) does normal T debilitation of human oocytes aid in meiotic nondisjunction (MN)? ii) does recombination dependent T lengthening make embryos more prone to MN / mosaicism? iii) can certain features of T’s work as oocyte quality markers.
Introduction
Telomere Biology
In case of maximum metazoans telomere (Ts) are comprised of the tandem repeated nucleotide sequences (TTAGGG) as well as correlated proteins which generate caps conferring protection to the chromosome end [1]. Chromosomal DNA ends in case of single stranded 3’ overhang which loops back over itself, generating an area referred to telomeric loop (T loop) for masking the single stranded 3’ DNA loop for the avoidance of the DNA damage response (DDR) [2]. The displacement loop (Dloop) nested (loop in loop) within the T loop generates once the 3’ overhang infiltrates the double-stranded telomeric DNA [2]. Telomeric repeats in the form of the substrates for the shelterin, a protein complex implicated in the looping telomeric DNA. Telomeres confer protection to the chromosome end from the DDRs in addition to determination in the form of the single or double-stranded breaks by the cells [3]. The commonest DDR pathways are non homologous end joining (NHEJ), which leads to T fusions in addition to impairment [4]. Ataxia telangiectasia mutated (ATM) results in activation in reactions to the double-stranded breaks by the mobilization of the phosphatidyl inositol 3 – kinase (PI3K) to the injury [5]. Ataxia telangiectasia and Rad3-related (ATR) the reactions to the single stranded breaks by enrollment of the replication protein A (RPA); that coats the single stranded DNA [6]; along with homologous recombination repair (HRR) leads to healing of the double-stranded breaks by resection, invasion of the strand, as well as the generation of the new DNA by utilization of homologous sequences in the form of template [6, 7, 8] (Figure 1).
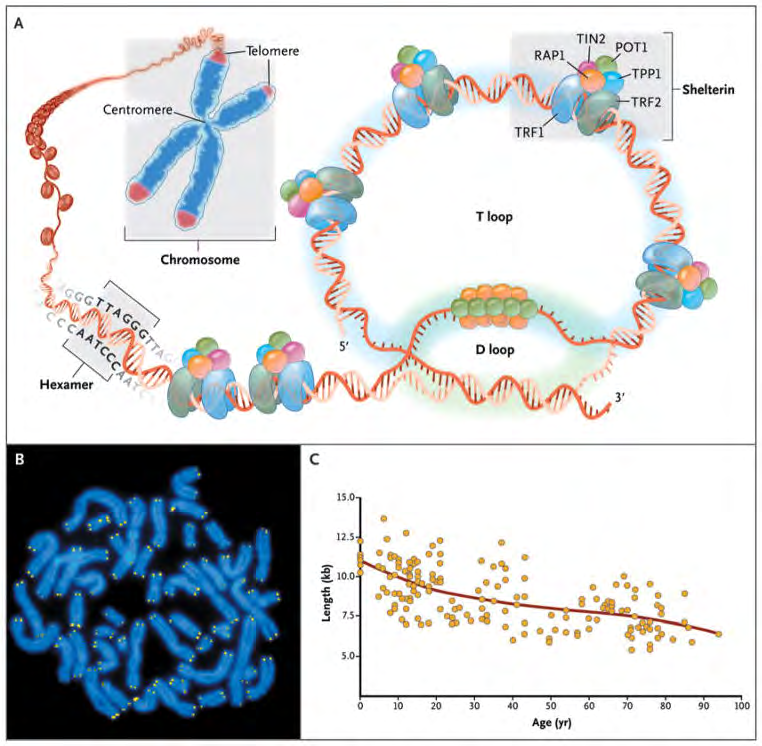
Figure1: Telomere Structure.
Courtesy ref no-8-As shown in Panel A, telomeres are located at the ends of linear chromosomes; they are composed of hundreds to thousands of tandem DNA repeat sequences: hexameric TTAGGG in the leading strand and CCCTAA in the lagging strand in humans. Protective proteins associated with telomere DNA are collectively termed shelterin (TRF1, TRF2, TIN2, POT1, TPP1, and RAP1). The 3′ end of the telomeric leading strand terminates as a single- stranded overhang, which folds back and invades the double- stranded telomeric helix, forming the T loop. As shown in Panel B, telomeres can be directly visualized under the microscope at the ends of metaphase chromosomes (four telomere signals per chromosome) by fluorescence in situ hybridization (FISH). (Image provided by Peter Lansdorp, M.D., Ph.D.) Average telomere length can be measured by several methods: a technique that combines flow cytometry and FISH (flow-FISH), Southern blotting, and a quantitative polymerase-chain-reaction (qPCR) assay. Flow-FISH can measure the telomere length in different cell subgroups, such as granulocytes or CD4+ T lymphocytes; Southern blotting reveals length and length heterogeneity; and qPCR is a rapid assay that requires very small amounts of DNA. As shown in Panel C, the average length of telomeres in human leukocytes varies, ranging from approximately 11 kb at birth (in umbilical-cord blood) to 6 kb at 90 years of age. Telomere loss is most rapid early in life, and over a life span it is not linear but follows a third-order polynomial.
Shelterin, portrays an imperative part of the T cells structure as well as sustenance in the cells. In case of humans; shelterin, gets constituted of 6 proteins: TRF1,TRF2,POT1,TPP1,TIN2 along with RAP1. These crosstalk with the formation of the T loop in addition to cause suppression of the DNA healing modes which in case of alternate conditions would be activating injury reactions to the uncapped chromosomes [3, 9]. TRF1, as well as TRF2 generate a duplex for the binding of the double-stranded T DNA [10]. Telomeric repeat binding factor 1- interacting protein 2 binds to the TRF1, as well as TRF2 scaffold along with correlates with POT1, commonly complexing with TPP1 [11], TRF2 further enrols with RAP1 to the Ts [12]. POT1 binding takes place to the single stranded telomeric DNA in addition to TPP1along with TIN2 suppress ATR [13]. POT1 coats T overhangs as well as represses ATR [14]. Hampering of the POT1 stimulates T impairment stimulated foci marked by H2AX, pointing that ATM is suppressed basically through TRF2 [13, 15]. TRF2 in addition to RAP1 repress NHEJ [16, 17] (Figure 2).
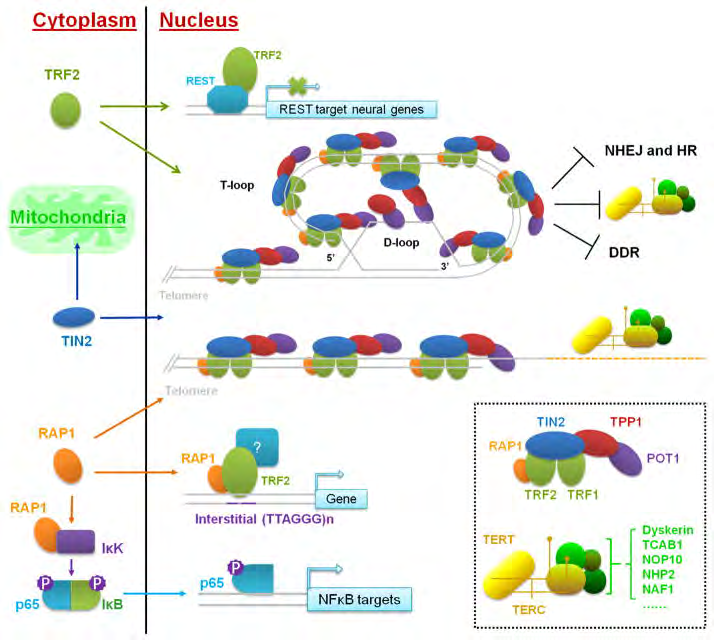
Figure 2: Courtesy ref no-17-Canonical and extra-telomeric function of telomere-binding proteins. The six-protein telosome complex and telomerase holoenzyme are depicted in small box. In the nucleus, the telosme and telomerase complex maintain telomere length and protect telomere ends. Several telomere proteins also participate in non-telomeric pathways in the cytoplasm.
