Rho Kinase Inhibitors for Glaucoma and Optic Neuropathy Treatment: Enzyme Activity, Cell Effects, IOP-Lowering and Neuroprotection Perspectives
Rho kinases, (Rho-Associated Coiled-Coil Kinases; ROCKs) are ubiquitous enzymes found inside all cells of the body. Consequently, ROCKs are involved in numerous cellular functions such as cell contraction, cell migration and proliferation. As such, their over-expression / activation has been implicated in diseases ranging from diabetes, erectile dysfunction, hypertension, asthma, cancer, cardiovascular hypertrophy, inflammatory diseases, pulmonary hypertension and so on. In the eye, ROCKs are involved in the development of glaucoma, Fuchs corneal dystrophy, diabetic retinopathy and age-related macular degeneration. Inhibitors of ROCKs have proven useful drugs to treat the afore-mentioned disorders, and in particular they are effective intraocular pressure lowering agents with added neuroprotective activities. This review addresses the latter aspect of ROCK physiology, pharmacology and pathology, and the clinical utility of ROCK inhibitors towards preservation of eyesight from the ravages of glaucomatous optic neuropathy.
Najam A Sharif *1-7
Glaucoma; RGC: Retinal Ganglion Cells; ROCK: Rho Kinase; SC: Schlemm’s Canal; TM: Trabecular Meshwork.
Introduction
Optic nerve damage leading to visual field loss is caused by several different forms of glaucoma, an optic neuropathy in which the death of retinal ganglion cells (RGCs) and their axons are end-stage characteristic features [1, 2, 3, 4]. While the exact factors involved and the natural history development of glaucomatous optic neuropathy (GON) are incompletely understood, accumulating evidence points to a multifactorial assault on various elements of the eye-brain axis [2, 3, 4]. This encompasses both structural and functional components. Fundamentally, it appears that chronically elevated intraocular pressure (IOP) is a principal culprit and trigger in the onset of primary open-angle glaucoma (POAG) [1, 2, 3, 4, 5, 6], while primary angle-closure glaucoma (PACG) results due to sudden acutely rising IOP. Even though, ocular hypertension (OHT) is a major risk factor for POAG, advancing / old age, low intracranial fluid pressure (ICFP), family history/African or Hispanic ancestry, IOP spikes [7], low retinal blood flow [8] and defects in axonal transport mechanisms within the optic nerve [9] also contribute to the visual impairment and eventual blindness in glaucoma patients [1, 2, 3, 4, 5]. Since glaucomatous optic nerve damage and visual field defects occur asymptomatically, they generally progress slowly over decades and the patient remains oblivious to that damage that is occurring to their RGCs and their axons within the optic nerve and various brain structures [1, 2, 3, 4, 5, 10, 11, 12, 13, 14, 15, 16, 17]. Hence, early detection, diagnosis and treatment of different forms of glaucoma is critical to restricting retinal/brain damage, and thus for preserving eyesight of those that suffer from GON [18, 19]. It is predicted that by 2040, there will be >112 million patients on our planet afflicted with glaucoma [20]. Thus, it is imperative that novel suitable treatment modalities be found and approved for effective, efficient and optimal medical management of glaucomas [21, 22, 23].
Pathological Aspects of OHT Associated with POAG
As is well documented, the anterior chamber (ANC) of the eye contains aqueous humor (AQH) which is formed and released by non-pigmented ciliary epithelial cells of the ciliary body[24] (Figure 1). The AQH provides oxygen and nutrients to the avascular tissues/cells lining the ANC and removes metabolic waste from the ANC as it flows towards the drainage pathway, the trabecular meshwork (TM) and the Schlemm’s canal (SC) [25, 26, 27, 28, 29] (Figure 1). The TM comprises arrays of beams of connective tissue possessing a core of elastic and collagen fibers which are covered by monolayers of endothelial-like TM, with loose extracellular matrix (ECM) laid down between neighboring beams [26, 27, 29]. Recent work has identified several different type of cells making up the TM which suggests multiple functionalities [30, 31]. The cribriform region is the outermost non-lamellated juxtacanalicular tissue (JCT) which is made up of a loose formless ECM web. This outermost cribriform layer is continuous with the endothelial lining of SC which drains the AQH into the venous circulation [28, 32].
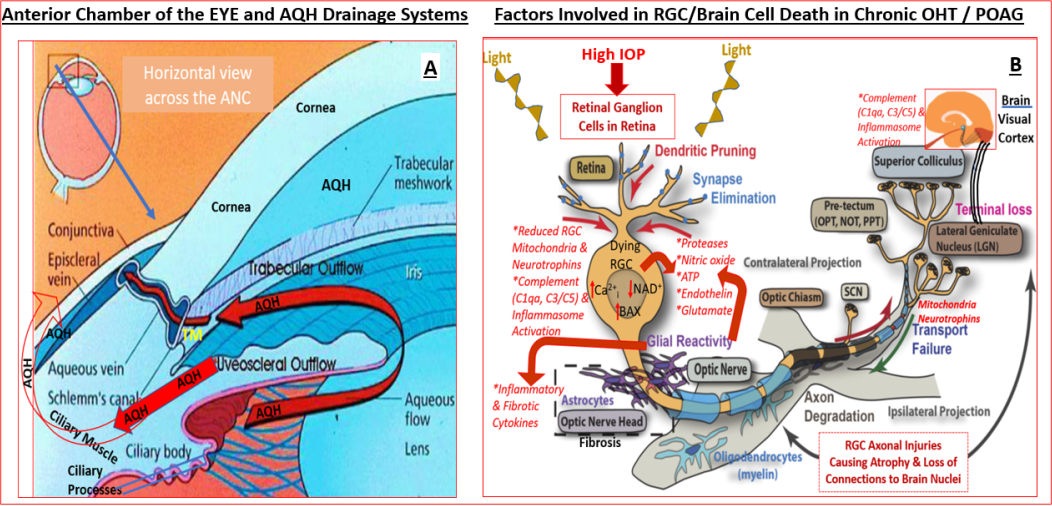
The aging process, coupled with other local pathological factors in POAG such as aberrant release of damaging cytokines like tissue growth factor-β (TGF-β) [33, 34] and connective tissue growth factor cause deposition of excess collagen and fibronectin [35, 36] in and around the TM thereby obstructing the efficient drainage of the AQH from the ANC. Normally, the TM actively produces and secretes proteases that digest the accumulated extracellular matrix (ECM) and the TM cells phagocytose and eliminate the resultant components of ECM. However, age-related stiffness [26, 37] and senescence of TM cells [38] and oxidative stress [39, 40, 41] and mitochondrial damage [42, 43] causes a diminution in ECM clearance [44] and clogging of the TM/ SC structures, with resultant accumulation of excess AQH in the AC and an increased IOP. Such chronically elevated IOP propagates and mechanically stretches, distorts [45] and damages the vulnerable tissues at the back of the eye at the optic nerve head (ONH) [46, 47, 48] and lamina cribosa (LC) [11, 49, 50] regions where the RGC axons gather before leaving the eyeball as part of the optic nerve connecting the retina to the brain. The ensuing local inflammation [51, 52, 53], vasoconstriction of retinal blood vessels [8] and RGC axonal constriction [9, 54, 55, 56], and some vascular leakage leads to release of cytokines [51], chemokines, proteases [57] and other substances like endothelin [56, 58] (Figure 1). Additionally, immune cells migrate to the retina and activated microglia [18, 59, 60, 61] release copious amounts of nitric oxide [46, 47] and other damaging factors [56, 58] and deposit complement components in the retina [61] and brain structures associated with visual perception [18, 62]. These events and agents conspire to cause retinal ischemia, restrict growth factor and mitochondrial transport from the brain to the RGCs which ultimately cause RGC axon atrophy and RGC apoptosis [18, 63, 64, 65] (Figure 1). If left untreated this leads to peripheral vision loss and can eventually result in blindness [1, 2, 3, 4, 5, 14].
Current and Future Treatment Modalities for OHT/POAG
OHT appears to be a major root cause of visual impairment in POAG and many other forms of glaucoma including normotensive glaucoma (NTG) [66, 67]. Several clinical trials have demonstrated the benefits of lowering IOP to slow down the progression of visual damage and loss of vision [68, 69, 70, 71, 72]. Indeed, strong evidence supports the concept that lowering of IOP whether by topical drugs [14, 68, 69, 70, 71, 72, 73] or after surgical intervention [74, 75] results in both structural and functional improvements. Thus, the removal of excess AQH from the ANC of the eye reverses LC displacement [73], reduces retinal nerve fiber layer thinning [76], and prevents RGC loss [75] coupled with enhancement of visual fields in the OHT/glaucoma patients. Thus, for every 1mmHg reduction in IOP there is a 10-13% reduction in the progression of GON [1, 2, 3, 4, 5, 14, 66, 67, 68, 69, 70].
As a result of the afore-mentioned positive effects of IOP-lowering, many drugs, AQH drainage shunts, and various surgical techniques to relieve the pressure in the eye have been developed and approved by health agencies worldwide. Thus, topical ocularly administered drugs to reduce AQH production (e.g. beta-blockers: timolol, betaxolol; carbonic anhydrase inhibitors: dorzolamide, brinzolamide; alpha-adrenergic agonists: brimonidine and apraclonidine) [1, 3, 24], to enhance AQH outflow from the ANC via the TM/SC (e.g. muscarinic agonists: pilocarpine; rho kinase [ROCK] inhibitors: ripasudil, netarsudil; FP- receptor prostaglandin agonists (latanoprost, travoprost, tafluprost [76, 77, 78]; EP2-receptor agonists: omidenepag isopropyl [79, 80, 81]) , and drugs to promote drainage of AQH via the uveoscleral pathway (e.g. FP-receptor prostaglandin agonists: latanoprost, travoprost, tafluprost [75, 76]; EP2- receptor agonists: omidenepag isopropyl [79, 80, 81]), and various conjugates (e.g. latanoprostene bunod [82]) and combination products [83, 84] are prescribed to lower and control IOP [1, 2, 3, 4] (Table 1). ROCK inhibitors (e.g. ripasudil [85, 86, 87]; netarsudil [88, 89]; and others) that promote AQH outflow via the TM/SC conventional pathway and also by reducing episcleral venous pressure will be discussed in more detail below. Many other classes of investigational compounds with ocular hypotensive activity have also been studied in various animal models of acute and chronic OHT and POAG [3, 90, 91, 92] (Table 2).
