Effects of Different Oxygen Levels and Time Points on Mitochondrial Respiratory Functional Analysis of Placental Tissue
The placenta plays an important role in fetal growth and development. The Oxygraph-2k mitochondrial function respirometer provides an accurate and realistic quantitative analysis of mitochondrial respiratory function and the energy metabolism of organisms. However, this machine is used less frequently in placental tissues, which consume do not consume as much oxygen as the heart, brain, or skeletal muscle tissues, therefore, it is crucial to develop a standard respiratory measurement protocol for placental tissues. For this, we tested varying oxygen concentrations (150–160 (ambient air), 200, 250, and 300 μM) and time tissue remains viable in respiratory medium (1.5~3, 3~4.5, 4.5~6, and 6~7.5 hour) to determine the optimal experimental conditions. As a result, we found no significant changes in mitochondrial respiratory capacity at different oxygen levels, whilst there was a decrease in mitochondrial respiratory capacity more than 3 hour after delivery.
Introduction
The placenta is not only a bridge for material exchange between the mother and fetus, but also has important immune, excretory, defensive, and endocrine functions [1]. Mitochondria are vital to virtually every aspect of cellular function, such as the supply of energy in the form of ATP(Adenosine triphosphate), processing metabolic intermediates, regulating Ca2+ signalling, controlling ROS (reactive oxygen species) production and playing a critical role [2]. The human placenta engages in high metabolic plasticity to meet dynamic oxygen tension changes during pregnancy [3, 4, 5].The placenta has been central to the pathophysiological study of hypoxia-related pregnancy disorders such as preeclampsia [6], where pathophysiological alterations can lead to a state of oxidative stress and a reduction in the capacity of mitochondria in the placenta [7]. Many previous studies have shown that mitochondrial dysfunction contributed to placental pathology underpinning gestational disorders [8].
Therefore, the study of placental mitochondria for both normal pregnancy and in the case of pregnancy complications is crucial. Mitochondrial function in a variety of tissues, including the placenta and isolated trophoblasts, has been assessed using mitochondrial respirometers (Oxygraph Respirometer, O2K) [9, 10, 11, 12]. However, there has been no previous attempt to standardize protocols for any of these tissues or cells with regard to the oxygen concentrations used to assess mitochondrial respiratory function. Furthermore, no attempt has yet been made to determine how long placental mitochondria remain viable in respiratory medium for respirometry studies.
Therefore, this study was designed to determine whether variations in oxygen concentration in the O2K chamber influenced the function of placental mitochondria, in addition to how long mitochondria remain viable in the respiratory medium.
Methods
Sample Collection and Materials
This research recruitment period runs from December 25, 2020 to December 25, 2022. Placental tissues were collected from 50 normal term (37–40 weeks of gestation) pregnancies, delivered through elective Caesarean section (multiple Caesarean section history and cephalopelvic disproportion). Samples from the foetal side of the placenta (50 mg) were biopsied within 15 min (minute) of delivery, mixed with PBS (Phosphate Buffered Saline), and delivered on ice to the laboratory. Placental villous tissue samples were then prepared by referring to the processing as previously described in a video [13]. Simply put, placental tissue is placed in a 10 cm diameter petri dish containing PBS, thoroughly rinsed, and the blood is washed out to the greatest extent possible, then the left hand holds the tweezers, and the right hand holds the glass slide, scraping the villi tissue, and removing the fibrous tissue and blood vessels to the greatest extent possible.
Human Ethical Approval Statement
Subjects were recruited from the Qinghai University Affiliated Hospital with the approval of the Qinghai University Affiliated Hospital Ethics Committee, and after obtaining informed written consent.