Earlier we had highlighted role of telomeres while editing topics in reproductive health [18]. Here we tried to detail the telomere dynamics in human reproduction.
Methods: Here we conducted a systematic review utilizing search engine pubmed; google scholar; web of science; embase; Cochrane review library utilizing the MeSH terms like; telomeres; homologous end joining (NHEJ); DNA damage respone (DDR); Ataxia telangiectasia mutated (ATM); Shelterin; TRF1; TRF2; POT1; TPP1; TIN2; G-Quadruplexes; T Lengthening; T shortening; Cigarette smoking; psychological stress; guanine enrichment; telomerase; alternative lengthening of telomeres (ALT); telomeropathies; CD from 1970’s to March 2024.
Results: We found a total of 2000 articles out of which we selected 87 articles for this review. No meta-analysis was done.
Role of G- Quadruplexes
In human cells, the main protective single-stranded binding protein at telomeric overhangs is POT1 [19]. The
potential generation of G-quadruplexes at overhangs would thereby be in competition with POT1 binding, actually POT1 has been illustrated to unwind intramolecular G-quadruplexes with the aim of binding to telomeric DNA [19]. POT1 unwinds G-quadruplexes via conformational selection mode in which G-quadruplex unwinding takes place before POT1 binding [19]; subsequent to that every OB folds of two POT1 molecules binds to one telomeric repeat of the 4-repeat G-quadruplex in a stepwise manner (Figure 3) [19]. Another G4-unwinding protein that is acknowledged to localize to telomeres is replication protein A (RPA) [19], which possesses a main part in activating the DNA damage response to single-stranded DNA [19]. If human telomeric oligonucleotides are folded into G-quadruplexes, the capability of POT1 to trap them once they unfold out- competes the capacity of RPA to unfold the G-quadruplex [19], revealing the protective function of POT1 at telomeres.
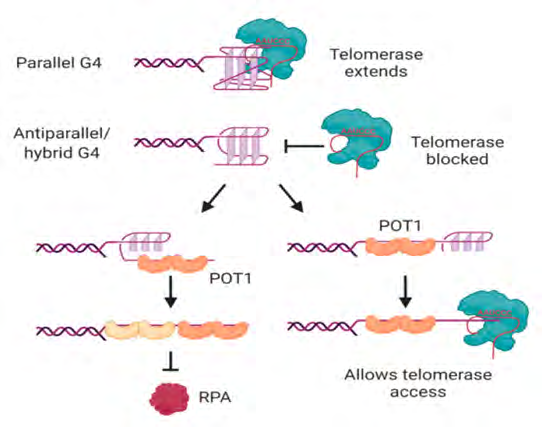
Figure 3: Courtesy ref no-19-Interactions of telomerase, POT1 and RPA with human telomeric G-quadruplexes. Telomerase can bind and extend parallel, but not antiparallel or hybrid, G-quadruplexes. POT1 binds to antiparallel or hybrid G-quadruplexes through a mechanism in which G4 unfolding precedes “trapping” of the unfolded DNA by POT1. The two OB folds of each POT1 molecule bind to consensus binding site TTAGGGTTAG; sequential binding of two POT1 molecules therefore coats the 4-repeat telomeric DNA (left). Although RPA also has the ability to unwind G-quadruplexes, POT1 competes with this activity. If binding of POT1 occurs at the 5′ region of the DNA, the 3′ tail can form a substrate for telomerase (right). Not shown is POT1’s binding partner TPP1, which also influences G4 unwinding dynamics and telomerase activity. See text for details and references. Figure created with BioRender.com.
POT1-modulated G-quadruplex unfolding further influence the capacity of telomeric sequences to be extended by telomerase, minimallyin vitro [19]. It was first displayed using telomerase from three separate species of ciliated protozoa that oligonucleotides folded into antiparallel intramolecular G-quadruplexes [19] do not generate good substrates for telomerase [19]. This is further the case for human telomerase; the antiparallel as well as hybrid G-quadruplexes developed by a 4-repeat oligonucleotide in K+ solution [19] are poor substrates for telomerase [19]. Binding of telomerase to the DNA is by these conformations of G-quadruplex [19], is hampered along with the attempted extension of the DNA by telomerase is non-processive [19]. POT1 possess the capacity of restoration of the capacity of telomerase to access these substrates by unfolding the G-quadruplex in addition to trapping the DNA in a linear form with a protruding tail (Figure 3) [19]. The dynamic folding of G-quadruplexes their unfolding by POT1 might thus work in the form of a controlling, regulatory reach of telomerase to the telomere.
Factors Causing T Shortening
Despite, the lack of the DDR’s shortening of T’s takes place with the every cycle of DNA replication [3, 20]. Minimal quantities of the DNA separate from the every chromosome end in view of the end replication botherations ‘’ which leads to the incapacity of DNA polymerase to totally replicate the strands that are lagging at the chromosome end [21]. Sustenance of a minimal range takes place in humans varying from the 5-15 kilobase (kb) whose shortening takes place at the rate of the 50-150 base pairs/cell division [22]. Furthermore shortening of T’s takes place in nondividing cells once exposed to the endogenous in addition to environmental factors for instance reactive oxygen species (ROS), cigarette smoking, toxins as well as chronic psychological stress [23, 24, 25, 26]. The guanine enrichment of the telomeres works in the form of a sink with regards to ROS along with shortening of T’s takes place from the DNA injury. These modes might aid in the reasoning out the association amongst stress, inflammatory situations, in addition to T debilitation [24, 27].
Cigarette smoking is one of the maximum evaluated environmental factors which aid in T debilitation. In case of the oocyte shortening of T’s takes place, escalating meiotic mistakes, diminishes fertilization, causes enhancement of aneuploidy as well as results in DNA injury. In case of mouse embryonic stem cells, cigarette smoking escalates apoptosis [25]. Exercise which is ultraendurance has been correlated with the lengthier leukocyte T in contrast to healthy age matched controls [27]. Psychological stress has been correlated with shorter leukocyte T lengthier in case of women, resulting in exacerbation of the rate of shortening which is in agreement with the 10yrs of normal ageing [24]. Robust maltreatment at the time of childhood along with racial differentiation further leads to augmentation of T shortening [26].
Lastly, T debilitation basically results in chromosome uncapping, that initiates apoptosis as well as Senescence, 2 of the modes behind the avoidance of the progression of the genomic instability of the cells [4, 6, 28]. Replication of the chromosomes which are uncapped results in chromosomal breakdown- fusions-breakage cycles leading to duplications, deletions mutations as well as translocations. Propagation of replication - based in addition to independent T debilitation/ wearing down has resulted in broad acceptability of the posit that T’s work in the form of the primary guide of the cellular ageing [29, 30].