| Brand Name of Drug & Year of Clinical Introduction/ FDA or EMEA or Japan Approval (where known) | Generic Name & Drug Type. ( IOP reduction achieved in OHT/POAG patients) | Dosage Type (%, w/v) | Topical Ocular Dosing Frequency | Mechanism of Action to Lower IOP | Some Side-Effects/ Adverse reactions |
|---|---|---|---|---|---|
| Approved Enhancers of Conventional Outflow (TM/SC Pathway) of AQH to Lower IOP | |||||
| Isopto carpine (1974); Pilopine | Pilocarpine (muscarinic receptor agonist) | 1%, 2%, 4% ; 4% Gel | 1 drop 2-4-times daily; single application of gel across the eye | Enhances conventional (TM) outflow of AQH | Brow-ache, miosis; accommodative change, eye irritation, eye pain, blurred vision, and/or visual impairment, potential tachycardia. |
| Isopto Carbachol | Carbachol (muscarinic receptor agonist) | 1.5, 3% solution | 1-2 drops up to 3-times daily | Enhances conventional (TM) outflow of AQH | Brow-ache, miosis, accommodative change, eye irritation, eye pain, blurred vision, and/or visual impairment, potential tachycardia. |
| Glanatec (2014 Japan) | Ripasudil (Rho kinase [ROCK] inhibitor) (3.5-4.5 mmHg IOP reduction) | 0.4% solution | Enhances conventional (TM) outflow of AQH | Conjunctival hyperemia, allergic conjunctivitis, blepharitis, punctate keratitis | |
| Rhopressa (2017) | Netarsudil (Rho kinase [ROCK] inhibitor) (5 mmHg IOP reduction) | 0.02% solution | 1 drop daily | Enhances conventional (TM) outflow of AQH; also decreases episcleral veinous pressure | Conjunctival hyperemia, corneal verticillata, instillation site pain, and conjunctival hemorrhage. |
| Approved Inhibitors of AQH Production to Lower IOP | |||||
| Timoptic (1978) Timoptic-XE Gel | Timolol (beta- adrenoceptor antagonist); | 0.25%, 0.5% solution or gel-forming solution | 1 drop 1-2-times daily | Reduces production of AQH from CB | Signs and symptoms of ocular irritation, (e.g. burning, stinging, itching, tearing, redness), conjunctivitis, blepharitis, keratitis, dry eyes, decreased corneal sensitivity, blurred vision, corneal erosion. Visual disturbance, including refractive changes |
| Betoptic (1985) | Betaxolol (beta- 1-selective adrenoceptor antagonist); | 0.25% suspension; 0.5% solution | 1 drop 2-times daily; 1-2 drops twice daily | Reduces production of AQH from CB | Transient ocular discomfort, Decreased corneal sensitivity, erythema, itching sensation, corneal punctate keratitis, anisocoria, blurred vision, foreign body sensation, tearing, dryness of eyes, inflammation, discharge, ocular pain, decreased visual acuity, crusty lashes and photophobia; Bradycardia, heart block; Pulmonary distress characterized by dyspnoea, bronchospasm, thickened bronchial secretions, asthma and respiratory failure; Insomnia, dizziness, vertigo, headaches, depression, lethargy |
| Alphagan (1996) | Brimonidine (2-6 mmHg IOP reduction) | 0.15%, 0.2% solution | 1 drop 3-times daily | Reduces production of AQH from CB and enhances UVS AQH outflow | Allergic conjunctivitis, conjunctival hyperemia, and eye pruritis; local ocular hypersensitivity; blurred vision, burning sensation of eyes, drowsiness, eye headache, stinging of eyes, foreign body sensation |
| Iopidine (1987) | Apraclonidine | 0.5% solution | 1-2 drops 3-times daily | Reduces production of AQH from CB | Hyperemia (redness), itching, tearing of the eye, Blurred vision or change in vision, chest pain, clumsiness or unsteadiness, depression, dizziness, eye discharge, irritation, or pain, irregular heartbeat |
| Trusopt (1994) | Dorzolamide (carbonic anhydrase inhibitor; | 2% solution | 1 drop 3-times daily | Reduces AQH generation by the CB | Transient bitter taste and superficial punctate keratitis, eye irritation, burning, stinging, and ocular discomfort; blurred vision, excessive tearing, dry eyes, and increased sensitivity to light, |
| Azopt (1998) | Brinzolamide (carbonic anhydrase inhibitor; | 1% suspension | 1 drop 3-times daily | Reduces AQH generation by the CB | Temporary blurred vision, bitter/ sour/unusual taste, dry eyes, temporary discomfort, itching, redness of the eye, foreign body sensation, eye discharge, and headache. |
| Approved Stimulators of UVS Outflow of AQH to Lower IOP | |||||
| Xalatan (1996) | Latanoprost (FP- prostaglandin receptor- selective agonist; | 0.005% solution | 1 drop at bedtime | Enhances AQH outflow via the UVS pathway and some via TM/SC pathway | Blurred vision, burning, stinging, itching, hyperemia, foreign body sensation, changes in eyelash number/color/length/thickness, iridial darkening, (pigmentation), periocular skin darkening, deepening of eyelid sulcus (loss of periorbital fat), dry eye, eyelid crusting and discomfort, increased sensitivity to light. |
| Travatan (2001) | Travoprost (FP- prostaglandin receptor- selective agonist; | 0.004% solution | 1 drop at bedtime | Enhances AQH outflow via the UVS pathway and some via TM/SC pathway | Blurred vision, burning, stinging, itching, hyperemia, foreign body sensation, changes in eyelash number/color/length/thickness, iridial darkening, (pigmentation), periocular skin darkening, deepening of eyelid sulcus (loss of periorbital fat), dry eye, eyelid crusting and discomfort, increased sensitivity to light. |
| Lumigan (2001) | Bimatoprost (FP- prostaglandin receptor- selective agonist; | 0.03% solution | 1 drop at bedtime | Enhances AQH outflow via the UVS pathway and some via TM/SC pathway | Increased conjunctival hyperemia, darkening of eyelids, increased thickening and number of eyelashes, dry eye, eye irritation, eye itching. Hirsutism (a condition of hair growth on parts of the body normally without hair). |
Tafluprost (FP- prostaglandin receptor- selective agonist;
Taflotan (2008 Japan) Zioptan (2012 USA)
0.0015% solution 1 drop at bedtime Unoprostone (FP- prostaglandin receptor agonist;
0.15% solution 1 drop twice daily Rescula (2000)
Omidenepag Isopropyl (EP2-receptor selective non- prostaglandin agonist;
0.002% solution 1 drop daily Eybelis (2018 Japan)
Some Approved Combination Products for Lowering IOP
Cosopt (1998) Dorzolamide + Timolol 2% + 0.5% 1 drop 2-times daily Combigan (2007) Brimonidine + Timolol 0.2% + 0.5% 1 drop every 12 hrs Simbrinza (2013) Brinzolamide + Brimonidine 1% + 0.2% 1 drop 3-times daily Roclatan (2019) Netarsudil + Latanoprost 0.02% + 0.005% 1 drop daily Xalacom Latanoprost + Timolol 0.005% + 0.5% 1 drop daily Duotrav Travoprost + Timolol 0.004% + 0.5% 1 drop daily Ganfort Bimatoprost + Timolol 0.03% + 0.5% 1 drop daily Ocular surface burning, stinging, irritation, hyperemia, foreign body sensation, dry eyes, watering eyes, iridial darkening, periocular skin darkening, abnormal eyelash growth, and increased sensitivity to light.
Enhances AQH outflow via the UVS pathway and some via TM/SC pathway Eye burning, stinging, dry eyes, itching, increased length of eyelashes, and injection; iridial darkening, blepharitis, cataract, conjunctivitis, corneal lesion, discharge from the eye, eye hemorrhage, eye pain, keratitis, irritation, and photophobia.
Enhances AQH outflow via the UVS pathway and some via TM/SC pathway Enhances AQH outflow via the UVS pathway and via TM/SC pathway Transient conjunctival hyperemia, corneal thickening.
Reduce AQH production from CB
Combination of side-effects from both drugs Reduce AQH production from CB
Combination of side-effects from both drugs Reduce AQH production from CB
Combination of side-effects from both drugs Enhancement of AQH outflow via TM/SC and UVS pathways Combination of side-effects from both drugs Enhancement of AQH outflow and by inhibiting AQH production Combination of side-effects from both drugs Enhancement of AQH outflow and by inhibiting AQH production Combination of side-effects from both drugs Enhancement of AQH outflow and by inhibiting AQH production Combination of side-effects from both drugs
| Taflotan + Timolol | Taflotan + Timolol (>13 mmHg IOP reduction; 40% lowering) | 0.0015% + 0.5% | 1 drop daily | Enhancement of AQH outflow and by inhibiting AQH production | Combination of side-effects from both drugs |
|---|---|---|---|---|---|
| Other Products for Lowering IOP | |||||
| Vyzulta (2017) | Latanoprostene Bunod (conjugate of latanoprost and an NO-donor agent) | 0.024% solution | 1 drop at bedtime | Enhances AQH outflow via the UVS pathway and via TM/SC pathway | Eye discomfort / irritation, hyperemia, temporary blurred vision, increase in eyelash number/ length/thickness and darkening of the eyelashes/eyelids and iris. |
| Durysta Implant (2020) | Intracamerally injected sustained- delivery biodegradable polymer containing bimatoprost | Not applicable | Once implanted into the ANC of the eye (intracameral injection), the drug elutes off the implant over 6-months. | Enhances AQH outflow via the UVS pathway and via TM/SC pathway | Conjunctival hyperemia, foreign body sensation, eye pain, photophobia, conjunctival hemorrhage dry eye, eye irritation increased IOP, corneal endothelial cell loss, vision blurred, iritis, headache. |
Table 2: Approved ocular hypotensive drugs to treat chronic OHT and POAG with highlighted ROCK inhibitors.
| Compound Classes | Drug Candidates | Reported or Potential Mode(s) Of Action |
|---|---|---|
| Conventional Outflow (via TM pathway) Promotors | ||
| Inhibitors of chloride transport | Ticrynafen; Ethacrynic acid; Indacrinone | Inhibition of Na+-K+-Cl--transporter activity in the TM changes cell shape & volume and thus AQH efflux is increased |
| Kinase inhibitors | Chelerythrine; Staurosporin; | Modification of actomyosin contractility that leads to changes in actin cytoskeleton of TM and this leads to AQH efflux; direct relaxation of the TM may also be involved |
| LIM-K inhibitors (e.g. LX7101); | ||
| Myosin-II ATPase inhibitor: Blebbistatin. | ||
| Src kinase inhibitor | ||
| Rho Kinase (ROCK) Inhibitors | Fasudil; Y-27632; AMA0076; ITRI-E-212 | Modification of actomyosin contractility that leads to changes in actin cytoskeleton of TM and this leads to AQH efflux; direct relaxation of the TM may also be involved. |
| Marine macrolids | Latrunculins A and B; Bumetanide; Swinholide | Promote sequestration of actin monomers and dimers in TM; cause cell TM shape change and thus AH efflux |
| Guanylate cyclase activators | Natriuretic peptides and constrained cyclic peptides: ANP; CNP; TAK-6 | Type-A and type-B receptor activation leads to cGMP production, TM relaxation and AQH efflux via TM. |
| NO Donors | Sodium nitroprusside; Hydralazine; 3-morpholinosyndnonimine; (S)- nitrosoacetylpenicillamine; NCX-125 | NO activates intracellular soluble guanylate cyclase to increase cGMP production, TM relaxation and AQH efflux via TM. |
Table 3: Approved ocular hypotensive drugs to treat chronic OHT and POAG with highlighted ROCK inhibitors.
