Replacing Sub-Therapeutic Antibiotics in Feed with Consumer Acceptable Performance Enhancers
Since introduction in the late 1940s, the role of antibiotics in animal production has changed. Originally a means of combating illnesses and maintaining the health of flocks and herds, it was soon recognized that antibiotics could drastically increase productivity and financial return through enhanced and expedited weight gain. Since then criticism has been leveled at the use of antibiotics at sub-therapeutic levels to promote growth and feed conversion efficiency. Although the recent demonstration that plasmid genes encoding for resistance are present in the environment and feces of swine and in carcasses there has been little evidence other than point-of-sale surveys that livestock are contributing to emerging drug resistance among bacterial pathogens affecting humans. Irrespective of the lack of firm scientific evidence that the use of antibiotics in intensive livestock production is directly contributing to drug resistance in hospital and community settings there is a wide perception among consumers that lax regulation over sub-therapeutic administration to food animals is deleterious to public health. Accordingly use of antibiotics for performance enhancement was banned in the EU in 2006 and in the US effective January 2017. Administration of antibiotics for therapy or prophylaxis is now strictly regulated in the EU and the U.S and subject to veterinary prescription applying Prudent Use Principles. Multiple stakeholders must be considered as food production responds to new legislation and rules to limit antibiotic use by farmers, producers, consumers, the medical profession and veterinarians. This paper identifies possible replacement modalities that are acceptable to consumers and the food industry without detrimental effects on animal health and performance. The five criteria producers should consider before adopting alternatives to antibiotics are reviewed. Alternatives include but are not limited to probiotics, prebiotics, short and medium chain fatty acids, enzyme feed supplements, essential oils and botanicals. The paper stresses that no single additive will replace the declining benefits of sub-therapeutic administration of antibiotics. It will be necessary in the future to create programs with a holistic approach to replacement of antibiotics in conformity with EU and U.S. restrictions. Accordingly greater attention should be applied to management, control of immunosuppressive viruses and protozoal parasites, nutrition and the selection of suitable genetic strains to achieve sustainable and safe production of livestock.
A Connolly 1* and S Shane 2
aconnolly@Alltech.com genetic strains to achieve sustainable and safe production of livestock.
Conceptual Paper
Open Access Journal of Veterinary Science & Research
Introduction
Antibiotics were first administered to livestock in the late 1940s when residue from the fermentation process used to produce tetracyclines and streptomycin became available to feed compounders. The remarkable improvement in growth rate and feed conversion efficiency at the time was attributed to the suppression of pathogens by the residual antibiotic. In addition, the fermented substrate supplied nutrients which were lacking in diets formulated applying then current knowledge of nutrient specifications for animals used in production. The practice of sub-clinical administration of antibiotics for growth promotion continued for almost 40 years with acceptance by regulatory agencies worldwide. During the past twenty years there has been a concerted effort to limit the routine application of antibiotics for non-therapeutic, subclinical administration to broiler and turkey flocks as well as pigs, beef, dairy, etc. The motivation has been growing consumer perceptions and scientific evidence that the practice of feeding antibiotics at low levels to food animals contributes to transmissible drug resistance among human bacterial pathogens. Concern over the administration of antibiotics to livestock was formalized in the Swann Report, entitled Use of Antibiotics in Animal Husbandry and Veterinary Medicine released in 1969. The Joint Committee determined that administration of antibiotics to livestock represented a potential risk to human and animal health based on the emergence of antibiotic resistant pathogens. Proposals advanced by the Joint Committee included banning of the addition of tetracycline and penicillin to animal feeds and