High-Resolution Respirometry
Samples were then washed in ice-cold PBS and permeabilized in saponin (50 ug) in BIOPS (1 mL) solution for 40 min on ice with continual mixing. Samples were subsequently washed twice for 10 min on ice in MiR05 respiration medium, as has been previously described [14]. The speed of the shaker was approximately 100–110 r/min, although this speed could not be too fast in order to prevent the mitochondrial membrane from breaking. Next, blot-dried before measuring wet weight, 5–10 mg wet weight was added to the O2K chamber containing respiratory medium MiR05 at 37℃ and was then exposed to varying supplemental oxygen levels (150–160, 200, 250, and 300 μM). The machine used was located at an altitude of 2200 m above sea level in Xining, Qinghai Province, with an atmospheric pressure of 581.57 mmHg and an oxygen pressure of 16.24 kPa. The machine was also subjected to final and proper calibration using zeroing powder every three months. In addition, air calibration was performed before each experiment, with the oxygen concentration in the chamber after this being approximately 150–160 uM. The first part of this experiment was designed to verify the effect of different oxygen concentrations in the chamber on mitochondrial respiratory function, whilst the second part aimed to study the duration of osmotic mitochondrial activity stored in the mitochondrial respiratory fluid. Sample preparation is estimated to take 1.5 hours from delivery to penetration to detection of oxidative phosphorylation. It then took approximately 1.5 h further to measure a single parameter of mitochondrial respiratory function. Permeabilized placental villous tissue used for testing was first stored at 4 °C in a refrigerator. Here, mitochondrial viability was determined at 1.5~3, 3~4.5, 4.5~6, and 6~7.5 h from delivery. Substrate uncoupling inhibitor titration (SUIT) was then performed as previously described [14] (Figure1): For SUIT protocol pyruvate (5 mM), glutamate (10 mM) and malate (2 mM) were added to determine complex I (CI) mediated LEAK respiration, then oxidative phosphorylation (OXPHOS) through CI was stimulated by the addition of ADP(Adenosine Diphosphate) (5 mM). Cytochrome c (10 uM) was then added to test the integrity of the outer mitochondrial membrane. Succinate (10 mM) was added directly after cytochrome c, to elevate OXPHOS through complexes I and II (OXPHOS CI, II). Titrations of CCCP (0.5uM) were also used to investigate the ETS capacity with CI and CII (ETS CI, II). Rotenone (0.5uM) and antimycin A (2.5uM) were then added to inhibit Complexes I and III respectively, with N,N,N,N-Tetramethyl-p-phenylenediamine dihydrochloride (TMPD:0.5 mM) and ascorbate (2 mM) being added to determine OXPHOS through complex IV.
Statistical Analysis
All data represent the results of more than three replicate experiments and are expressed as mean ± standard error. All statistics were completed through the statistical software Graphpad Prism 9.0. One-Way ANOVA was used to compare the means of multiple samples, and P<0.05 indicated that the difference was statistically significant.
![Figure 1: ** Substrate-uncoupler-inhibitor titration (SUIT) protocol. Representative oxygraph traces of oxygen flux relative to tissue mass. The vertical lines show the introduction of various substrates or inhibitors. LEAK refers to non-phosphorylating respiration; OXPHOS CI, CII, and CIV represent oxidative phosphorylation through mitochondrial complexes I, II, and IV, respectively; ETSmax represents electron transfer system maximum capacity; ROX represents non-ETS respiration; PMG represents pyruvate, glutamate, and malate; CYC represents cytochrome c; ROT represents rotenone; SCC represents succinate; CCCP represents the uncoupler, 2-[(3-chlorophenyl)hydrazinylidene]propanedinitrile (CCCP); AMA represents antimycin A; TA represents 1-N,1-N,2-N,2-N-tetramethylbenzene-1,2-diamine;dihydrochloride and ascorbate.](/fulltextimages/12560/fig_1.png)
Figure1: Substrate-uncoupler-inhibitor titration (SUIT) protocol. Representative oxygraph traces of oxygen flux relative to tissue mass. The vertical lines show the introduction of various substrates or inhibitors. LEAK refers to non-phosphorylating respiration; OXPHOS CI, CII, and CIV represent oxidative phosphorylation through mitochondrial complexes I, II, and IV, respectively; ETSmax represents electron transfer system maximum capacity; ROX represents non-ETS respiration; PMG represents pyruvate, glutamate, and malate; CYC represents cytochrome c; ROT represents rotenone; SCC represents succinate; CCCP represents the uncoupler, 2-[(3-chlorophenyl)hydrazinylidene]propanedinitrile (CCCP); AMA represents antimycin A; TA represents 1-N,1-N,2-N,2-N-tetramethylbenzene-1,2-diamine;dihydrochloride and ascorbate.