Soluble guanylate cyclase activators IWP-953; MGV354;
k-opioid receptor agonists Bremazocine; Dynorphin Release natriuretic peptides and thus raise cGMP in TM leading to its relaxation & thus AQH efflux Cannabinoid receptor agonists WIN55212-2; CP55940; SR141716A Receptor stimulation opens BKC-channels and relaxes TM which then causes AQH efflux via TM and SC Serotonin-2 receptor antagonists ketanserin Local production of MMPs; ECM degradation; stimulation of AQH efflux via TM/SC Autotaxin/ Lysophosphatidic acid inhibitors Aiprenon Promotion of AQH egress from TM/SC pathway Uveoslceral Outflow promotors (via CM bundles and sclera) EP2- and EP4- PG-receptor agonists AL-6598 ; Butaprost ; ONO-AE1-259-01; PF- 04217329 ; PF-04475270 These compounds directly activate intracellular soluble guanylate cyclase to increase cGMP production, TM relaxation and AQH efflux via TM.
Receptor activation increases cAMP that relaxes CM & TM; EP2 agonists also cause release of MMPs that breakdown ECM (“clog”) around CM bundles and within sclera thus causing UVS outflow of AQH Contraction / relaxation of CM and TM by activation of 5HT2 receptors. May also release MMPs and/or PGs or other local mediators that promote CM remodeling and thus promote UVS outflow Serotonin-2 (5HT-2) receptor agonists (R)-DOI; a-methyl-5HT; AL-34662 B2-receptor activation causes PI hydrolysis production of IPs and DAG; cause PG release and release of MMPs that digest ECM and this promote UVS outflow in cynomolgus monkey; conventional outflow also stimulated in isolated bovine /porcine anterior eye segments Dual pharmacophore PGs FP/EP3 receptor agonist (ONO-954) Promote UVSC outflow Inflow inhibitors (reduce AQH production) Bradykinin B2-receptor agonists Bradykinin; FR-190997; BKA278 Chloride channels inhibitors 5-nitro-2-(3-phenylpropylamino)-benzoate (NPPB) Ion flux of CP NPE cells causes reduction of AQH formation Dopamine receptor agonists PD128907; CHF1035; CHF1024; SDZ GLC-756; (S)-(-)-3-hydroxyphenyl)-N-n- propylpiperidine (3-PPP) Na+-K+-ATPase inhibitors Ouabain; Digoxin analogs Ciliary process Na+-K+-ATPase inhibited leading to inhibition of AQH production Aquaporin Inhibitors Various aromatic sulfonamides and dihydrobenzofurans Inhibit release of NE & prevent AQH production Other IOP-lowering agents Inhibit release of NE & prevent AQH production; may also release natriuretic peptides
| Mas receptor stimulator Angiotensin-II receptor antagonists | DIZE via ACE-2 activation | Prevent ECM (including TGFβ) accumulation (outflow stimulation ?) |
|---|---|---|
| CS-088 | Various mechanisms of action; not robust IOP- lowering | |
| Ca2+-channel inhibitors | Lomerazine; Nivaldipine; Nifedipine; Nimodipine; Verapamil; Brovincamine; Iganidipine | Enhance retinal blood flow; some may lower IOP; work well in normal tension glaucoma patients |
| Alpha-adrenergic receptor antagonists | Oxymetazoline; 5-methylurapidil; Ketanserin | Work mostly via outflow mechanism but this needs to be defined |
| ATP-sensitive K+-channel activators | Cromakalim; Levocromakalim; CKPL1 | Purported MOA involving episcleral veinous pressure modulation |
Table 4: Recently discovered ocular hypotensive agents and their potential modes of action in animal models of OHT / POAG.
Of the approved drugs to treat these diseases, FP-receptor prostaglandin agonists and recently approved (in Japan) EP2- receptor agonist (omidenepag isopropyl) represent first-line treatment modalities. When all drug classes fail to lower and/or control IOP, surgery [74, 75, 93, 94] and implantation of AQH microshunt devices [95, 96] with or without topical ocular medications are required to remove excess AQH from the ANC to lower and control eye pressure. Since patients who continue to lose vision also include those with normal IOPs (normotensive glaucoma; NTG), further reduction of IOP is usually needed. In future, in addition to reducing IOP, patients with all forms of glaucoma will require drugs and other combinatorial neuroprotective / cytoprotective agents and rejuvenating treatments that can reduce RGC/RGC axons loss and thus protect the retina-brain axis components to preserve eyesight [4, 21, 22, 23, 64, 97, 98].
Why are ROCK Inhibitors Important for OHT/Glaucoma Treatment?
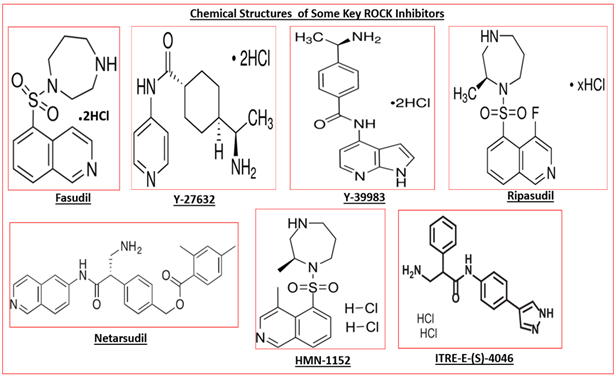
Due to the important and beneficial roles of AQH perfusing through the AC of the eye, drugs or treatment options which promote AQH egress via the TM/SC system to lower IOP are preferred over those that prevent production of AQH. Until recently, the only approved ocular hypotensive drugs that specifically helped remove excess AQH from the ANC of the eye via the conventional TM/SC pathway are pilocarpine and to a lesser extent brimonidine. Due to the local ocular side-effects of both drugs (i.e. brow-ache, lens accommodative issue, eye allergy, etc.), new drugs that are TM/SC AQH drainage promoters have been actively sought. ROCK inhibitors such as fasudil, HMN-1152, Y-27632,Y-39983, ripasudil, netarsudil, AMA-0076, ITRI-E-(S)4046, (Tables 1 & 2) are such drugs, of which only ripasudil and netarsudil have been approved for OHT/POAG treatment thus far.
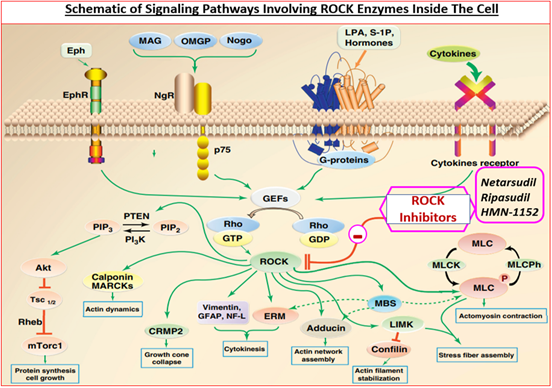
ROCKs belong to a family of serine-threonine kinase enzymes. ROCK-1 (ROCK-β) and ROCK-2 (ROCK-α), sometimes also labeled ROCK-I and ROCK-II, have been discovered and they share a 65% amino acid sequences in common and 92% homology within their functional active enzymic region. Each ROCK sub-type is composed of 5 domains, a catalytic kinase domain at the N-terminus, followed by a central coiled-coil domain containing a rho-binding domain and a C-terminal pleckstrin-homology domain including an internal cysteine- rich domain [88, 99]. While all ROCKs primarily reside in the cytoplasm, they also migrate to the nucleus and membrane depending on the extra- and intra-cellular signaling (Figure 2). ROCK-1 appears preferentially expressed in kidney, liver, spleen and testis, whereas brain, heart, lung, skeletal muscle and the eye appear enriched in ROCK-2. ROCKs phosphorylate different substrates including LIM kinase, myosin light chain (MLC) and MLC phosphatase which regulate actin filament arrangement and contractility [86, 87, 88, 99] (Figure. 2). Due to the relative ubiquity of ROCKs in cells and tissues of the body they are involved in many functions / dysfunctions [88, 99]. In the eye, TM and ciliary muscle contain relatively high expression of ROCKs, and these enzymes appear to be higher in the ONH of the glaucomatous eye than in normal eyes of patients which has implications for neurodegeneration within the retina [100].
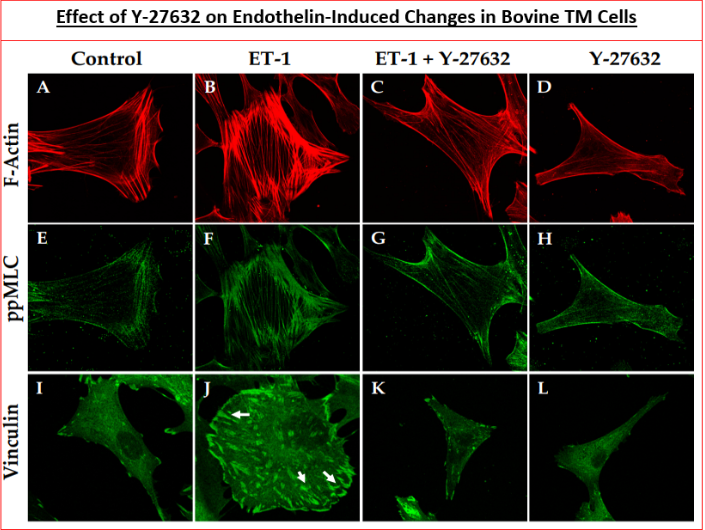
Many studies have shown that ROCK inhibitors (Figure 3) that actively compete for the catalytic ATP-binding site (Figure 4; Table 3) [101, 102, 103] dramatically inhibit actomyosin- driven cell contractility [104, 105, 106, 107], thereby causing relaxation of the cells (Figure 4), disruption of filamentous actin and loss of focal contacts within microfilaments (Figure 5) [89, 104, 105]. Additionally, inhibitors of ROCK block MLC phosphorylation, promote myosin II activity, increase the actin depolymerization and decrease cell-ECM adherence (Figure 5) [89, 104, 105, 106, 107]. Consequently, microtubule bundles are assembled thus generating strong tensile forces, which then induce ECM remodeling within TM/SC. Thus, ROCK inhibitors cause cell relaxation, expand ECM spaces thereby increasing the paracellular fluid flow across inner wall of Schlemm′s canal and JCTs and thus promoting AQH outflow via the TM (Figure 6) [107, 108, 109, 110, 111]. Conversely, substances that activate Rho GTPases such as endothelin-1, sphingosine-1-phosphate, TGF-β2 and lysophosphatidic acid (LPA), decrease the AQH outflow from TM/SC [107, 108, 109, 110, 111]. In enhancing TM/SC-based outflow of AQH from the ANC of the eye in normotensive and ocular hypertensive rats, rabbits, Cynomolgus monkey (and human eyes), ROCK inhibitors lower IOP for over several hours in numerous species (Figures 7 & 8) [89, 102, 103, 107, 108, 109, 110, 111]. A prominent new feature of one particular ROCK inhibitor, netarsudil, is the inherent norepinephrine transporter (NET) inhibitory activity and its reduction of episcleral venous pressure, which contribute to its overall ocular hypertensive actions [88, 89]. A fixed-dose combination of netarsudil and the FP- receptor agonist latanoprost yields additive IOP-lowering in animals and OHT/POAG patients [112, 113].