establishing stricter control of medication through veterinary prescription. Following increasing concern over the deleterious effect of routine sub-clinical use for performance enhancement, led by the Nordic nations, the EU implemented a ban on all antibiotics in animal feed for the purposes of stimulating growth and enhancing feed conversion efficiency effective January 1st 2006. Specified antibiotics were permitted under veterinary prescription for therapeutic and preventive purposes. Parallel developments in the U.S. proceeded at the level of Congress through the U.S. Food and Drug Administration, designated as the major regulatory body. In 2003 the FDA issued Guidance Document 152 listing antibiotic classes and compounds of critical and major medical importance as used in human medicine. The document proposed restricting in-feed compounds to a Veterinary Feed Directive (VFD), a concept previously established during the late 1990s. The Guidance Document recommended elimination of over-the-counter drugs and placed the onus on Veterinarians to prescribe antibiotics in water and issuing VFDs for feed supplementation. It was required that Veterinarians follow FDA Prudent Use Principles with regard to VFDs and prescriptions. In April 2012, the FDA issued Guidance Document 209 which eliminated all claims for growth promotion or performance enhancement for antibiotics then approved for this purpose. Basically the Guidance Document restricted use of drugs regarded as “Critical” and “Very Important” for human therapy. Only the ionophore anticoccidials and bacitracin were regarded as not being of importance to human medicine. In support of the recommendations outlined in Guidance Document 209 the FDA referenced the 1969 Swann Report, a 1980 National Academy of Sciences monograph entitled The Effects on Human Report of Sub-therapeutic Use of Antimicrobial Drugs in Animal Feeds and a 1997 World Health Organization report, The Medical Impact of Antimicrobial Use in Food Animals. FDA Guidance Document 213 dated December 2013 required adoption of Judicious Use Principles and required sponsors of existing drugs approved for sub-therapeutic administration to voluntarily withdraw label directions. The Guidance Document specified both prescriptions for antibiotic administered in drinking water and Veterinary Feed Directives to be justified according to the following criteria:
- Evidence of effectiveness of an antibiotic for a specific application.
- Administration of an antibiotic to flocks to be consistent with accepted veterinary practice.
- Selection of an antibiotic to be linked to a specific etiologic (causing) agent.
- Administration targeted to animals at risk of developing a specific disease.
- Antibiotics common to human use to be prescribed or the subject of a VFD, only if no alternative medication or therapeutic modality is available. Although the US FDA Guidance Documents banned administration of antibiotics for growth promotion by January 1, 2017, many broiler and turkey producers preemptively withdrew antibiotics for growth enhancement prior to the statutory cut-off date. Cargill Inc. removed growth promoting antibiotics from diets fed to specific brands of their turkey products in July 2014.
Open Access Journal of Veterinary Science & Research
Tyson Foods eliminated antibiotics common to human therapy in May 2015.Foster Farms introduced a brand raised without antibiotics in June 2015. A similar announcement was made by Perdue Farms on July 2015 and in October 2016 the company announced that it had initiated a USDA-Verified No Antibiotics Ever program for all chicken products, and the swine and beef industries are following suit. Actions by producers were in large measure stimulated by demands by major quick service restaurant chains and food-service distributors to initially cease administration of sub-therapeutic antibiotics, followed by elimination of antibiotics common to human therapy for prevention or treatment of disease and then to completely eliminate antibiotics from their supply chains. An annual fast food report card is produced by CNN Health [1].
Mechanism of Antibiotic Action
A number of studies have demonstrated mechanisms for the benefits of antibiotics through their action in the intestinal tract of chickens. These include:
- Suppression of pathogens in the microbiota.
- Reducing deleterious metabolites elaborated by the microbiota.
- Modulating inflammatory response in the intestinal mucosa as a result of stimulation by pathogens.