Results
Respiration of Placental Mitochondria under Different Oxygen Levels
Mitochondrial respiration capacity did not change significantly under the variation of oxygen levels. As shown in (Figure 2), mitochondrial respiration capacity in ambient air (150–160 μM) was not significant differently from when exposed to oxygen levels of 200, 250, and 300 μM in oxidative phosphorylation (OXPHOS) with substrates for Complex I (CI) and Complex II (CII). In addition, the same was observed for the maximum respiration of the electron transfer system (ETSmax). OXPHOS with substrates for Complex IV (CIV), non-phosphorylating respiration (LEAK), and non-electron transfer system respiration remained stable across different supplementary oxygen concentrations. These results indicated that changes in oxygen concentration in the O2K chamber used to detect the respiratory capacity of placental mitochondria did not alter mitochondrial respiratory function. This was sufficiently in line with the manufacturer’s recommendations and extensive literature demonstrating air calibration prior to closure of the system (a procedure that is required).
However, this study was focused on mitochondrial metabolism in the highland adaptation to low oxygen levels. The equipment was used to conduct research at high altitudes 3780 m (meters) since high-resolution respirometry measurements cannot be performed on frozen materials. It was important to keep in mind that compared to low altitudes (below 2500 m), the machine calibration approach utilized here to study the characteristics of mitochondrial metabolism at high altitudes was different. The instrumental background could also be performed at the higher altitude [11, 15]. Altitude calibration schemes for different levels are different, and therefore close communication with engineers was required.
Oxygen was added to the tissue preparation prior to measuring respiratory function to ensure that oxygen did not limit respiratory capacity (especially at high respiratory rates) and to counteract the formation of oxygen diffusion gradients. When studying the effect of oxygen concentration on mitochondrial respiratory function, oxygen was injected into the chamber respiratory fluid via a syringe to limit the oxygen concentration in the chamber before the start of each experiment. Oxygen concentration in the chamber was also decreased with the addition of mitochondrial substrates, uncouplers, and inhibitors. It was found that despite the laboratory being located at an altitude of 2200 m, after O2K calibration, oxygen concentration in the chamber was 150– 160 uM. During the placental tissue experiment, by reducing the detected tissue to 5-10 mg, the experiment could be carried out smoothly without additional increases in oxygen concentration within the chamber. However, for heart and skeletal muscle tissue with high oxygen consumptions, it may be necessary to add enough oxygen to meet mitochondrial oxygen consumption.