| Compound | [γ-33P]-ATP-Based Assay (IC , nM) 50 | IMAP Fluorescence Polarization-Based Assay (IC , nM) 50 |
|---|---|---|
| O O S N N N Fasudil | 1690 ± 185 nM (N = 10) | 291 ± 43 nM (N = 9) |
| O O N S N N H-7 | 2341 ± 395 nM (N = 5) | 913 ± 644 nM (N = 3) |
| O N H NH2 Y-27632 | 2802 ± 865 nM (N = 3) | 797 ± 206 nM (N = 3) |
| N OO N S N A | 3463 ± 1800 nM (N = 4) | 270 ± 113 nM (N = 3) |
| N NSOO N B | 485 ± 207 nM (N = 3) | 108 ± 53 nM (N = 2) |
| O N N H O N O S NNH2 C | 1512 ± 704 nM (N = 4) | 2007 ± 85 nM (N = 3) |
| N O O N S N D | 2625 ± 307 nM (N = 4) | 2390 ± 1260 nM (N = 3) |
| N O N S O N HMN-1152 | 47 ± 14 nM (N = 4) | 30 ± 14 (N = 4) |
| O O N S N N HA-135 | 6702 ± 900 nM (N = 2) | 3900 ± 2100 (N = 2) |
Table 5: ROCK-2 Inhibition Constants (IC50) Obtained from Two Different Assay Systems in vitro.
Data in the above table are mean ± SEM; N = the number of independent assays conducted using each type of assay system. Compounds A-D represents new ROCK inhibitors.
Literature ROCK-2 inhibition constant (IC50) data for other ROCK inhibitors are: ripasudil = 9.0 nM; netarsudil= 1 nM; AMA0076= 2.3 nM; ITRI-E-(S)-4046 = 3.2 nM; Y-39983 = 3.6
nM. Table 3 illustrates the relative potencies and selectivities of various ROCK inhibitors for ROCK-1, ROCK-2 and other protein kinases. Note that potency is inversely related to IC50 values.
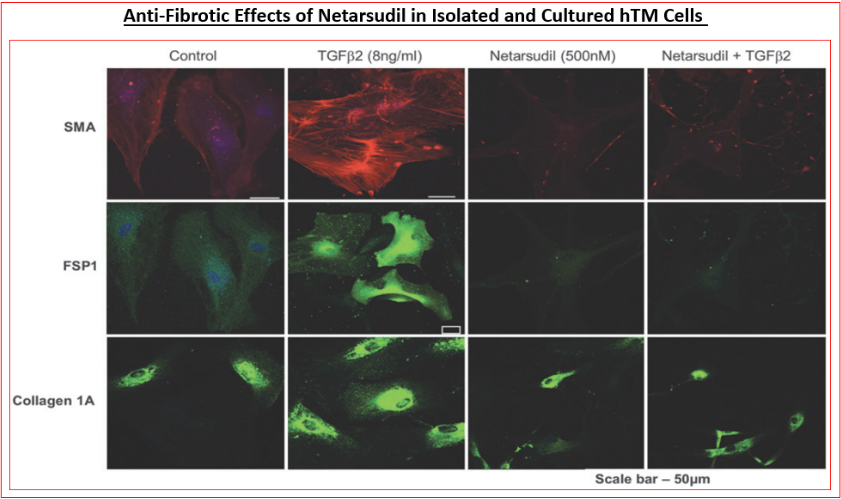
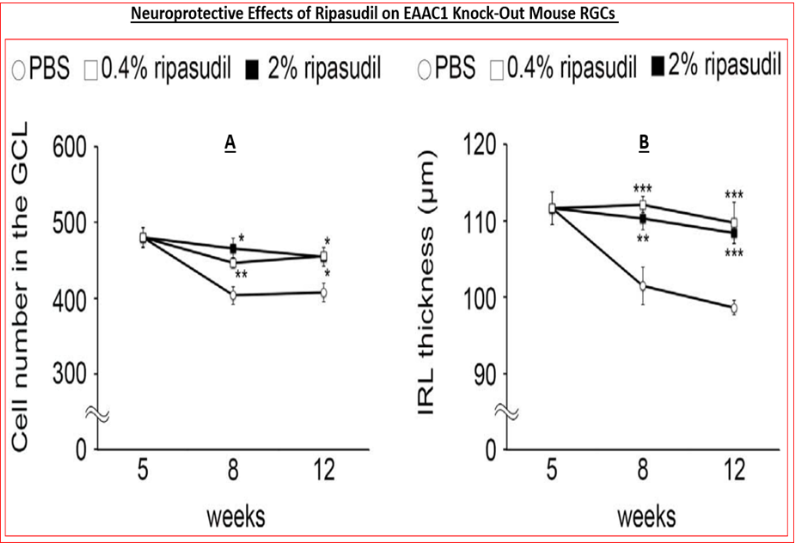
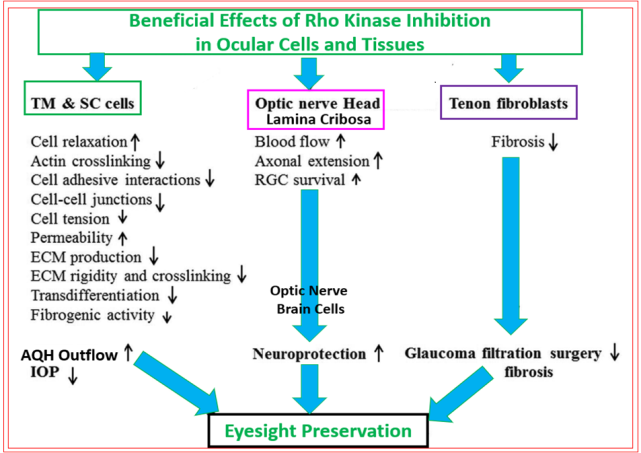
The other reasons that ROCK inhibitors represent important new drugs for ocular disease treatments involve their ability to reduce fibrosis induced by pro-fibrotic endogenous substances released by stressed TM cells such as TGF-β2 (Figure 9) [89, 114, 115], relax blood vessels to enhance local blood flow at the back of the eye [116, 117, 118], including the ONH, thereby overcoming ischemia/hypoxia that prevails in POAG/normotensive glaucoma [NTG] and in other forms of glaucoma [1, 2, 3, 4, 7, 71, 72]. Additionally, ROCK inhibitors exert neuroprotective activities by protecting (and preserving) RGCs from neuro-excitants like glutamate receptor agonists and other agents like endothelin that cause Ca2+-overloading of retinal neurons [119, 120, 121] (Figure 10). RGC axonal regeneration due to neurite extension by ROCK inhibitors such as Y-27632 and Y-39983 has also been reported [118, 121, 122, 123, 124, 125]. Consequently, blockers of the active sites of ROCKs are very useful addition to the clinicians’ armamentarium in the quest to find new generation of outflow-promoting drugs for OHT/glaucoma treatment for the preservation of eyesight that exert their beneficial effects via multiple mechanisms of action that are mediated via the actomyosin components [104, 105, 126] and potentially by dampening the microglial activity within the retina, optic nerve and the brain centers involved in transmission and decoding of visual signals [127] (Figure 11).
Some Selected Studies Characterizing Inhibitors of ROCKs
The human kinome is reported to contain over 500 different kinase enzymes grouped in families that catalyze phosphorylation of various lipids, carbohydrates and proteins, including other enzymes. Unsurprisingly, ROCKs are cornerstones of cellular signaling and communication, and their up-regulation is implicated in many diverse diseases/ disorders ranging from systemic hypertension, cardiovascular hypertrophy, diabetes mellitus, pulmonary hypertension, renal disease/vasospasm, asthma, cancer, erectile dysfunction, hyper-proliferative diseases, inflammatory diseases, and glaucoma [99, 106, 128, 129, 130, 131, 132, 133, 134, 135]. These findings prompted discovery and characterization of many classes of ROCK inhibitors (Figure 3) [86, 89, 106, 108, 116, 136, 137] and their eventual introduction into clinical practice for the medical treatments of the latter maladies.
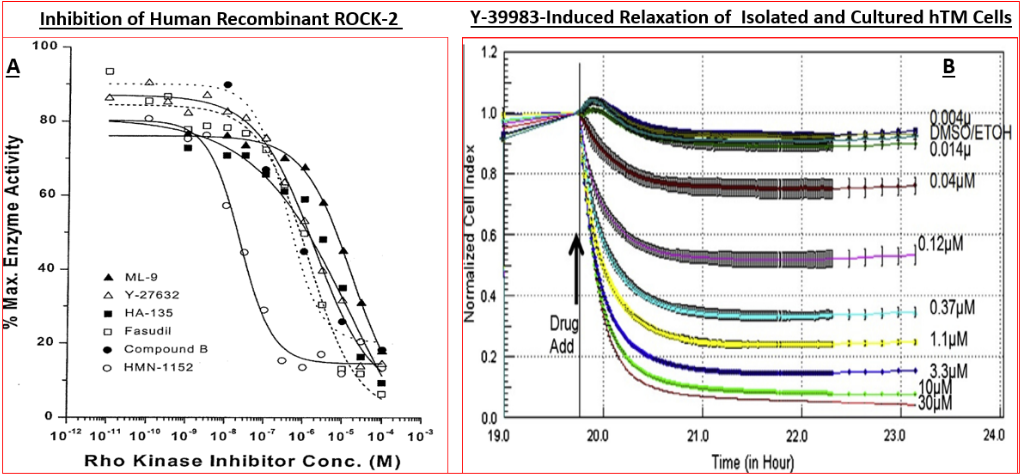
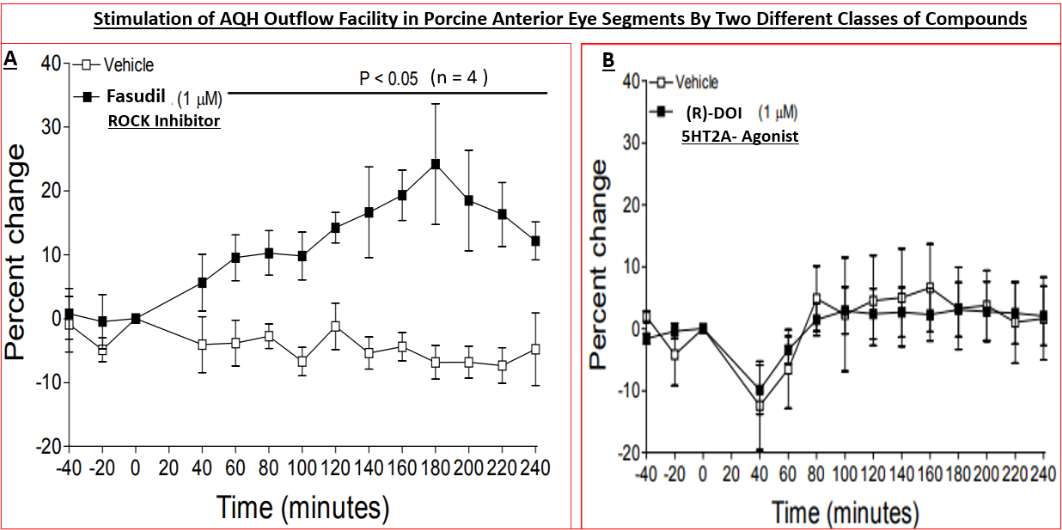
Specifically, nanomolar affinity/potency compounds that competitively compete for the active ATP-binding site of ROCK-1 and ROCK-2 have been reported that potently block these enzymes in vitro / ex vivo in a concentration- dependent manner (Figures 4A, 4B, 5 & 6) (Table 3) [89, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111, 112, 113, 114, 115, 116] and in vivo in animal models of disease to exert their biological effects such as potently and efficaciously lowering IOP and enhancing retinal blood flow in a dose-dependent manner (Figures.7, 8 & 10) [87, 89, 102, 103, 108, 109, 110, 111, 112, 113, 114, 115, 116, 117, 118]. All ROCK inhibitors traverse the cell membranes to engage with the cytoplasmic ROCKs and prevent down-stream signaling thereby relaxing cells/tissues, (e.g. isolated TM cells) (Figure 4B) [89, 102, 103, 104, 105], preventing cell migration/proliferation, reducing focal adhesions and phosphorylation of myosin light-chain kinase (Figure 5) [89, 102, 103, 104, 105], preventing pro- fibrotic actions (Figure 9) [89, 114, 115] and thus promoting fluid drainage from the ANC of the eye (Figure 6A) [137] where another class of compound failed to induce such an effect (Figurer 6B). In ocular physiology/pathology, ROCK- 2 seems more important that ROCK-1, but both isoforms of ROCK were elevated in the ONH of glaucoma patients [100]. Thus, most researchers have focused on inhibitors of ROCK-2, although due to the high homology between these isoforms, most such inhibitors also carry weaker ROCK-1 inhibitory activity as well [89, 102, 103], and most also show some degree of selectivity for ROCKs relative to other kinases (Table 4) [89].