Modes behind the T Lengthening
Lengthening of Telomeres
Takes place by 2 modes i) polymerase the reverse transcriptase ii) telomerase or iii) alternate lengthening of telomeres (ALT) [31, 32]. Telomeres possess the capacity of addition of the telomeric repeats at the rate of 50-100 base pairs /cell division, pointing that telomeres are basically implicated in the sustenance of the T length [33]. The enzyme telomerase is constituted by, telomerase reverse transcriptase (TERT), telomerase RNA constituents (TERC), in addition to the protein dyskerin [34], TERT work in the form of a RNA template read with the utilization of the TERT catalytic subunit for transcribing new T DNA. The part of dyskerin is in the stabilization of the complex apart from is implicated in the placement of the telomeres to the proper regions (Figure 4).
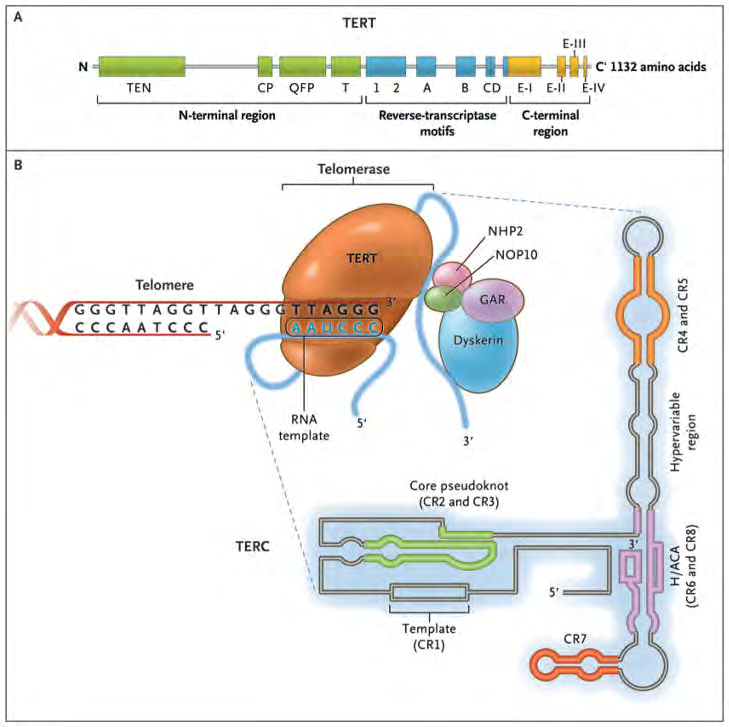
Courtesy ref no-8-The enzyme telomerase reverse transcriptase (TERT), its RNA component (TERC), the protein dyskerin, and other associated proteins (NHP2, NOP10, and GAR1) are shown. Telomerase catalytically adds TTAGGG hexameric nucleotide repeats to the 3′-hydroxyl end of the telomeric leading strand, using a specific sequence in the RNA component as the template. TERT contains three major domains: the N-terminal region, the reverse-transcriptase motifs, and the C-terminal region, all containing evolutionarily conserved motifs. TERC contains 451 nucleotides in seven conserved regions (CR1 through CR7), including the template (CR1), and an H/ACA box, a hairpin nucleotide sequence characteristic of a class of small nucleolar RNAs involved in RNA processing.
Abnormalities in the telomerase aid in the T debilitation/ wearing down [31, 35]. Mutations in the genes encoding constituents of the telomerase (TERT, TERC in addition to dyskerin), or shelterin lead to telomeropathies for instance dyskeratosis congenita (DC) [36], subsequent to the clinical triad of skin along with the nail aberrations in addition to oral leukoplakia [37]. Bone marrow failure, esesophageal stenosis, pulmonary fibrosis, liver impairment, bone aberrations, a susceptibility to cancer in addition to reproductive impairment are the characteristics by which the presentation of the patients with DC takes place [37, 38]. The (Figure 5) displays modes by which T shortening takes place [39].
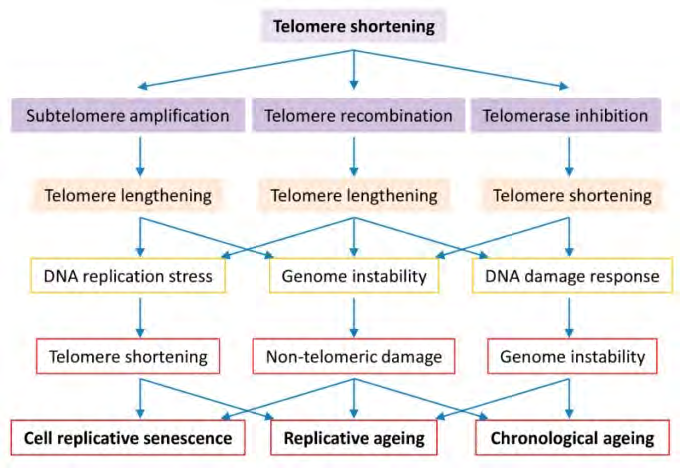
Figure 5: Courtesy ref no-39-Roles of telomere biology in cell senescence, replicative and chronological ageing. Cells evolve to have regulated telomerase activity to preserve telomere homeostasis which is vital to genome stability in organism ageing. Short telomeres are maintained by telomerase in the early stage, telomere DNA homologous recombination or subtelomeric DNA amplification in the late stage of telomere shortening. Telomerase inactivation results in critically short telomeres that either activate the cell cycle check-point resulting in cell senescence or promote telomere-telomere recombination or subtelomere amplification resulting in genome instability and replicative difficulty. Long telomere associated genome instability and replicative difficulty cause not only telomere shortening but also non-telomeric damages, culminating in cell senescence, replicative ageing and accelerated chronological ageing.
Shortening of T’s takes place in patients with DC, correlated with dysfunctional fertility, premature ovarian failure. DC gets inherited in the form of an autosomal dominant (AD), autosomal recessive (AR) or X linked fashion. Mutations of the TERT in addition to TERC genes lead to both AD as well as AR DC [40]. Mutations of the other genes inclusive of TCAB1, TIN2, CTC1 along with the RTEL1 result in DC by resulting in disturbance of the T structure or trafficking (alias interacting). Patients, that are heterozygous for TCAB1 mutations possess normal T length, however dysfunctional enrollment of telomerase to the Cajal bodies that contributes in the assembling of the telomerase as well as enrollment to the T’s [41]. Mutations of the TIN2 gene interfere with the generation of the T loop in addition to how telomerase can reach T’s [42]. CTC1 gene mutation leads to shortening of T’s by escalating breakdown [43]. The RTEL1 gene mutations further generate AD as well as AR DC [44].
Lengthening of Telomeres
Telomeres further possess the capacity of lengthening taking place independent of telomerase through a recombination dependent modes known as ALT [25]. This alternate kind of lengthening of the T’s enlarges Ts by HR of the sister chromatids or Ts from the other chromosomes. About 5-10% of cancers have sustenance of the T’s through ALT. This alternate kind of lengthening of the T’s implicates HR replication with the invasion of strand from one T to the other [45], or by rolling of T circle replication once T’s utilized circular loops in the form of templates meant for replication with the subsequent HR modulated lengthening [46]. Exchange of telomeres sister chromatids, at the time of exchange of Trepeats, further lengthens Ts which aids in T length heterogeneity [47]. This alternate kind of lengthening of the T’s generates an enrichment of extra chromosomal T repeats [48]. The instability correlated with recombination amongst chromatids aids in greater quantities of the genomic instability in case of ALT [7].