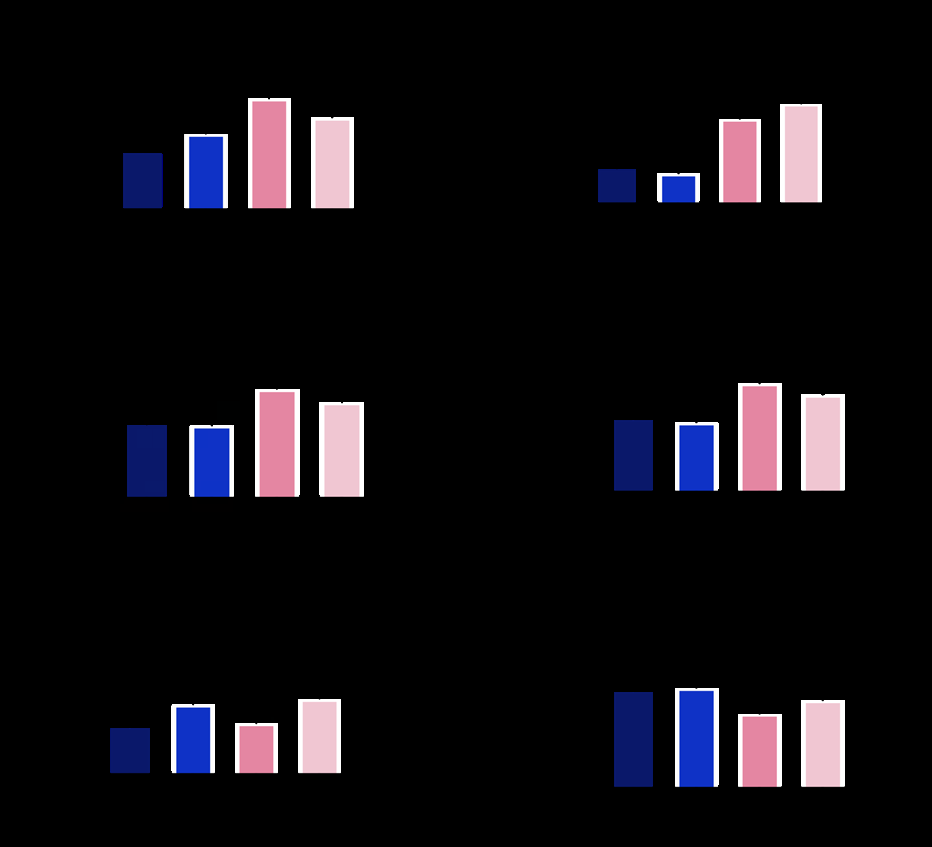
Respiration of Placental Mitochondria in Different Time Points
The mitochondrial respiration capacity changed as time from placental delivery increased. As shown in (Figure 3), there was a significant decrease in OXPHOS(Oxidative phosphorylation) with substrates for mitochondrial CI, CI + CII, and ETSmax after 4.5 h storage in the respiratory medium compared with 3 h. OXPHOS with substrates for CIV, LEAK, and ROX did not significantly change at different time points. Despite being stored in respiratory medium, some activity of the placental mitochondria was impaired after 3h. Considering that it took 1.5 h (from delivery to penetration ready for testing) to prepare the tissue and 1.5 h (from the addition of tissue to the chamber to the final addition of the substrate of complex IV) to test a single mitochondrial respiratory parameter, we recommend testing no more than two parameters per sample so as not to exceed the 3 h time limit.
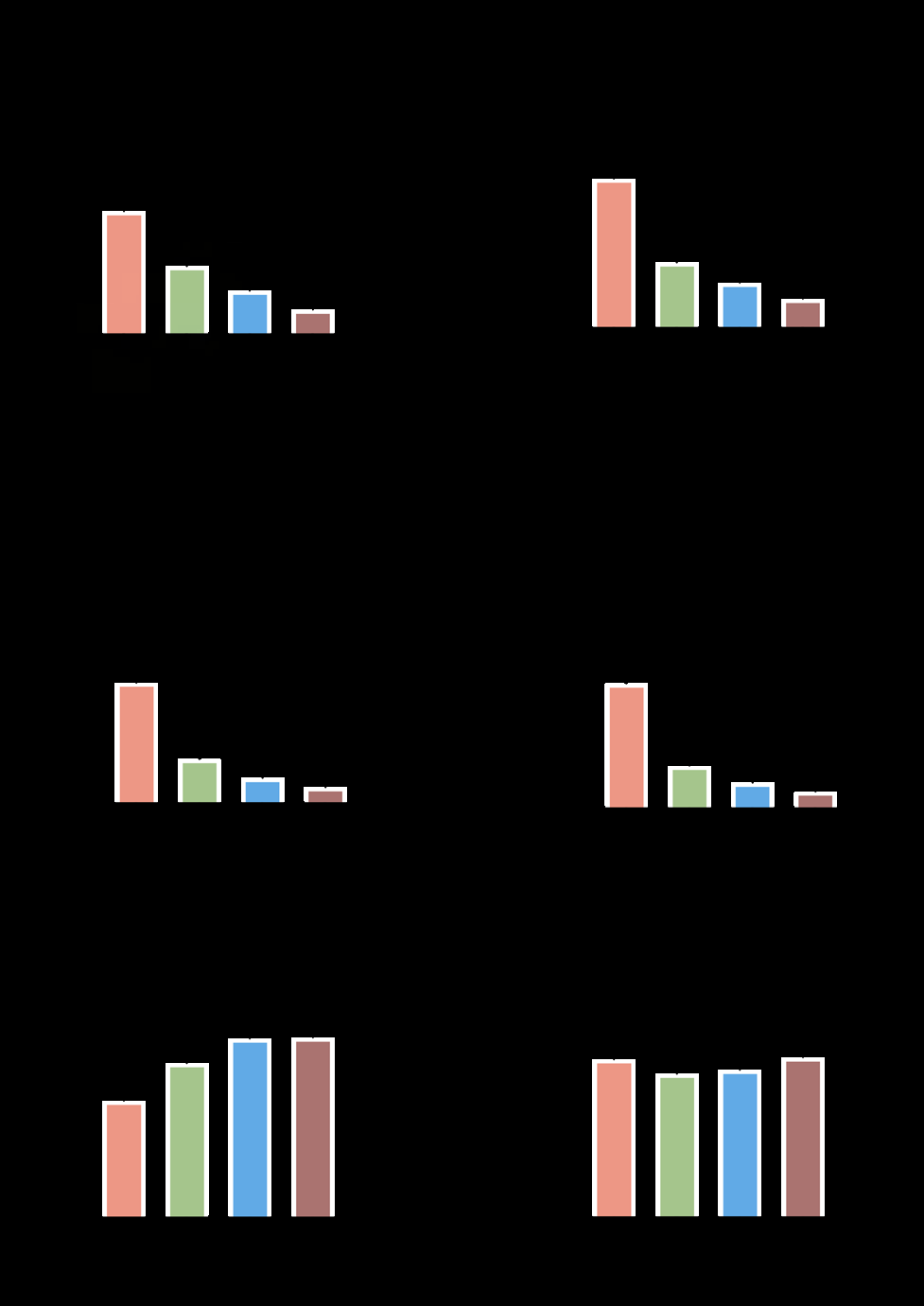
Figure 3: Mitochondrial respiration capacity differs at different time points. Mitochondrial respiration through Complex I, Complex II, Complex IV, Leak Respiration, ETSmax, and ROX were measured at different time points after preparation (1.5~3, 3~4.5, 4.5~6, 6~7.5 h). Asterisk denotes significance between different time points. *P ≤ 0.05, P ≤ 0.01,*P ≤ 0.001, ****P ≤ 0.0001; ns, nonsignificant. Data are mean ± SE.
In conclusion, here we recommend not utilizing placental samples more than 3 h after delivery.
Submission Statement
All authors have read and agree with manuscript content,also in this section is a statement regarding that while this manuscript is being reviewed for this journal, the manuscript will not be submitted elsewhere for review and publication.
Human Ethical Approval Statement
The studies involving human participants were reviewed and approved by Qinghai University Affiliated Hospital Ethics
Committee (P-SL-2020092). The patients/participants provided their written informed consent to participate in this study.
Authors Contribution Statement
TW designed the experiments. HL and NT performed the experiments and HL and TW analyzed the data. HL, NT, MP, TW, and R-lG wrote, read, edited, and approved the final version of manuscript. All authors contributed to the article and approved the submitted version.
Conflict of Interest Statement
There were no conflicts of interest regarding this manuscript. All have read and understood the journal’s guidelines on conflicts of interest and affirm that there are no financial or personal relationships that could potentially influence the objectivity in reporting the research findings.
Funding
This work was supported by the National Natural Science Foundation of China (82060338).
References
-
Maltepe E, Fisher SJ (2015) Placenta: the forgotten organ. Annu Rev Cell Dev Biol 31: 523-552.