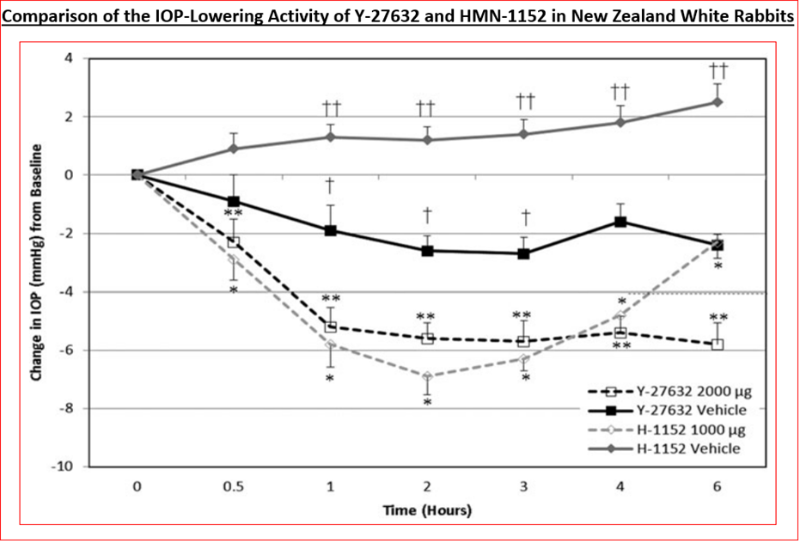
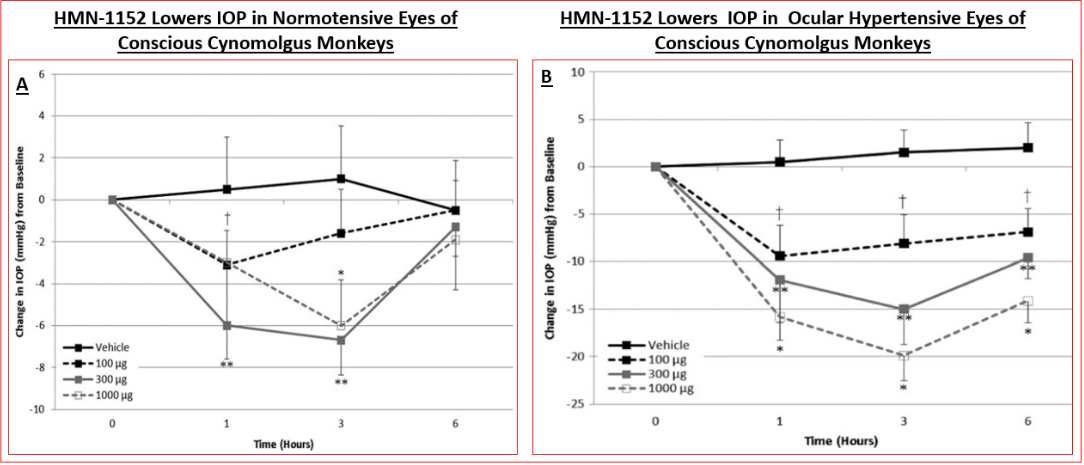
Functional activity of various ROCK inhibitors at recombinant human ROCK-2, using both radiometric (using [γ-33P]-ATP) and fluorescence polarization assays has been determined (Figure 4A) (Table 3) [89, 101, 136, 137]. Data from the literature shows that whilst the absolute inhibitory potency values for various ROCK inhibitors differ (Table 3), due to the use of human recombinant kinases from different sources and disparate cell-based assays (Figures 4B, 5 & 6), the relative rank order of potencies are fairly consistent among many laboratories [89, 101, 136, 137]. Despite such differences, the in vitro potencies are well translated in the IOP-lowering models of OHT/POAG in vivo whether ocular hypotensive activity is studied in rabbits or non- human primates [87, 89, 103, 107, 108]. Thus for instance, the higher affinity and potency of HMN-1152 relative to Y-27632 (Table 3) is borne out by its greater IOP-reducing activity after topical ocular dosing in both species (Figure 7) (Table 5). Indeed, the usefulness of the rabbit- and/ or monkey-based in vivo testing models of OHT/POAG have permitted optimization and selection of future drug candidates [102, 103]. Furthermore, the ability of at least ripasudil [85, 86, 87] and netarsudil [88, 106, 113, 138] to lower and control IOP in human subjects has been demonstrated in numerous clinical trials, and the mechanism of action to reduce IOP confirmed to be by enhancing AQH drainage via the conventional TM/SC outflow pathway [139]. Due to this robust outflow promoting effect of netarsudil and ripasudil, fixed-dose combination products of each drug [112], with additive ocular hypotensive actions, have been also approved by many health authorities of several different countries. Whether the anti-fibrotic, neuroprotective and axonal regenerating efficacies of these ROCK inhibitors observed in vitro and in animal models noted above can be translated to the human RGC, RGC axonal and thalamic nuclei/visual cortical degenerative conditions characteristic of POAG/NTG patients remains to be determined [140].
In Vitro Potency OF Rho-Associated Protein Kinase Inhibitors (nM) Compound ROCK1 Ki ROCK2 Ki PKA Ki PKCT Ki MRCKA Ki CAM2A Ki PTM IC50 HTM IC50 Netarsudil 1 1 5 92 129 5,3 12 79 16 Netarsudil-MI 0.2 0.2 1 27 7 13,689 2 3 AR-12286 2 2 69 9,322 28 5,855 924 818 Y27632 22 41 21,006 413 485 16,863 9,970 1,738 Fasudil 76 47 216 3,162 5,983 3,162 10,060 3,942 Values represent the average of ≥3 replicate experiments.CAM2A, calcium/calmodulin-dependent protei n kinase 2A; HTM, human trabecular meshwork ; IC50, half maximal inhibitory concentration; K1, inhibition constant; MRCKA, myotonic dystrophy kinase-related CDC42-binding kinase A; PKA, protein kinase A; PKCT, protei n kinase C, theta; PTM, porcine trabecular meshwork; ROCK1, Rho-associated protei n kinase I ; ROCK2, Rho-associated protein kinase 2. Table 4: The relative potency and selectivities of various ROCK inhibitors determined in vitro. Note that potency is inversely related to IC50 or K1 values.
- IOP Reduction By Various ROCK Inhibitors in OHT Monkey Eyes
- %Max. IOP Reduction in Ocular Hypertensive Monkey Eyes
- Compound
- Topical Ocular Dose
- (µg)
- 1 Hour Post
- Dosing
- 3 Hour Post
- Dosing
- 6 Hour Post
- Dosing
- 100 µg
- 28
- 25
- 19
- HMN-1152 (ROCK II IC50=47 nM)
- 300 µg
- 29
- 30
- 21
- 1mg
- 42
- 51
- 36
- Fasudil (ROCK II IC50=1690 n M)
- 500 µg
- 33
- 28
- 16
- H-7 (ROCK II IC50=2341 n M)
- 1 mg
- 31
- 21
- 12
- Y-27632 (ROCK II IC50=2802 n M)
- 300 µg
- 15
- 14
- 8 l mg
- 24
- 36
- 32
- HA-135 (ROCK II IC50= 6702 n M)
- 500 µg
- 19
- 15
- 1
- Cmpd D (ROCK II IC50=2625 nM)
- 500 µg
- 23
- 20
- 20
- ML-9 (ROCK II IC50 = 12,000 nM)
- 300 µg
- 7
- 6
- 5
Table 6: Relative efficacies of ROCK inhibitors in lowering IOP.
Conclusions
This mini-review has endeavored to present current knowledge of the role of ROCKs in various pathological conditions, and the utility of inhibitors of ROCK-1 and ROCK-2 to ameliorate such human ocular diseases as ocular hypertension, POAG and NTG. Due to the multiple beneficial actions of ROCK inhibitors in ocular physiology ranging from the front of the eye to the retina and beyond (Figure 11), they represent uniquely valuable new drugs in the tool chest of ophthalmic clinicians to help them treat the ever- increasing number of global patients with chronic OHT and various forms of glaucoma. The TM/SC cell, RGC and RGC axon protecting actions of ROCK inhibitors observed in vitro and in animals, coupled with their ocular hypotensive activity demonstrated in animals and humans, and enhancement of retinal blood flow in several species clearly differentiates them from other classes of drugs used to treat OHT and glaucoma. However, whether the neuroprotective, neurite and axonal elongation and blood-flow enhancing actions of ROCK inhibitors can unequivocally be shown in a reproducible manner in multiple clinical trials remains to be investigated. The exaggerated hyperemic activity associated with most if not all ROCK inhibitors may limit their topical ocular utility but it is too early to predict that aspect since they only have a recent approval history and much more data need to be generated and shared. We hope that suitable formulations and other forms of drug delivery, alone or in combination with other agents, can be discovered and deployed to reduce the ocular surface redness that ROCK inhibitors cause without impacting their beneficial actions in terms of helping to preserve vision on a long-term basis.