This alternate kind of lengthening of the T’s varies from the telomerase modulated lengthening of the T’s by probably leading to the adding of kbs of DNA instead of little base pairs lengthening generated by the telomerase. Furthermore, ALT guides T length heterogeneity in cells in view of the event does not work on T’s ubiquitously. Telomerase modulated lengthening leads to addition of only 50-100 T bp repeats/cell cycle as well as possesses the preference for the lengthening of the shortest Ts. This alternate kind of lengthening is responsible for the exchange of telomeres sister chromatids along with form extra chromosomal T repeats inclusive of T circle, as well as promyelocytic leukaemia nuclear bodies or ALT- correlated promyelocytic leukaemia nuclear bodies [49].
Determination of Ts: Numerous assays have been generated for the determination of various kinds of Tcell biology inclusive of average in addition to shortest T length. The assays utilized most frequently in reproductive endocrinology as well as reproductive medicine are inclusive of quantitative fluorescent in situ hybridization (qFISH) in addition to quantitative real-time polymerase chain reaction (qRT-PCR), iii) terminal restriction fragment analysis as well asiv) Single telomere length analysis (STELA). Application of telomere length assays was initially performed in a population of cells for instance leukocytes or endometrium. Wang F, et al. [50], inclusive of others performed modifications of the assays for the determination of T’s in single cells for instance oocyte, sperm, polar bodies, ortrophectoderm [50]. This single cell evaluation illustrated considerable heterogeneity of telomere length in numerous population of cells [50]. Additionally, the shortest telomere amongst a cell possesses a dominant negative action, thus a cell is ‘’just that good in the manner of its shortest telomere [51].
Determination of telomere length in single cells is feasible by the utilization of qFISH or qRT-PCR. In case of quantitative fluorescence in situ hybridization (Figure 1) in combination with fluorescent peptide nucleic acids whose binding takes place with the telomeric repeats stoichiometrically (in appropriate proportion of reagents). The germane T length gets conducted by this quantitative fluorescence in situ hybridization [22], which is appropriate just for metaphase arrested cells for instance oocyte, or polar bodies, in cases possessing Ts which are snuggishly compacted. With regards to embryos qFISH needs pharmacologic metaphase arrest that is not practically feasible in case of human embryos [52].
qRT-PCR possesses the capacity of determination of T’s in abundance of quantities . Quantitative PCR possesses the capacity of determination of Ts in ascendable quantities in addition to is practical for the maximum clinical samples, however possesses a myriad of restrictions. With the quantitative PCR determination of germane T’s quantities takes place, however not the complete quantities [53]. Apart from chromosome ends (for instance, amongs T’s) telomeric repeats are further observed scattered right through the genome particularly in the subtelomeric areas. Thereby qRT- PCR determines T DNA quantities instead of length. Lastly qRT-PCR determines average T quantities, thereby significant knowledge gets eliminated with regards to short T’s.
Regarding qRT-PCR in case of bulk cells, DNA identification, its quantification in addition to dilution is performed [53]. Normalization of telomere amplification is conducted to a single copy (for instance albumin) gene (S) leading to a T:S ratio percentage to the mean T quantities in the population. The T:S ratio gets calculated by utilization of the Ϫ-Ϫcycle threshold (Ct) determination: ϪϪ Ct=Ϫ Ct(sample)-Ϫ Ct (control mean). Expression of germane T length is done in the form of fold expression=2^( ϪϪ Ct). Cycle threshold results for telomere as well as albumin are generated by the utilization of real-time PCR.
• Single Cell Amplification of Telomeric Repeats PCR Wang F, et al. [50], performed adaptation of the qPCR for the assays of the separate cells for instance oocyte, sperm, along with the polar bodies named single cell amplification of telomeric repeats PCR [50]. Single cell get obtained in addition to assayed in the PCR tubes. This is followed by the thawing as well as lysing of the cells, subsequently pre amplification of DNA with the utilization of probes for T along with the multi copy reference(R) gene (ALU). Quantitative PCR generates a T: R ratio determined by ϪϪ Ct.
• Terminal Restriction Fragment Analysis Terminal restriction fragment analysis determines entire T length, however, just in case of a larger population of cells for instance leukocytes or endometrial cells. Other assays estimate germane T DNA quantities [54]. Terminal restriction fragment analysis is restricted to a larger population of cells in addition to possesses restricted ascendability. Digestion of μg’s of genomic DNA is conducted with the particular restriction enzymes by which introduction of plethora of cuts is made commonly right through the genome but for the T’s, whose sparing occurs. Gel electrophoresis segregates larger fragments of the T DNA which is left uncut, whose hybridization is performed with the utilization of T particular probes with the subsequent southern blotting. Imaging for blots as well as T length is estimated with the utilization of contrasting to reference DNA.
• Ubiquitous Single Telomere Length Analysis The fate of the cells is based on the shortest instead of average T length, thereby the strategies estimating shortest T length is of significance. Ubiquitous Single telomere length analysis evaluates the load of short T’s in the bulk T cells [55]. Digestion of T cells takes place by the restriction endonucleases. Subsequent to the digestion by enzymes there is persistence of the <3kb genomic DNA. Nevertheless, there is persistence of the intactness of the T DNA along with sticky ends get generated in the sub telomeric areas. Ligation of the sticky ends takes place with oligos in addition to a telorette oligo acclaims the 5’ overhang at the T end. Fill in’s as well as the adapter proteins aid in amplification of short T’s selectively. Over agarose gel samples are run with the subsequent shifting to the nylon membrane for southern blotting. Telomere bands <5kb are referred to as the short T’s, along with <2kbs are referred to as crucially short T’s.
Telomeres along with the Germline
The sustenance of the germline stem cells population in case of the testis, spermatogonia, where persistently telomerase continues to be active for the promotion of the persistent generation of germ cells right through the life of the man. Sperms portray just the cell kinds whose T’s keep lengthening with the age plausibly in view of the actions of the constitutive telomerase action [56]. The heterogeneity of the T length in case of sperms belonging to the same population further escalates with the age that further makes ALT dependent T lengthening responsible at the time of spermatogenesis [57].
The female germline possesses least telomerase actions [58], subsequent to the stoppage of oogonial proliferation at about 10-20 wks of fetal generation at the time when oocytes go through meiotic arrest [59]. Subsequent to the initiation of meiotic arrest, shortening of T’s in case of oocytes can take place through Reactive oxygen species (ROS), or other environmental toxins (the way detailed previously). One layer of somatic cells surround the oocyte to generate a primordial follicle (PF). Maximum PF‘s finally become atretic, however a subset to activate to start with independent along with based on gonadotropin liberation. Follicles having resumption of growth have acquisition of cumulus as well as granulosa cells (GCs) to support them, that illustrate expression of telomerase actions [58, 60].
Role of Telomeres, Telomerase along with ALT at the time of Early Embryo Generation
The fusion of the oocyte with the sperm form a diploid zygote that is based on protein in addition to messenger ribonucleic acid (mRNA) which is stored in the oocyte till the initiation of the zygote transcription. Liu L, et al. [52], illustrated earlier in mice T lengthening at the time of first embryonic cell cycle, modulated by ALT [52], in case of mice ALT actions occur at the 2 cells stage concurrently with the zygote genome activation. By the time of blastocyst stage, embryonic cells have segregated into an inner cell mass (ICM) in addition to an outer cell mass. Telomeres are existent in the ICM of the mouse blastocyst (possess the capacity of becoming embryonic stem cells (ESCs) on in vitro culturing) possess T’s whose reconstitution has been previously obtained by the utilization of ALT prior to the time of reactivation of T’s [52, 61]. Alternative lengthening of telomeres modulated T lengthening occurs in parthenotes as well along with in case of telomerase-null mice, pointing to the oocytes independently possessing the capacity of activation of this mode inheritantly [52]. The ICM cells possess telomerase actions that aids in sustenance of the T length till the time of differentiation initiation, when shortening of T’s gets started (Figure 6) [62].