-
Fisher J, McKeating D, Pennell E, Cuffe J, Holland O, et al. (2019) Mitochondrial isolation, cryopreservation and preliminary biochemical characterisation from placental cytotrophoblast and syncytiotrophoblast. Placenta 82: 1-4.
-
Burton GJ (2009) Oxygen, the Janus gas; its effects on human placental development and function. Journal of Anatomy 215(1): 27-35.
-
Burton GJ, Charnock-Jones DS, Jauniaux E (2006) Working with oxygen and oxidative stress in vitro. Methods Mol Med 122: 413-425.
-
Soothill PW, Nicolaides KH, Rodeck CH, Campbell S (1986) Effect of gestational age on fetal and intervillous blood gas and acid-base values in human pregnancy. Fetal Ther 1(4): 168-175.
-
Lei J, Zhao M, Li L, Ji B, Xu T, et al. (2022) Research progress of placental vascular pathophysiological changes in pregnancy-induced hypertension and gestational diabetes mellitus. Front Physiol 13: 954636.
-
Hu XQ, Zhang L (2021) Hypoxia and Mitochondrial Dysfunction in Pregnancy Complications. Antioxidants (Basel) 10(3): 405.
-
Mando C, Anelli GM, Novielli C, Panina-Bordignon P, Massari M, et al. (2018) Impact of Obesity and Hyperglycemia on Placental Mitochondria. Oxid Med Cell Longev 2018: 2378189.
-
Purhonen J, Grigorjev V, Ekiert R, Aho N, Rajendran J, et al. (2020) A spontaneous mitonuclear epistasis converging on Rieske Fe-S protein exacerbates complex III deficiency in mice. Nat Commun 11(1): 322.
-
Rajendran J, Purhonen J, Tegelberg S, Smolander OP, Morgelin M, et al. (2019) Alternative oxidase- mediated respiration prevents lethal mitochondrial cardiomyopathy. EMBO Mol Med 11(1): e9496.
-
Horscroft JA, Kotwica AO, Laner V, West JA, Hennis PJ, Levett DZH, et al. (2017) Metabolic basis to Sherpa altitude adaptation. Proc Natl Acad Sci U S A 114(24): 6382-6387.
-
Khera A, Dong LF, Holland O, Vanderlelie J, Pasdar EA, et al. (2015) Selenium supplementation induces mitochondrial biogenesis in trophoblasts. Placenta 36(8): 863-869.
-
Sagrillo-Fagundes L, Clabault H, Laurent L, Hudon- Thibeault AA, Salustiano EM, et al. (2016) Human Primary Trophoblast Cell Culture Model to Study the Protective Effects of Melatonin Against Hypoxia/reoxygenation- induced Disruption. J Vis Exp (113): 54228.
-
Holland OJ, Hickey AJR, Alvsaker A, Moran S, Hedges C, et al. Changes in mitochondrial respiration in the human placenta over gestation. Placenta 57: 102-112.
-
Liu H, Tenzing N, van Patot MT, Qile M, Ge RL, et al. (2021) Enhanced Placental Mitochondrial Respiration in Tibetan Women at High Altitude. Front Physiol 12: 697022.
- Postpartum Maternal Mental Health - A Narrative Review
- Beta HCG in Cervico-Vaginal Secretion as a Predictor of Preterm Delivery
- Successful Management of Mid Trimester Foetal Death with Major Placenta Previa by Expectant Management Followed by Induction of Labour
- To Evaluate the Expression of Egr2 Gene in Term Low Birth Weight Newborns
- Impact of Maternal Obesity on Maternal and Foetal Outcomes: A Prospective Cohort Study from Northern India
- ‘’Benefit of Pulsatile GnRH Therapy in Treatment of Functional Hypothalamic Amenorrhea (FHA) and Congenital Hypogonadotropic Hypogonadism(CHH) in Infertile Patients Over Canonical Gonadotropins with IVF –A Short Communication’’