- Eliminating competition for nutrients between the microbiota and host. It is generally accepted that over the past decade in the US and the EU the beneficial impact of antibiotics on animal performance has been reduced. Modalities contributing to the declining value of antibiotics include satisfying nutritional requirements with an emphasis on micronutrients and enzyme supplements; achieving optimal control of litter moisture through adequate ventilation; suppression of coccidiosis and immunosuppressive viral infections. Discontinuing sub-therapeutic antimicrobial administration was reviewed in Denmark [2]. A retrospective study was conducted on 6,815 flocks harvested from November 1995 to July 1999 in cooperation with the Danish Poultry Council. It was demonstrated that administration of antibiotics had no beneficial effect on the biomass (kilograms of live broilers produced per square meter of housing) or livability as a result of withdrawal of antibiotic growth promoters. There was a 0.06 improvement in feed conversion efficiency (kilograms of feed consumed per kilogram of live weight). The data presented in the review demonstrated that if translated to current U.S. production parameters, antibiotic administration would cost 0.7 cents per bird assuming a live weight of 1.8 kg and a feed conversion efficiency of 1.8.The cost of supplementation with antibiotic growth promoters included in both the starter and finisher diets would amount to between 0.8 to 1.0 cents per bird approximating an unacceptable benefit- to-cost ratio of unity. Perdue Farms undertook a prospective evaluation of antibiotics extending from 1988 to 2001 involving 168 paired-house comparisons with seven million broilers in North Carolina and the Delmarva region. The study showed that withdrawing antibiotics reduced livability by 0.17 percent, live weight was depressed by 15 grams per bird and feed conversion efficiency was reduced by 0.014. There was no difference in either condemnation rate or intensity of skin pigment [3]. Subsequently the data was analyzed by a group at the Johns Hopkins Bloomberg School of Public Health [4] confirming the lack of any positive benefit-to-cost relationship attributable to antibiotic administration. In the swine industry, several studies have estimated the potential economic impact of banned AGPs to be wide ranging, from $1.37 [5] to $2.33/pig [6] to even $4.50/pig [7]. Other impacts include 9% decrease in net profits [8], 2% increase in production costs [6], and 4.5% increase in production costs [7].
Mechanisms of Antibiotic Resistance
Some bacteria including Enterobacter faecalis are naturally resistant to specific antibiotics. This is specifically the case with virginiamycin, contrasted with the susceptibility of Enterobacter faecium to the compound. Accordingly field surveys of processed poultry which denotes “half of the Enterobacters isolated were resistant to virginiamycin” is a spurious finding since E. faecalis is not suppressed by virginiamycin. Acquired resistance can arise either by point mutation with subsequent horizontal transmission or by plasmid transfer. This mechanism is possibly a means by which drug resistance developed in livestock populations can be transmitted to humans. The recognition of the role of the mcr-1 gene which creates a plasmid mediated resistance to colistin is a well-documented mechanism (Liu et al., 2015). The mcr-1 gene was present in E. coli isolated in 15 percent of 520 samples of raw meat in a survey conducted in China over the period 2011 through 2014. Under laboratory conditions the mcr-1 plasmid mediated gene was transmitted from E. coli to Klebsiella pneumoniae and Pseudomonas aeruginosa. Subsequently the mcr-1 gene Open Access Journal of Veterinary Science & Research
that has been identified in Europe and in the U.S. has been isolated from a human patient and a number of hogs at slaughter during a structured surveillance program conducted by the Centers for Disease Control, the U.S. Department of Agriculture and the Food and Drug Administration. Antibiotic resistance can arise from the following mechanisms:
- Drug inactivation such as occurs with inhibition of penicillin by beta lactamase elaborated by pathogens.
- Alteration of binding sites with methicillin-resistant Staphylococcus aureus an example of this mechanism.
- Reducing intercellular drug concentration can be implemented through efflux pumps. The mechanisms by which low levels of antibiotics, heavy metals and anti-bacterial maintain resistance among bacterial populations has yet to be defined but experimental data suggests that plasmid genes are responsible [9]. The study conducted on nosocomial isolates of Klebsiella pneumonia and E. coli obtained from patients in Upsalla hospitals demonstrated persistence of resistance to a wide range of antibiotics when bacteria were maintained at low levels of exposure to antibiotics over prolonged periods such as would be encountered in contaminated hospital or farm environments. It was also possible to establish resistance by transferring genes from plasmids to chromosomes. The issue of transmissible drug resistance was the subject of a May 2016 feature article in The Economist. The article reviewed the report of the UK Parliamentary Committee chaired by Lord O’Neil of Gatley which predicted that increasing drug resistance would reduce worldwide gross domestic product by 2 to 3 percent and result in annual costs exceeding $20 billion annually by 2030. It also estimated that by 2050 potentially 10 million people could die from drug resistant infections per annum. The O’Neil Report revisited the Swann Report reinforcing suggestions made in 1969. The O’Neil Committee recommended improved training of physicians in the use of antibiotics, banning the indiscriminate sale and administration of antibiotics to livestock and for patients, especially in non-industrialized nations, surveillance of antibiotic resistance among human populations and implementation of awareness campaigns to reduce pressure on physicians to prescribe antibiotics when not clinically indicated. It is ironic that Sir Alexander Fleming, who identified the antibacterial action from an extract of Penicillinum which led to the antibiotic revolution of the late 1940s, categorically warned of the dangers of developing resistance among bacterial pathogens in his Nobel acceptance speech.