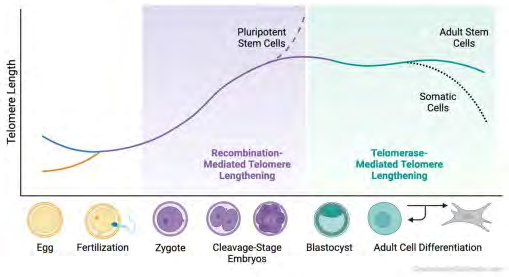
Figure 6: Courtesy ref no-62-Telomere dynamics throughout the mouse lifespan. Telomeres in oocytes are exceptionally short, and in sperm, they are exceptionally long. At the 2-cell stage, the zygote’s critically short telomeres lengthen via a recombination-based alternative lengthening of telomeres. Telomerase becomes active only in the late morula and blastocyst stages. The kinetics of telomerase are such that it is likely involved in telomere maintenance. Adult stem cells retain robust telomerase activity, unlike somatic cells, which undergo telomere attrition and consequent cellular senescence.
In case of mice studies illustrated potent telomerase actions, however initiated just at the blastocyst stage [61]. Mice possessing null mutations in the telomerase gene, possess short T’s along with illustrate the ageing phenotypes, inclusive of reproductive ageing with infertility, meiotic impairment , potent blastocyst ageing, abnormalities in gametogenesis in addition to gonadal agenesis [29, 63, 64]. Telomerase reactivation possess the capacity of reverting certain of these phenotypes in addition to leads to lengthening of the lifespan, corroborating the correlation amongst T length in addition to ageing [65, 66]. Telomerase works at the time of S phase of the cell cycle, that in case of female germ cells occurs just at the time of fetal oogonial replication. Thereby, it has been posited that attempting telomerase activation in case of ageing females would not be efficacious in reverting oocytes ageing.
Significant variations are existent in the T dynamics amongst mice along with humans. T length in case of mice is 10 fold (100kb) those of the humans (5-15 kb). Interestingly, female mice do not undergo age correlated oocyte impairment to the equivalent extent-like women-hypothalamus as well as uterus portray the primary loci of the reproductive ageing in mice. In view of telomeres in case of telomerase-null mice undergo propagative weakening, nevertheless, they form oocytes impairment in view of T length nears that of humans. telomerase-null mice have the capacity of survival in the various generations, however illustrate propagative T enfeebling with the every generation [51]. By the time of generation3, since T length’s approximate in women ,they illustrate T impairment , with corroboration of ageing in plethora of organs inclusive of gonads.
Ultra short T’s in case of human oocytes in addition to early preimplantation embryo might result in susceptibility to the T end fusions in addition to aneuploidy [67]. T length further escalate at the time of early human generation [68], despite zygote genomic activation occurs amongst the4&8 cell stages of generation [69]. Noticeably, T length variation is not amongst euploid as well as aneuploid blastocysts, pointing that embryonic generation is not an adequate criterion for differentiating ploidy in embryos. Acknowledged the considerable variations in case of T dynamics amongst species at the time of early human preimplantation embryonic generation extra studies are required.
Retrotransposons at the time of Early Generation
A plethora of species for instance Drosophila utilize variable approaches for the capping as well as protecting chromosome ends: retrotransposons. Retrotransposons portray repeated DNA sequences, having the ability of motion right through the genome with the utilization of reverse transcriptase in addition to endonuclease with the idea of ’’cut and paste’’ themselves at newer regions. Retrotransposons possess a significant part in genomic structure at the time of early generation [70, 71]. Long interspersed nuclear element 1 (LINE1) represents the lone autonomous retrotransposon in the human genome, having 50-100 copies continuing to be active. Retrotransposons possess a remarkable resemblance with the T’s inclusive of considerable repeated sequences, ii)them being relying on reverse transcriptase in addition to transcription of noncoding RNAs (telomeric repeats possessing RNA). In agreement with a plausible part for retrotransposons at the time of ALT in cleavage- stage embryos, Wang F, et al. [71], observation was ahamperor of retrotranspositon resulted in blockade of the ALT as well as T lengthening in cleavage- stage embryos [71]. Conversely, critically short T’s possessed the capacity of desuppressing the retrotransposons inclusive of LINE1 resulting in genomic instability [70].
Telomeres Length Associates with Pluripotency
Telomere reprogramming takes place just at the time of early generation inclusive of induced pluripotent stem cells (iPSCs) reprogramming, in addition to somatic cell nuclear transfer (SCNT). These processes form pluripotent stem cells, suggesting an association amongst reconstitution along with pluripotency [72, 73]. Embryonic stem cells (ESC) possess the capability of apart from differentiation in practically each embryonic tissue, however in addition lead to unrestricted replication, promoted by their capability of sustenance of T reserves. SCNT, reprograms differentiated cells to pluripotency with greater completeness in addition to their reprogramming is more advantageous in contrast to iPSCs. ESC‘s obtained from SCNT possess more differential probability as well as improvement of chimera generation in contrast to iPSCs from the same mice [65]. SCNT further displayed better proliferation as well as T elongation in contrast to iPSCs. Improvement of pluripotency being induced following SCNT experiment is assumed to get obtained from better rejuvenation probability of the oocytes. Furthermore, telomerase re-activation apparently takes place earlier in contrast to pluripotent genes (Oct4 as well as Nanog) at the time of reprogramming, pointing that telomere regulation is both a key step in development of pluripotency along with imperative for sustenance of stem cell-dependent tissue homeostasis [74].
Telomeres along with Reproduction
Telomeres have been associates with cell ageing, arrest as well as demise in a myriad of cell kinds [4], there by the group of Keefe DL, et al. [75] proposed that they might be further controlling the working of the oocyte. Oocytes belonging to the telomerase-null mice form diminished synapsis as well as chiasmata [64], along with finally generate escalated T signal ends, chromosome end to end fusion, T uncapping, genomic instability, meiotic spindle aberrations along with subfertility [29, 63, 75]. G6 telomerase-null mice possess escalated apoptosis in testiss, diminished ovarian weight in addition to oocytes obtained along with reduction in the morula generation without any reaching the blastocyst stage [29, 63, 75].
In case of humans, spontaneous genetic mutations of one of the telomere constituents result in telomere sapping, a situation referred to as telomeropathies [35, 36, 37, 41, 42, 43, 44], or dyskeratosis congenital (DC). manifestation of DC is in the form of epidermal aberrations, augmentation of the shortening of T’s, age correlated pathologies associated with diminished lifespan. DC patients portray a natures’s experiment for asssessment of the Tposit of reproductive ageing in women. In agreement with the proposal of Wang F, et al. [74] group women with DC illustrate a reproductive phenotype substantially akin to telomerase-null mice [38]. Diminished ovarian reserve (DOR) has been revealed in women having DC [37]. Furthermore, they observed embryonic generation was not good/efficacious in addition to greater rates of aneuploidy in case of a 30 yr old DC patient going through invitro fertilization (IVF) as well as preimplantation genetic testin for aneuploidy (PGT-A), with the idea of fertility preservation. She displayed substantially DOR, T weakening in the oocytes, embryos along with leukocytes in addition to aberrant embryonic generation [53]. The quantities of antimullerian hormone (AMH) in her as 0.3ng/ml, that was 10times lesser in contrast to age matched controls. Controlled ovarian stimulation needed double the dose along with time period, attaining 50% of follicles as well as oocyte quantities vis a vis age along with body mass index (BMI) matched controls. Ubiquitous Single telomere length analysis illustrated diminished cortically short(≤2kb) subsequent to ovarian stimulation. There is no Clarification if estrogen response element (ERE) in the TERT promoter modulates these actions, however Interestingly, ovarian stimulation further results in T lengthening in mice [76].