References
-
Weinreb RN, Aung T, Medeiros FA (2014) The pathophysiology and treatment of glaucoma: a review. JAMA 311(18): 1901-1911.
-
Jonas JB, Aung T, Bourne RR, Ritch R, Jonas SP (2017) Glaucoma. Lancet 390(10108): 2183-2193.
-
Sharif NA (2017) Ocular hypertension and glaucoma: a review and current perspectives. Int J Ophthalmol Vis Sci 2(2): 22-36.
-
Sharif NA (2021) Therapeutic drugs and devices for tackling ocular hypertension and glaucoma, and need for neuroprotection and cytoprotective therapies. Front Pharmacol 12: 729249.
-
Harwerth RS, Quigley HA (2006) Visual field defects and retinal ganglion cell losses in patients with glaucoma. Arch Ophthalmol 124(6): 853-859.
-
Johannesson G, Eklund A, Linden C (2018) Intracranial and intraocular pressure at the lamina cribrosa: gradient effects. Curr Neurol Neurosci Rep 18(5): 25.
-
Kim JH, Caprioli J (2018) Intraocular pressure fluctuation: is it important? J Ophthalmic Vis Res 13(12): 170-174.
-
Pasquale LR (2016) Vascular and autonomic dysregulation in primary open-angle glaucoma. Curr Opin Ophthalmol 27(2): 94-101.
-
Pease ME, McKinnon SJ, Quigley HA, Baumrind LAK, Zack DJ (2000) Obstructed axonal transport of BDNF and its receptor TrkB in experimental glaucoma. Invest Ophthalmol Vis Sci 41(3): 764-774.
-
Burgoyne CF, Downs JC, Bellezza AJ, Suh JKF, Hart RT (2005) The optic nerve head as a biomechanical structure; a new paradigm for understanding the role of IOP-related stress and strain in the pathophysiology of glaucomatous optic nerve head damage. Prog Retinal Eye Res. 24(1): 39-73.
-
Downs JC, Roberts MD, Sigal IA (2011) Glaucomatous cupping of the lamina cribrosa: a review of the evidence for active progressive remodeling as a mechanism. Exp Eye Res 93(2): 133-140.
-
Quigley HA, Dunklberger GR, Green WR (1989) Retinal ganglion cell atrophy correlated with automated perimetry in human eyes with glaucoma. Am J Ophthalmol 107(5): 453-464.
-
Nuschke AC, Farrell SR, Levesque JM, Chauhan BC (2015) Assessment of retinal ganglion cell damage in glaucomatous optic neuropathy: axon transport, injury and soma loss. Exp Eye Res 141:111-124.
-
Yucel YH, Zhang Q, Weinreb RN, Kaufman PL, Gupta N (2001) Atrophy of relay neurons in magno- and parvocellular layers in the lateral geniculate nucleus in experimental glaucoma. Invest Ophthalmol Vis Sci 42(13): 3216-3222.
-
Gupta N, Ly T, Zhang Q, Kaufman PL, Weinreb RN, et al. (2007) Chronic ocular hypertension induces dendrite pathology in the lateral geniculate nucleus of the brain. Exp Eye Res 84(1): 176-184.
-
Yu L, Xie L, Dai C, Xie B, Liang M, et al. (2015) Progressive thinning of visual cortex in primary open-angle glaucoma of varying severity. PLoS One 10(3): e0121960.
-
Trivedi V, Bang JW, Parra C, Colbert MK, O’Connell C, et al. (2019) Widespread brain reorganization perturbs visuomotor coordination in early glaucoma. Sci Rep 9(1): 14168.
-
Calkins DJ, Horner PJ (2012) The cell and molecular biology of glaucoma: axonopathy and the brain. Invest Ophthalmol Vis Sci 53(5): 2482-2484.
-
Crabb DP (2016) A view on glaucoma- are we seeing it clearly?. Eye 30: 304-313.
-
Tham YC, Li X, Wong TY, Quigley HA, Aung T, et al. (2014) Global prevalence of glaucoma and projections of glaucoma burden through 2040. Ophthalmol 121(11): 2081-2090.
-
Sharif NA (2018) iDrugs and iDevices discovery and development - preclinical assays, techniques and animal model studies for ocular hypotensives and neuroprotectants. J Ocular Pharmacol Ther 34(1): 7-39.
-
Sharif NA (2018) Glaucomatous optic neuropathy treatment options: the promise of novel therapeutics, techniques and tools to help preserve vision. Neural Regen Res 13(7): 1145-1150.
-
Howell GR, MacNicoll KH, Braine CE, Soto I, Macalinao DG, et al. (2014) Combinatorial targeting of early pathways profoundly inhibits neurodegeneration in a mouse model of glaucoma. Neurobiol Dis 71: 44-52.
-
Civan M, Macknight AD (2004) The ins and outs of aqueous humor secretion. Exp Eye Res 78(3): 625-631.
-
Carreon T, van der Merwe E, Fellman RL, et al. (2017) Aqueous outflow- a continuum from trabecular meshwork to episcleral veins. Prog Retin Eye Res 57: 108-133.
-
Buffault J, Labbe A, Hamard P, Baudouin FB, Baudouin C (2020) The trabecular meshwork: Structure, function and clinical implications. A review of the literature. J Fr Ophtalmol 43(7): e217-e230.
-
Acott TS, Vranka JA, Keller KE, Raghunathan V, Kelley MJ (2020) Normal and glaucomatous outflow regulation. Prog Retin Eye Res 82: 100897.
-
Stamer WD, Braakman ST, Zhou EH, Ether CR, Fredberg JJ, et al. (2015) Biomechanics of Schlemm’s canal endothelium and intraocular pressure reduction. Prog Retin Eye Res 44: 86-98.
-
Johnson DH (1996) Human trabecular meshwork cell survival is dependent on perfusion rate. Invest Ophthalmol Vis Sci 37(6): 1204-1208.
-
Babizhayev MA, Yegorov YE (2011) Senescent phenotype of trabecular meshwork cells displays biomarkers in primary open-angle glaucoma. Curr Mol Med 11(7): 528-552.
-
Patel G, Fury W, Yang H, Caraballo MG, Bai Y, et al. (2020) Molecular taxonomy of human ocular outflow tissues defined by single-cell transcriptomics. Proc Natl Acad Sci USA 117(23): 12856-12867.
-
Wang LY, Su GY, Wei ZY, Zhang ZJ, Liang QF (2020) Progress in the basic and clinical research on the Schlemm’s canal. Int J Ophthalmol 13(5): 816-821.
-
Von Zee CL, Langert KA, Stubbs EB (2012) Transforming growth factor-β2 induces synthesis and secretion of endothelin-1 in human trabecular meshwork cells. Invest Ophthalmol Vis Sci 53(9): 5279-5286.
-
Nakamura N, Yamagishi R, Honjo M, Igarashi N, Shimizu S, et al. (2021) Effects of topical TGF-β1, TGF-β2, ATX, and LPA on IOP elevation and regulation of the conventional aqueous humor outflow pathway. Mol Vis 27: 61-77.
-
De Groef L, Andries L, Siwakoti A, Bollaerts I, Noterdaeme L, et al. (2016) Aberrant collagen composition of the trabecular meshwork results in reduced aqueous humor drainage and elevated IOP in MMP-9 Null Mice. Invest Ophthalmol Vis Sci 57(14): 5984-5995.
-
Kasetti RB, Maddineni P, Millar JC, Clark AF, Zode GS,et al. (2017) Increased synthesis and deposition of extracellular matrix proteins leads to endoplasmic reticulum stress in the trabecular meshwork. Sci Rep 7(1): 14951.
-
Yemanyi F, Vranka J, Raghunathan VK (2020) Crosslinked extracellular matrix stiffens human trabecular meshwork cells via dysregulating β-catenin and YAP/TAZ signaling pathways. Invest Ophthalmol Vis Sci 61(10): 41.
-
Alvarado J, Murphy C, Juster R (1984) Trabecular meshwork cellularity in primary open-angle glaucoma and nonglaucomatous normals. Ophthalmology 91(6): 564-579.
-
Eiki O, Yukihiro M, Kiyomi A (2010) Free radicals in the aqueous humor of patients with glaucoma. Clin Ophthalmol 4: 653-660.
-
Sacca SC, Pascotto A, Camicione P, Capris P, Izzotti A (2005) Oxidative DNA damage in the human trabecular meshwork: clinical correlation in patients with primary open-angle glaucoma. Arch Ophthalmol 123(4): 458- 463.
-
Izzotti A, Bagnis A, Sacca SC (2006) The role of oxidative stress in glaucoma. Mutat Res 612(2): 105-114.
-
Izzotti A, Longobardi M, Cartiglia C, Sacca SC (2011) Mitochondrial damage in the trabecular meshwork occurs only in primary open-angle glaucoma and in pseudoexfoliative glaucoma. PLoS One 6(1): e14567.
-
He Y, Ge J, Tink TJ (2008) Mitochondrial defects and dysfunction in calcium regulation in glaucomatous trabecular meshwork cells. Invest Ophthalmol Vis Sci 49(11): 4912-4922.
-
Porter K, Hirt J, Stamer WD, Liton PB (2015) Autophagic dysregulation in glaucomatous trabecular meshwork cells. Biochim Biophys Acta 1852(3): 379-385.
-
Xu G, Weinreb RN, Leung CK (2014) Optic nerve head deformation in glaucoma: the temporal relationship between optic nerve head surface depression and retinal nerve fiber layer thinning. Ophthalmol 121(2): 2362- 2370.
-
Neufeld AH, Hernandez MR, Gonzalez M (1997) Nitric oxide synthase in the human glaucomatous optic nerve head. Arch Ophthalmol 115(4): 497-503.
-
Neufeld AH (1999) Microglia in the optic nerve head and the region of parapapillary chorioretinal atrophy in glaucoma. Arch Ophthalmol 117(8): 1050-1056.
-
Hernandez MR, Luo XX, Andrzejewska W, Neufeld AH (1989) Age-related changes in the extracellular matrix of the human optic nerve head. Am J Ophthalmol 107(5): 476-484.
-
McElnea EM, Quill B, Docherty NG, Irnaten M, Siah WF, et al. (2011) Oxidative stress, mitochondrial dysfunction and calcium overload in human lamina cribrosa cells from glaucoma donors. Mol Vis 17: 1182-1189.
-
McElnea EM, Hughes E, McGoldrick A, McCann A, Quill B, et al. (2014) Lipofuscin accumulation and autophagy in glaucomatous human lamina cribrosa cells. BMC Ophthalmol 14:153.
-
Wilson GN, Inman DM, Crish DCM, Smith MA, Crish SD (2015) Early pro-inflammatory cytokine elevations in the DBA/2J mouse model of glaucoma. J Neuroinflamm 12: 176.