Producing Poultry and Livestock without Growth-Promoting Antibiotics
The potential problems following withdrawal of routine sub-therapeutic antibiotics in feed include clinically apparent and sub-clinical clostridial enterotoxemia complex including necrotic enteritis, cholangio hepatitis, clostridial gangrenous dermatitis and reduced growth rate and feed conversion efficiency. Consumer Acceptable Performance Enhancers (CAPES) which function as alternatives to antibiotics are commercially available and can be used as part of a holistic approach to achieving the genetic potential of broilers. In addition to feed additives, a range of concurrent management procedures are required to compliment the beneficial effect of feed additives on intestinal microflora. These include:
- Reducing the effect of immunosuppressive viruses including infectious bursal disease and Marek’s disease.
- Control of coccidiosis which serves as a precursor of clostridial enterotoxemia [10].
- Maintaining acceptable levels of ventilation to maintain dry litter.
- Feeding balanced diets formulated without animal protein byproducts or animal fat.
- Adding supplementary enzymes to diets to enhance digestion and specifically degrade non-starch polysaccharides. CAPES which in part replace antibiotics can be classified into six categories: 1. Probiotics including live bacterial cultures principally Lactobacilli, Bacilli, and Bifido bacterium. 2. Prebiotics, principally oligosaccharides derived from the cell wall of Saccharomyces cerevisiae. (If probiotics and prebiotics are combined, these commercial products are referred to as symbiotic.) 3. Botanicals comprise extracts of herbs which influence the composition of the bacterial flora and may suppress pathogens including Eimeria spp. 4. Essential oils including oregano and cinnamon. 5. Short-chain fatty acids including butyrate which promote beneficial flora through the release of butyric acid. Medium chain fatty acids also have a Open Access Journal of Veterinary Science & Research
potentially beneficial effect on the intestinal microbiota. 6. Enzymes commonly produced by bacteria or fungus using submerged fermentation or solid state fermentation accelerate digestive reactions in the body that might otherwise proceed very slowly or not at all.
Probiotics
Live cultures include a range of Lactobacillus spp. including L. acidophilus, L. casei, L. reuteri, L. salivarius and L. lactis. Bifido bacteriums pp. includes B. bifidum, B. lactis, and B. animalis. Other organisms which have been incorporated into probiotic mixtures include Lacto coccuslactis, Pediococcus acidilactici and Streptococcus thermophila, Propionibacterium freudenreichii and Bacillus cereus [11]. The mechanism of action of probiotics has not been completely elucidated and in all probability individual strains have specific actions under varying conditions involving nutrient composition rate of feed passage and the presence of pathogens which affect intestinal integrity and function. It is generally accepted that probiotics are capable of contributing to stability of the intestinal flora of the distal ileum and cecum through the following mechanisms:
- The earliest hypothesis to explain the beneficial action of probiotics involved competitive exclusion by blocking receptor sites on enterocytes.
- Subsequently it was proposed that probiotic strains compete with pathogens for essential nutrients.
- Probiotic bacteria in the lower intestinal tract produce short chain organic acids including acetic and lactic acids which lower the pH of the lumen to the detriment of pathogens including Salmonella spp.
- Probiotic cultures can enhance the barrier functions of the mucosa. Lactobacillus spp. is involved in up- regulation of genes producing intestinal mucins which prevent potentially pathogenic bacteria adhering to enterocytes [12].
- Lactobacillus spp. has the capability to regulate genes encoding for adherence junction proteins which increase tissue resistance to pathogens.
- Lactobacillus spp. demonstrates a mannose-specific adhesion to enterocytes stimulating the release of cytokines which enhance mucosal host immunity [13].