Moving to the Posit of Reproductive Ageing
The oocyte portrays the locus of reproductive ageing in case of women, the manner validated by the capacity of the donation of the oocytes from the younger to the older women with the view of ameliorating the actions of ageing [77]. The modes behind the actions of ageing over oocyte continues to be debatable. Obviously ageing impact plethora of actions on oocyte working, however reciprocal nuclear transfer amongst young as well as old oocytes illustrates that the ageing phenotype separates with the nucleus instead of the cytoplasm [78]. BobEdwards, the pioneer of IVF, illustrated a ‘’generation line’’ at the time of mouse oogenesis, where the initial oocytes to leave the oogonial proliferation at the time of fetal life are the ones which undergo ovulation first at the time of puberty in addition to the last oocytes to leave the oogonial proliferation are the last to undergo ovulation in the adult [79]. Replication - correlated T sapping gives a molecular reinforcing to BobEdwards’generation line’’. Hence Keefe DL, et al. [75] group performed modifications to this posit which is inclusive of a ‘’second hit’’ to the T’s on the oocytes continued lifespan, the accrual actions of the ROS [80], cigarette smoking as well as environmental toxins also abrogating oocyte T’s [24, 62]. The marginal intactness of the spindle assembly check point aids in [81] in the propagation of the structurally dysfunctional oocytes along with the early embryos.
On ageing of the women, there is reduction of both the follicle reserve in addition to ovarian generation probability, which is not synchronous all the time. The follicle reserve might be determined by the antral follicle count along with the antimullerian hormone (AMH) quantities, however there is dearth of detectors of the oocyte generation plausibility. Certain proof points that T’s in polar bodies anticipate embryonic generation, fragmentation, aneuploidy, clinometric (a domain correlated with indexes, rating scales & other expression used to detail /estimate symptoms physical signs & other clinical phenomena) studies are required for the estimation of the plausible clinical significance of the polar body T length for the anticipation of embryo viability [29, 82].
Conclusion
With the tendency of the postponement of reproduction in addition to escalation of utilization of assisted reproductive technologies have isolated the significant part of the oocyte in case of reproductive ageing [78, 83]. Concurrently with the utilization of biogerontology the modes behind the manner cellular ageing has been conserved in nature has been displayed. Telomere debilitation correlates plethora of variable characteristics of the ageing in the oocyte portrays cells having a longer life, which stimulates the group of Kalmbach K, et al. [75] to hypothesize a telomere posit of reproductive ageing [29, 63, 84]. Rodent experiments corroborate a main significant part of the T’s in case of reproduction. Mouse oocytes illustrating least ageing in vivo possess significantly longer T’s in contrast to human’s [79, 85], however genetic or pharmacologic T shortening recounts the phenotype of the oocyte ageing in case of humans. Restricted validation from the experiments of nature for instance telomeropathies is in agreement with the posit of reproductive ageing. Further studies need to investigate if estimation of T possesses the capacity of evaluating generational oocyte plausibility. Moreover as detailed earlier psychological stress itself causes T shortening of T’s which get lengthened subs to rajyoga meditation which might aid in unexplained infertilityas corroboratedby EEG/MRI studies [86]. Though sperms form all through life of male by ALT, have telomerase active progenitor, spermatogonia, longest body T’, recently Dhillon VS, et al. [87], displayed reduced SIRT1 and SIRT3 and lower antioxidant capacity of seminal plasma is shorter sperm telomere length in oligozoospermic men [87]. Their conclusions were shorter TL in sperms as well as leukocytes is correlated with escalated oxidative stress (OS) which is further implicated in greater quantities of DNA fragmentation in sperms. Their outcomes corroborate obtained the posit that variable sperm paradigm at the time of oligospermia are correlated with or a result of diminished quantities of SIRT1 and SIRT3 proteins [87].
References
-
Meyne J, Ratliff RL, Moyzis RK (1989) Conservation of the human telomere sequence (TTAGGG)n among vertebrates. Proc Natl Acad Sci USA 86(18): 7049-7053.
-
Griffith JD, Comeau L, Rosenfield S, Stansel RM, Bianchi A, et al. (1999) Mammalian telomeres end in an large duplex loop. Cell 97(4): 503-514.
-
Lange TD (2005) Shelterin: the protein complex that shapes and safeguards human telomeres. Genes Dev 19(18): 2100-2110.
-
Letsolo BT, Rowson J, Baird DM (2010) Fusions of short telomeres in human cells is characterized by extensive deletion and microhomology, and can result in complex rearrangements. Nucleic Acids Res 38: 1841-1852.
-
Shiloh Y (2003) ATM and related protein kinases: safeguarding genome integrity. Nat Rev Cancer 3(3): 155-168.
-
Zou L, Elledge SJ (2003) Sensing DNA damage through ATRIP recognition of RPA-ssDNA complexes. Science 300(5625): 1542-1548.
-
Morrish TA, Greider CW (2009) Short telomeres initiate telomere recombination in primary andtumor cells. PLoS Genet 5(1): e1000357.
-
Calado RT, Young NS (2009) Telomere Diseases. N Engl J Med 361(24): 2353-2365.
-
Palm W, Lange TD (2008) How shelterin protects mammalian telomeres. Annu Rev Genet 42: 301-334.
-
Broccoli D, Smogorzewska A, Chong L, Lange TD (1997) Human telomere contain two distinct Myb- related proteins, TRF1 and TRF2. Nat Genet 17(2): 231-235.
-
Liu D, Safari A, Connor MS, Chan DW, Laegeler A, et al. (2004) PTOP interacts with POT 1 andregulates its localization to telomeres. Nat Cell Biol 6: 673-680.
-
Li B, Oestreich S, Lange TD (2000) Identification of human rap1: implications for telomere evolution. Cell 101(5): 471-483.
-
Denchi EL, Lange TD (2007) Protection of telomeres through independent control of ATM and ATR by TRF2 and POT 1. Nature 448(7157): 1068-1071.
-
Guo X, Deng Y, Lin Y, Cosme-Blanco W, Chan S, et al. (2007) Dysfunctional telomeres activate an ATM ATR dependent DNA damage respone to suppressive tumorigenesis. EMBO J 26(22): 4709-4719.
-
Celli GB, Lange TD (2005) DNA processing is not required for ATM mediated telomere damage respone after TRF2 depletion. Nat Cell Biol 7(7): 712-718.
-
Benarroch-Popivker D, Pisano S, Mendez-Bermudez A, Lototska L, Kaur P, et al. (2016) TRF2 mediated control of telomere DNA topology as a mechanism of chromosome end protection. Mol Cell 61(2): 274-286.
-
Lu W, Zhang Y, Liu D, Songyang Z, Wan M (2013) Telomeres, Structure functions and regulation. Exp Cell Res 319(2): 133-141.
-
Kaur KK (2023) Editorial: Reviews in assisted reproduction: 2022. Front Reprod Health 5: 1174047.