-
Adornetto A, Russo R, Parisi V (2019) Neuroinflammation as a target for glaucoma therapy. Neural Regen Res 14(3): 391-394.
-
Evangelho K, Mogilevskaya M, Barragan LM, Sanchez VJK (2019) Pathophysiology of primary open-angle glaucoma from a neuroinflammatory and neurotoxicity perspective: a review of the literature. Int Ophthalmol 39(1): 259-271.
-
Hollander H, Makarov F, Stefani FH, Stone J (1995) Evidence of constriction of optic axons at the lamina cribrosa in the normotensive eye in humans and other mammals. Ophthalmic Res 27(5): 296-309.
-
Howell GR, Libby RT, Jakobs TC, Smith RS, Phalan FC, et al. (2007) Axons of retinal ganglion cells are insulted in the optic nerve early in DBA/2J glaucoma. J Cell Biol 179(7): 1523-1537.
-
Stokely ME, Brady ST, Yorio T (2002) Effects of endothelin-1 on components of anterograde axonal transport in optic nerve. Invest Ophthalmol Vis Sci 43(10): 3223-3230.
-
Chintala SK (2006) The emerging role of proteases in retinal ganglion cell death. Exp Eye Res 82(1): 5-12.
-
Chaphalkar RM, Stankowska DL, He S, Kodati B, Philips N, et al. (2020) Endothelin-1 mediated decrease in mitochondrial gene expression and bioenergetics contribute to neurodegeneration of retinal ganglion cells. Sci Rep 10(1): 3571.
-
Bosco A, Steele MR, Vetter ML (2011) Early microglia activation in a mouse model of chronic glaucoma. J Comp Neurol 519(4): 599-620.
-
Rashid K, Akhtar-Schaefer I, Langmann T (2019) Microglia in retinal degeneration. Front Immunol 10: 1975.
-
Silverman SM, Kim BJ, Howell GR, Miller J, John SWM, et al. (2016) C1q propagates microglial activation and neurodegeneration in the visual axis following retinal ischemia/reperfusion injury. Mol Neurodegener 11: 24.
-
Lam D, Jim J, To E, Rasmussen C, Kaufman PL, et al. (2009) Astrocyte and microglial activation in the lateral geniculate nucleus and visual cortex of glaucomatous and optic nerve transected primates. Mol Vis 15: 2217- 2229.
-
Maddineni P, Kasetti RB, Patel PD, Millar JC, Kiehlbauch C, et al. (2020) CNS axonal degeneration and transport deficits at the optic nerve head precede structural and functional loss of retinal ganglion cells in a mouse model of glaucoma. Mol Neurodegener 15(1): 48.
-
Osborne NN, Alvarez CN, Joglar B, Aguado SDO (2016) Glaucoma: focus on mitochondria in relation to pathogenesis and neuroprotection. Eur J Pharmacol 787: 127-133.
-
Ou Y, Jo RE, Ullian EM, Wong ROL, Santina LD (2016) Selective vulnerability of specific retinal ganglion cell types and synapses after transient ocular hypertension. J Neurosci 36(35): 9240-9252.
-
Park HY, Lee KI, Lee K, Shin HY, Park CK (2014) Torsion of the optic nerve head is a prominent feature of normal- tension glaucoma. Invest Ophthalmol Vis Sci 56(1): 156- 163.
-
Zhang HJ, Mi XS, So KF (2019) Normal tension glaucoma: from the brain to the eye or the inverse?. Neural Regen Res 14(11): 1845-1850.
-
(CNTGSG) Collaborative Normal-Tension Glaucoma Study Group (1998) The effectiveness of intraocular pressure reduction in the treatment of normal-tension glaucoma. Am J Ophthalmol 126(4): 498-505.
-
AGIS Investigators (2000) The Advanced Glaucoma Intervention Study (AGIS): 7. The relationship between control of intraocular pressure and visual field deterioration. Am J Ophthalmol 130(4): 429-440.
-
Heijl A, Leske MC, Bengtsson B, Hyman L, Bengtson B, et al. (2002) Reduction of intraocular pressure and glaucoma progression: results from the Early Manifest Glaucoma Trial. Arch Ophthalmol 120(10): 1268-1279.
-
Kass MA, Heuer DK, Higginbotham EJ, Johnson CA, Keltner JL, et al. (2002) The Ocular Hypertension Treatment Study: a randomized trial determines that topical ocular hypotensive medication delays or prevents the onset of primary open-angle glaucoma. Arch Ophthalmol 120(6): 701-713.
-
Musch DC, Gillespie BW, Niziol LM, Lichter PR, Varma R, et al. (2011) Intraocular pressure control and long-term visual field loss in the Collaborative Initial Glaucoma Treatment Study. Ophthalmology 118 (9): 1766-1773.
-
Lee EJ, Kim TW, Weinreb RN, Kim H (2013) Reversal of lamina cribrosa displacement after intraocular pressure reduction in open-angle glaucoma. Ophthalmology 120(3): 553-559.
-
Bhandari A, Crabb DP, Poinoosawmy D, Fitzke FW, Hitchings RA, et al. (1997) Effect of surgery on visual field progression in normal-tension glaucoma. Ophthalmology 104(7): 1131-1137.
-
Sehi M, Grewal DS, Goodkin ML, Greenfield DS (2010) Reversal of retinal ganglion cell dysfunction after surgical reduction of intraocular pressure. Ophthalmology 117(12): 2329-2336.
-
Tu S, Li K, Ding X, Hu D, Li K, et al. (2019) Relationship between intraocular pressure and retinal nerve fibre thickness loss in a monkey model of chronic ocular hypertension. Eye (Lond) 33(12): 1833-1841.
-
Toris CB, Gabelt BAT, Kaufman PL (2008) Update on the mechanism of action of topical prostaglandins for intraocular pressure reduction. Surv Ophthalmology 53 (S1): S107-S120.
-
Hellberg MR, McLaughlin MA, Sharif NA, DeSantis L, Dean TR, et al. (2002) Identification and characterization of the ocular hypotensive efficacy of Travoprost, a potent and selective FP prostaglandin receptor agonist, and AL-6598, a DP prostaglandin receptor agonist. Surv Ophthalmol 47(S1): S13-S33.
-
Kirihara T, Taniguchi T, Yamamura K, Iwamura R, Yoneda K, et al. (2018) Pharmacologic characterization of omidenepag isopropyl, a novel selective EP2 receptor agonist, as an ocular hypotensive agent. Invest Ophthalmol Vis Sci 59(1): 145-153.
-
Fuwa M, Toris CB, Fan S, Taniguchi T, Ichikawa M, et al. (2018) Effects of a novel selective EP2 receptor agonist, omidenepag isopropyl, on aqueous humor dynamics in laser-induced ocular hypertensive monkeys. J Ocul Pharmacol Ther 34(7): 531-537.
-
Aihara M, Lu F, Kawata H, Iwatabc A, Kawabata NO, et al. (2020) Omidenepag Isopropyl versus latanoprost in primary open-angle glaucoma and ocular hypertension: The Phase 3 AYAME Study. Am J Ophthalmol 220: 53-63.
-
Cavet ME, DeCory HH (2018) The role of nitric oxide in the intraocular pressure lowering efficacy of latanoprostene bunod: review of nonclinical studies. J Ocul Pharmacol Ther 34(1-2): 52-60.
-
Hollo G, Topouzis F, Fechtner RD (2014) Fixed- combination intraocular pressure-lowering therapy for glaucoma and ocular hypertension: advantages in clinical practice. Expert Opin Pharmacother 15(12): 1737-1747.
-
Asrani S, Bacharach J, Holland E, McKee H, Sheng H, et al. (2020) Fixed-dose combination of netarsudil and latanoprost in ocular hypertension and open-angle glaucoma: pooled efficacy/safety analysis of phase 3 MERCURY-1 and -2. Adv Ther 37(4): 1620-1631.
-
Tanihara H, Inoue T, Yamamoto T, Kuwayama Y, Abe H, et al. (2013) Phase-2 randomized clinical study of a rho kinase inhibitor, K-115, in primary open-angle glaucoma and ocular hypertension. Am J Ophthalmol 156(4): 731- 776.
-
Garnock-Jones KP (2014) Ripasudil: first global approval. Drugs 74(18): 2211-2215.
-
Inoue T, Tanihara H (2017) Ripasudil hydrochloride hydrate: targeting rho kinase in the treatment of glaucoma. Expert Opin Pharmacother 18(15): 1669- 1673.
-
Kopczynski CC, Epstein DL (2014) Emerging trabecular outflow drugs. J Ocul Pharmacol Ther 30(2-3): 85-87.
-
Lin CW, Sherman B, Moore LA, Laethem CL, Da-Wen Lu, et al. (2018) Discovery and preclinical development of netarsudil, a novel ocular hypotensive agent for the treatment of glaucoma. J Ocul Pharmacol Ther 34(1-2): 40-51.
-
Savinainen A, Prusakiewicz JJ, Oswald J, Spencer E, Lou Z, et al. (2019) Pharmacokinetics and intraocular pressure- lowering activity of TAK-639, a novel C-type natriuretic peptide analog, in rabbit, dog, and monkey. Exp Eye Res 189: 107836.
-
Roy Chowdhury U, Dosa PI, Fautsch MP (2017) ATP sensitive potassium channel openers: A new class of ocular hypotensive agents. Exp Eye Res 158: 85-93.
-
Li G, Nottebaum AF, Brigell M, Navarro ID, Ipe U, et al. (2020) A small molecule inhibitor of VE-PTP activates Tie2 in Schlemm’s canal increasing outflow facility and reducing intraocular pressure. Invest Ophthalmol Vis Sci 61(14): 12.
-
Geffen N, Assia EI, Melamed S (2017) Laser-assisted techniques for penetrating and nonpenetrating glaucoma surgery. Dev Ophthalmol 59: 100-112.
-
Garg A, Gazzard G (2018) Selective laser trabeculoplasty: past, present, and future. Eye (Lond) 32(5): 863-876.
-
Pillunat LE, Erb C, Jünemann AG, Kimmich F (2017) Micro-invasive glaucoma surgery (MIGS): a review of surgical procedures using stents. Clin Ophthalmol 11: 1583-1600.
-
Sadruddin O, Pinchuk L, Angeles R, Palmberg P (2019) Ab externo implantation of the MicroShunt, a poly (styrene- block-isobutylene-blockstyrene) surgical device for the treatment of primary open-angle glaucoma: a review. Eye Vision (Lond) 6: 36.
-
Guymer C, Wood JP, Chidlow G, Casson RJ (2019) Neuroprotection in glaucoma: recent advances and clinical translation. Clin Exp Ophthalmol 47(1): 88-105.
-
Boia R, Ruzafa N, Aires ID, Pereiro X, Ambrósio AF, et al. (2020) Neuroprotective strategies for retinal ganglion cell degeneration: current status and challenges ahead. Int J Mol Sci 21(7): 2262.