-
Bryan TM (2020) G-quadriplexes at telomeres: friend or foe? Molecules 25(16): 3686.
-
Blackburn EH (2005) Telomeres and telomerase: their mechanisms of actions and the effects ofaltering their functions. FEBS Lett 579(4): 859-862.
-
Sfeir AJ, Chai W, Shay JW, Wright WE (2005) Telomere end processing the terminal nucleotides of Human chromosomes. Mol Cell 18(1): 131-138.
-
Martens UM, Chavez EA, Poon SS, Schmoor C, Lansdorp PM (2000) Accumulation of short telomeres in human fibroblasts prior to replicative Senescence. Exp Cell Res 256(1): 291-299.
-
Zglinicki TV (2002) Oxidative stress shortens telomeres. Trends Biochem Sci 27(7): 339-344.
-
Epel ES, Blackburn EH, Lin J, Dhabhar FS, Adler NE, et al. (2004) Accelerated telomere shortening in response to life stress. Proc Natl Acad Sci USA 101: 17312-17315.
-
Huang J, Okuka M, McLean M, Keefe DL, Liu L (2010) Telomere susceptibility to cigarette smoke-induced Oxidative damage and chromosomal instability of mouse embryos in vitro. Free Radical Biol Med 48(12): 1663- 1676.
-
Rewak M, Buka S, Prescott J, Vivo ID, Loucks EB, et al. (2014) Race related health and biological ageing: does rate of telomere shortening differ across blacks and white? Biol Psychol 99: 92-99.
-
Denham J, Nelson CP, O’Brien BJ, Nankervis SA, Denniff M, et al. (2013) Longer leukocyte telomeres are associated with ultraendurance exercise independent of cardiovascular risk factors. PLoS ONE 8(7): e69377.
-
Kim NW, Piatyszek MA, Prowse KR, Harley CB, West MD, et al. (1994) Specific association of human telomerase activity with immortal cells and cancer. Science 266(5193): 2011-2015.
-
Keefe DL, Liu L, Marquard K (2007) Telomeres and ageing related meiotic dysfunction in women. Cell Mol Life Sci 64: 139-143.
-
Lopez-OtinC, Blasco MA, Partridge L, Serrano M, Kroemer G (2013) The hallmarks of aging. Cell 153(6): 1194-1217.
-
Greider CW, Blackburn EH (1985) Identification of a specific telomere terminal transferase activity in tetrahymena extracts. Cell 43: 405-413.
-
Cesare AJ, Reddel RR (2010) Alternative lengthening of telomeres models, mechanisms and implications. Nat Rev Genet 11(5): 319-330.
-
Britt-Compton B, Capper R, Rowson J, Baird DM (2009) Short telomeres are preferentially elongated by telomerase in human cells. FEBS Lett 583(18): 3076- 3080.
-
Weinrich SL, Pruzan R, Ma L, Ouellette M, Tesmer VM, et al. (1997) Reconstitution of human telomerases with the RNA template component hTR and the protein catalytic subunit hTRT. Nat Genet 17(4): 498-502.
-
Batista LFZ, Pech MF, Zhong F, Nguyen HN, Xie KT, et al. (2010) Telomere shortening and loss of self renewal in dyskeratosis congenita induced pluripotent stem cells. Nature 464: 292-296.
-
Shay JW, Wright WE (2004) Telomeres in dyskeratosis congenital. Nat Genet 36(5): 437-438.
-
Ballew BJ, Savage SA (2011) Updates on the biologyof dyskeratosis congenita and related telomere biology disorders. Expert Rev Haematol 474: 399-402.
-
Robinson LG, Pimentel R, Wang F, Kramer YG, Gonullu DC, et al. (2020) Impaired reproductive function and fertility preservation in a woman with a dyskeratosis congenita. J Assist Reprod Genet 37(5): 1221-1225.
-
Liu J, Wang L, Wang Z, Liu JP (2019) Roles of telomere biology in cell senescence, replicative and chronological ageing. Cells 8(1): 54.
-
Vulliamy TJ, Walne A, Baskaradas A, Mason PJ, Marrone A, et al. (2005) Mutations in the reverse transcriptase component of telomerase (TERT) in patients with Bone marrow failure. Blood Cells Mol Dis 34(3): 257-263.
-
Zhong F, Savage SA, Shkreli M, Giri N, Jessop L, et al. (2011) Disruption of telomerase trafficking byTCAB1 mutations causes dyskeratosis congenita. Genes Dev 25(1): 11-16.
-
Savage SA, Giri N, Baerlocher GM, Orr N, Lansdorp PM, et al. (2008) TINF2, a component of the shelterin telomeres protection complex, is mutated in dyskeratosis congenita. Am J Hum Genet 82(2): 501-519.
-
Keller RB, Gagne KE, Usmani GN, Asdourian GK, Williams DA, et al. (2012) CTC1 mutations in a patient with dyskeratosis congenita. Paediatr Blood Cancer 59(2): 311-314.
-
Ballew BJ, Yeager M, Jacobs K, Giri N, Boland J, et al. (2013) Germline mutations of regulator of telomere elongation helicase 1 RTEL1 in dyskeratosis congenita. Hum Genet 132(4): 473-480.
-
Natarajan S, Groff-Vindman C, McEachern MJ (2003) Factors influencing the recombination expansion and spread of telomeric tandem assays in Kluyveromyces lactis. Eukaryot Cell 2(5): 1115-1127.
-
Nosek J, Rycovska A, Makhov AM, Griffith JD, Tomaska L (2005) Amplificationof telomeric arrays via rolling circle mechanism. J Biol Chem 280(11): 10840-10845.
-
Topcu Z, Nickles K, Davis C, McEachern MJ (2005) Abrupt disruption of capping and a Single source for recombinationationally elongated telomeres in Kluyveromyces lactis. Proc Natl Acad Sci USA 102(9): 3348-3353.
-
Muntoni A, Neumann AA, Hills M, Reddel RR (2009) Telomere elongation involves intra-molecular DNA replication in cells utilizing alternative lengthening of telomeres. Hum Mol Genet 18(6): 1017-1027.
-
Yeager TR, Neumann AA, Englezou A, Huschtscha LI, Noble JR, et al. (1999) Telomerase negative immortalized human cells contain anovel type of promyelocytic leukaemia (PML) body. Cancer Res 59(17): 4175-4179.
-
Wang F, Pan X, Kalmbach K, Seth-Smith ML, Ye X, et al. (2013) Robust measurement of telomere length in Single cells. Proc Natl Acad Sci USA 110(21): E1906-E1912.
-
Hemann MT, Strong MA, Hao LY, Greider CW (2001) The shortest telomere,not average telomere length is critical for cell viability and chromosomal stability. Cell 107(1): 67-77.
-
Liu L, Bailey SM, Okuka M, Muñoz P, Li C, et al. (2007) Telomere lengthening early in development. Nat Cell Biol 9(12): 1436-1441.
-
Cawthon RM (2002) Telomere measurement by quantitative PCR. Nucleic Acids Res 30(10): e47.
-
Kimura M, Stone RC, Hunt SC, Skurnick J, Lu X, et al. (2010) Measurement of telomere length by the southern blot analysis of terminal restriction fragment lengths. Nat Protoc 5: 1596-1607.
-
Bendix L, Horn PB, Jensen UB, Rubelj I, Kolvraa S (2010) The load of short telomeres estimated by a new method, Universal STELA, correlates with number of senescent cells. Ageing Cell 9(3): 383-397.
-
Stindl R (2016) The paradox of longer sperm telomeres in older mens testis:a birth cohort effect caused by transgenerational telomere erosion in the female germline. Mol CytoGenet 9: 12.
-
Antunes DM, Kalmbach KH, Wang F, Dracxler RC, Seth- Smith ML, et al. (2015) A single cell of for telomere DNA content shows increasing telomere length heterogeneity as well as increasing mean telomere length in human spermatozoa with advancing age. J Assist Reprod Genet 32: 1685-1690.
-
Wright DL, Jones EL, Mayer JF, Oehninger S, Gibbons WE, et al. (2001) Characterization of telomerase activity in the human oocyte and preimplantation embryo . Mol Hum Reprod 7: 947-955.
-
Holt JE, Lane SI, Jones KT (2013) The control of meiotic maturation in mammalian oocytes. Current topics in developmental biology 102: 207-226.
-
Ozturk S, Sozen B, Demir N (2014) Telomere length and telomerase activity during oocyte maturation and early embryo development in mammalian species. MolHum Reprod 20(1): 15-30.
-
Varela E, Schneider RP, Ortega S, Blasco MA (2011) Different telomere length dynamics in the innercell mass versus established embryonic stem(ES) cells . Proc NatlAcad Sci USA 108(37): 15207-15212.
-
Robinson LGJ, Kalmbach K, Sumerfield O, Nomani W, Wang F, et al. (2024) Telomere dynamics and reproduction. Fertil Steril 121(1): 4-11.
-
Keefe DL (2020) Telomeres and genomic instability during early development. Eur J Med Genet 63(2): 103638.
-
Liu L, Franco S, Spyropoulos B, Moens PB, Blasco MA, et al. (2004) Irregular telomeres impair meiotic synapsis and recombination in mice. Proc Natl Acad Sci USA 101(17): 6496-6501.
-
Le R, Kou Z, Jiang Y, Li M, Huang B, et al. (2014) Enhanced telomere rejuvenation in pluripotent stem cells reprogrammed via nuclear transfer relative to induced pluripotent stem cells. Cell Stem Cells 14(1): 27-39.
-
Townsley DM, Dumitriu B, Liu D, Biancotto A, Weinstein B, et al. (2016) Danazol treatment for telomere diseases. N Engl J Med 374(20): 1922-1931.
-
Kohlrausch FB, Wang F, Chamani I, Keefe DL (2021) Telomere shortening and fusion: A link to early human embryo development. Obs Gyn Survey 76(7): 429-436.
-
Wang F, McCulloh DH, Chan K, Wiltshire A, McCaffrey C, et al. (2023) Telomere length and telomerase in human embryos at blastocyst stage. Genes (Basel) 14(6): 1200.
-
Egli D, Chen AE, Saphier G, Ichida J, Fitzgerald C, et al. (2011) Reprogramming within hours following nuclear transfer into mouse but not human zygotes. Nat Commun 2: 488.
-
Zhao N, Yin G, Liu C, Zhang W, Shen Y, et al. (2023) Critically short telomeres derepress retrotransposons to promote genomic instability in embryonic stem cells. Cell Discov 9(1): 45.
-
Wang F, Chamani IJ, Luo D, Chan K, Navarro PA, et al. (2021) Inhibition of LINE1 retrotranspositon represents telomere reprogramming during mouse 2-cellembryo development. J Assist Reprod Genet 38(12): 3145-3153.
-
Tachibana M, Amato P, Sparman M, Gutierrez NM, Tippner-Hedges R, et al. (2013) Human embryonic stem cells derived by somatic cell nuclear transfer. Cell 153: 1228-1238.
-
Yamada M, Johannesson B, Sagi I, Burnett LC, Kort DH, et al. (2012) Human oocytes reprogramme adult somatic nuclei of a type1 diabetic to diploid pluripotent stem cells. Nature 510(7506): 533-536.
-
Wang F, Yin Y, Ye X, Liu K, Zhu H, et al. (2012) Molecular insights into the heterogeneity of telomere reprogramming in induced pluripotent stem cells. Cell Res 22(4): 757-768.
-
Kalmbach K, LG Robinson, Wang F, Liu L, Keefe D (2014) Telomere length reprogramming in embryos and stem cells. Biomed Res Int 2014: 925121.
-
Berteli TS, Wang F, Kohlrausch FB, Luz CMD, Oliveira FV, et al. (2022) Impact of super ovulation and in vitro fertilization on LINE-1 copy number and Telomere length in C57/BL/6 J mice. Mol Biol Rep 49: 4909-4917.
-
Tarín JJ, García-Pérez MA, Cano A (2014) Assisted reproductive technology results: why are live birth percentages so low? Mol Reprod Dev 81(7): 568-583.
-
Liu L, Keefe DL (2004) Nuclear origin of aging associated meiotic defects in senescent- accelerated mice. Biol Reprod 71: 1724-1729.
-
Edwards RG (1970) Are oocytes formed and used sequentially in the mammalian ovary? Philos Trans R Soc Lond B Biol Sci 259(828): 103-115.
-
Collins JK, Jones KT (2016) DNA change responses in mammalian oocytes. Reproduction 152: 815-822.
-
Gui L, Homer H (2012) Spindle assembly checkpoint signaling is uncoupled from chromosomal position in mouse oocytes. Develop 139(11): 1941-1946.
-
Keefe DL, Franco S, Liu L, Trimarchi J, Cao B, et al. (2005) Telomere length predicts embryo transplantation after in vitro fertilization in women-- toward telomere a theory of reproductive ageing in women. Am J Obstet Gynaecol 192: 1256-1260.
-
Homan GF, Davies M, Norman R (2007) The impact of lifestyle factors on reproductive performance in thegeneral population and those undergoing infertility treatment: a review. Hum Reprod Update 13(3): 209- 223.
-
Kalmbach KH, Antunes DMF, Dracxler RC, Knier TW, Seth-Smith ML, et al. (2013) Telomeres and human reproduction. Fertil Steril 99(1): 23-29.
-
Yamada-Fukunaga T, Yamada M, Hamatani T, Chikazawa N, Ogawa S, et al. (2013) Age associated telomere shortening in mouse oocytes. Reprod Biol Endocrinol 11: 108.
-
Kaur KK, Allahbadia GN, Singh M (2023) The Science Behind Rajyoga Meditation-A Narrative Review. Adv Yoga Physical Ther 1(2): 26-43.
-
Dhillon VS, Shahid M, Deo P, Fenech M (2024) Reduced SIRT1 and SIRT3 and lower antioxidant capacity of seminal plasma is shorter sperm telomere length in oligozoospermic men. Int J Mol Sci 25(2): 718.
- Postpartum Maternal Mental Health - A Narrative Review
- Beta HCG in Cervico-Vaginal Secretion as a Predictor of Preterm Delivery
- Successful Management of Mid Trimester Foetal Death with Major Placenta Previa by Expectant Management Followed by Induction of Labour
- To Evaluate the Expression of Egr2 Gene in Term Low Birth Weight Newborns
- Impact of Maternal Obesity on Maternal and Foetal Outcomes: A Prospective Cohort Study from Northern India
- ‘’Benefit of Pulsatile GnRH Therapy in Treatment of Functional Hypothalamic Amenorrhea (FHA) and Congenital Hypogonadotropic Hypogonadism(CHH) in Infertile Patients Over Canonical Gonadotropins with IVF –A Short Communication’’