-
Rao PV, Pattabiraman PP, Kopczynski C (2017) Role of the rho GTPase/rho kinase signaling pathway in pathogenesis and treatment of glaucoma: bench to bedside research. Exp Eye Res 158: 23-32.
-
Goldhagen B, Proia AD, Epstein D L, Rao PV (2012) Elevated levels of rho A in the optic nerve head of human eyes with glaucoma. J Glaucoma 21(8): 530-538.
-
Sharif NA, Drace C, Williams G (2008) High- throughput assay for human rho kinase activity with enhanced signal-to-noise ratio. US Patent Application 20080096238.
-
Henderson AJ, Hadden M, Guo C, Douglas N, Decornez H, et al. (2010) 2,3-Diaminopyrazines as rho kinase inhibitors. Bioorg Med Chem Lett 20(3): 1137- 1140.
-
Chen HH, Namil A, Severns B, Ward J, Kelly C, et al. (2014) In vivo optimization of 2,3-diaminopyrazine rho kinase inhibitors for the treatment of glaucoma. Bioorg Med Chem Lett 24(8): 1875-1879.
-
Ramachandran C, Patil RV, Combrink K, Sharif NA, Srinivas SP, et al. (2011) Rho-Rho kinase pathway in the actomyosin contraction and cell-matrix adhesion in immortalized human trabecular meshwork cells. Mol Vis 17: 1877-1890.
-
Ramachandran C, Patil RV, Sharif NA, Srinivas SP (2011) Effect of elevated intracellular cAMP on actomyosin contraction in bovine trabecular meshwork cells. Invest Ophthalmol Vis Sci 52(3): 1474-1485.
-
Vohra V, Chawla H, Gupta M (2014) ROCK inhibitors: future of anti-glaucoma medication. Ophthalmol Res An Int J 2(60): 361-367.
-
Nakajima E, Nakajima T, Minagawa Y, Shearer TR, Azuma M, et al. (2005) Contribution of ROCK in contraction of trabecular meshwork: proposed mechanism for regulating aqueous outflow in monkey and human eyes. J Pharm Sci 94(4): 701-708.
-
Rao PV, Deng PF, Kumar J, Epstein DL (2001) Modulation of aqueous humor outflow facility by the rho kinase-specific inhibitor Y-27632. Invest Ophthalmol Vis Sci 42(5):1029-1037.
-
Honjo M, Tanihara H (2018) Impact of the Clinical use of ROCK inhibitor on the pathogenesis and treatment of glaucoma. Jpn J Ophthalmol 62(2): 109-126
-
Tokushige H, Inatani M, Nemoto S, Sakaki H, Katayama K, et al. (2007) Effects of topical administration of Y-39983, a selective rho-associated protein kinase inhibitor, on ocular tissues in rabbits and monkeys. Invest Ophthalmol Vis Sci 48(7): 3216-3222.
-
Yu M, Chen X, Wang N, Cai S, Qiu J, et al. (2008) H-1152 Effects on intraocular pressure and trabecular meshwork morphology of rat eyes. J Ocul Pharmmacol Ther 24(4): 373-379.
-
Leary KA, Steibel JP, Harman CD, Anderson AL, Komaromy AM, at al. (2021) Safety and efficacy of topically administered netarsudil-latanoprost fixed dose combination (FDC; Rocklatan™) in normal and glaucomatous dogs with ADAMTS10-open-angle glaucoma (ADAMTS10-OAG). Vet Ophthalmol 24(6): 610-619.
-
Asrani S, Bacharach J, Holland E, McKee H, Sheng H, et al. (2020) Fixed-dose combination of netarsudil and latanoprost in ocular hypertension and open-angle glaucoma: pooled efficacy/safety analysis of phase 3 MERCURY-1 and -2. Adv Ther 37(4): 1620-1631.
-
Pattabiraman PP, Rinkoski T, Poeschla E, Proia A (2015) RhoA GTPase-induced ocular hypertension in a rodent model is associated with increased fibrogenic activity in the trabecular meshwork. Am J Pathol 185(2): 496-512.
-
Li G, Lee C, Read AT, Wang K, Ha J, et al. (2021) Anti-fibrotic activity of a rho-kinase inhibitor restores outflow function and intraocular pressure homeostasis. Elife 10: e60831.
-
Watabe H, Abe S, Yoshitom T (2011) Effects of Rho-Associated Protein Kinase Inhibitors Y-27632 and Y-39983 on Isolated Rabbit Ciliary Arteries. Jpn J Ophthalmol 55(4): 411-417.
-
Sugiyama T, Shibata M, Kajiura S, Okuno T, Tonari M, et al. (2011) Effects of fasudil, a rho-associated protein kinase inhibitor, on optic nerve head blood flow in rabbits. Invest Ophthalmol Vis Sci 52: 64-69.
-
Tokushige H, Waki M, Takayama Y, Tanihara H (2011) Effects of Y-39983, a selective rho-associated protein kinase inhibitor, on blood flow in optic nerve head in rabbits and axonal regeneration of retinal ganglion cells in rats. Curr Eye Res 36(10): 964-970.
-
Yamashita K, Kotani Y, Nakajima Y, Shimazawa M, Yoshimura S, et al. (2007) Fasudil, a rho kinase (ROCK) inhibitor, protects against ischemic neuronal damage in vitro and in vivo by acting directly on neurons. Brain Res 1154: 215-224.
-
Yamamoto K, Maruyama K, Himori N, Omodaka K, Yokoyama Y, et al. (2014) The novel rho kinase (ROCK) inhibitor K-115: a new candidate drug for neuroprotective treatment in glaucoma. Invest Ophthalmol Vis Sci 55(11): 7126-7136.
-
Shaw PX, Sang A, Wang Y, Ho D, Douglas C, et al. (2017) Topical administration of a ROCK/NET inhibitor promotes retinal ganglion cell survival and axon regeneration after optic nerve injury. Exp Eye Res 158: 33-42.
-
Bertrand J, Winton MJ, Rodriguez-Hernandez N, Campenot RB, McKerracher L (2005) Application of Rho antagonist to neuronal cell bodies promotes neurite growth in compartmented cultures and regeneration of retinal ganglion cell axons in the optic nerve of adult rats. J Neurosci 25(5): 1113-1121.
-
Sagawa H, Terasak H, Nakamura M, Ichikawa M, Yata T, et al. (2007) A novel rock inhibitor, Y-39983, promotes regeneration of crushed axons of retinal ganglion cells into the optic nerve of adult cats. Exp Neurol 205(1): 230-240.
-
Ichikawaa M, Yoshidaa J, Saitoa K, Sagawab H, Tokita Y, et al. (2008) Differential effects of two ROCK inhibitors, fasudil and Y-27632 on optic nerve regeneration in adult cats. Brain Res 1201: 23-33.
-
Yang Z, Wang J, Liu X, Cheng Y, Deng L, Zhong Y (2013) Y-39983 downregulates rho A/rho-associated kinase expression during its promotion of axonal regeneration. Oncol Rep 29(3): 1140-1146.
-
Keller KE, Kopczynski C (2020) Effects of netarsudil on actin-driven cellular functions in normal and glaucomatous trabecular meshwork cells: a live imaging study. J Clin Med 9(11): 3524.
-
Roser AE, Tonges L, Lingor P (2017) Modulation of microglial activity by rho kinase (ROCK) inhibition as therapeutic strategy in Parkinson’s disease and amyotrophic lateral sclerosis. Front Aging Neurosci 9: 94.
-
Koch JC, Tatenhorst L, Roser AE, Saal KA (2018) ROCK inhibition in models of neurodegeneration and its potential for clinical translation. Pharmacol Ther 189: 1-21.
-
Sharma P, Roy K (2020) ROCK-2-selective targeting and its therapeutic outcomes. Drug Discov Today 25(2): 446-455.
-
Martín-Cámara O, Cores Á, López-Alvarado P, Menéndez JC (2021) Emerging targets in drug discovery against neurodegenerative diseases: Control of synapsis disfunction by the RhoA/ROCK pathway. Eur J Med Chem 225: 113742.
-
Saha BC, Kumari R, Kushumesh R, Ambasta A, Sinha BP (2021) Status of rho kinase inhibitors in glaucoma therapeutics-an overview. Int Ophthalmol.
-
Mendoza FA, Jimenez SA (2021) Serine-threonine kinase inhibition as antifibrotic therapy: TGF-β and ROCK inhibitors. Rheumatology (Oxford).
-
Singh A, Behl T, Sehgal A, Singh S, Sharma N, et al. (2021) Exploring the therapeutic promise of targeting Rhokinase in rheumatoid arthritis. Inflammopharmacol 29(6): 1641-1651.
-
Kimura T, Horikoshi Y, Kuriyagawa C, Niiyama Y (2021) Rho/ROCK Pathway and noncoding RNAs: implications in ischemic stroke and spinal cord injury. Int J Mol Sci 22(21): 11573.
-
Kim S, Kim SA, Han J, Kim IS (2021) Rho-Kinase as a Target for Cancer therapy and its immunotherapeutic potential. Int J Mol Sci 22(23): 12916.
-
Velde SV, Bergen TV, Sijnave D, Hollanders K, Defert O, et al. (2014) AMA0076, a novel, locally acting rho kinase inhibitor, potentially lowers intraocular pressure in New Zealand white rabbits with minimal hyperemia. Invest Ophthalmol Vis Sci 55(2): 1006-1016.
-
Ashwinbalaji S, Haribalaganesh R, Krishnadas S, Muthukkaruppan V, Senthilkumari S (2020) SB772077B (SB77) alleviated the aqueous outflow resistance mediated by cyclic mechanical stress in perfused human cadaveric eyes. Sci Rep 10(1): 10202.
-
Serle JB, Katz LJ, McLaurin E, Heah T, Ramirez‐Davis N, et al. (2018) Two Phase 3 clinical trials comparing the safety and efficacy of netarsudil to timolol in patients with elevated intraocular pressure: rho kinase elevated IOP treatment trial 1 and 2 (ROCKET-1 and ROCKET-2). Am J Ophthalmol 186: 116-127.
-
Ren R, Li G, Le TD, Kopczynski C, Stamer WD, et al. (2016) Netarsudil increases outflow facility in human eyes through multiple mechanisms. Invest Ophthalmol Vis Sci 57(14): 6197-6209.
-
Chauhan BC, Drance SM (1992) The relationship between intraocular pressure and visual field progression in glaucoma. Graefes Arch Clin Exp Ophthalmol 230(6): 521-526.
- Screening of Hospital Staff During World Glaucoma Week in a Tertiary Eye Care Centre
- Angioid Streaks with Macular Neovascularization: Clinical Insights from Two Cases
- Giant Kissing Naevus: An Oculoplastic Challenge
- Why Freedom of Vision Should Not Cost the Freedom of Feeling - LASIK in the Climate of Change
- Asymmetric Optic Nerve with Small Disc and Large Cup: A Rare and Challenging Case of Unilateral Optic Nerve Hypoplasia
- Large Angle Exotropia in a Child: A Case Report