- Probiotics elaborate antimicrobial peptides. Bacteriocins inhibit cell wall synthesis by pathogens.
Defensins are antimicrobial peptides which disrupt the cytoplasmic membranes of susceptible micro- organisms [12].
- It has been demonstrated that probiotic strains of Biofido bacterium and Lactobacillus can enhance local immunity by increasing the phagocytic capacity of macrophages. Probiotics also enhance natural killer cell activity possibly mediated throughinterleukin- 12.Probiotic strains stimulate the production of IgA by lymphoid cells in the intestinal sub-mucosa.
- Specific Bacillus subtilis strains are capable of germinating in the small intestine from ingested spores. After colonizing the lower intestinal tract, selected B. Subtilis strains showed activity equivalent to bacitracin to prevent necrotic enteritis [14]. The toxicity of certain bacillus strains was reviewed during the European Union's feed additive approval process because of the risk of potentially toxic metabolites [15]. In evaluating a probiotic as a candidate for replacement of antibiotics [16], it is necessary that the selected strains individually or in combination should satisfy the following criteria:
- A probiotic culture must be a pathogenic, characterized as generally regarded as safe.
- Probiotic Lactobacillus spp. and other genera must have the ability to adhere to mucosal cells of the target species. Adherence is associated with competitive exclusion as demonstrated by studies comparing the effect of strains of Lactobacillus fermentum on preventing colonization by various Salmonella serovars including S. Enteritidis.
- Suitable strains should proliferate after adhesion to achieve colonization. Probiotic candidates should survive physical conditions associated with mixing of feed although it is accepted that unless specifically protected, probiotics are destroyed by the temperature applied in pelleting. Accordingly for broilers, post-pellet spray of live cultures is required. Alternatively probiotics can be administered by coarse aerosol at the hatchery, administered as a gel suspension which is ingested during transport in chick boxes or may be administered in drinking water after placement of chicks on a farm.
Prebiotics
Prebiotics are defined as non-digestible food ingredients that benefit the host by modifying the composition of the intestinal microbiota [17]. Prebiotics
Open Access Journal of Veterinary Science & Research
must satisfy the requirement that they are not hydrolyzed after ingestion or absorbed perse in the gastrointestinal tract. Prebiotics must promote the proliferation of beneficial components of the microbiome to the benefit of the host. Within the context of monogastric nutrition, prebiotics should serve as substrates for fermentation in the ileum and cecum, lead to higher levels of short chain fatty acid evolution by the microbiome, contributing to low pH levels in the lower intestinal tract. Virtually all prebiotics are carbohydrates with the oligosaccharides predominating. Fructo oligosaccharides and galacto oligosaccharides have been evaluated in humans and laboratory animals, for poultry and other monogastric livestock [18]. Mannan oligosaccharides (MOS) derived from the cell wall of Saccharomyces cerevisiae were introduced commercially in the early 1990s but adoption by the animal industry intensified after 2000 based on the need to replace antibiotic growth promoters. Commercial Mannan oligosaccharide additives such as Bio-Mos® are derived from alpha-D-mannanose polysaccharides comprising alpha-(1,2)-and alpha-(1,3)- D-mannose branches which form alpha-(1,6)-D-mannose chains [19]. The MOS-protein conjugates which are formed are hydrophilic and have branch-like structures to attach to receptors on enterocytes and to Type-1 fimbriae of pathogens. Inclusion of MOS in diets has a significant effect on the development of the intestinal mucosa, promoting longer villi thereby increasing the surface for secretion of mucus, enzymes and also absorption of nutrients. It has been demonstrated that MOS inhibits attachment of pathogenic bacteria to enterocytes, thereby inhibiting colonization [20].The mechanism of action of MOS is multifactorial with the principal effect relating to the composition of the microbiome. Samples obtained from commercially reared broilers at weekly intervals through 35 days of age showed that supplementation with MOS favored Bacteroidetes, which replaced Firmicutes. By altering the composition of the intestinal flora of the cecum, beneficial effects which are manifested in improved growth rate or enhanced feed conversion efficiency can be quantified [21] with specific relation to energy metabolism [22]. Bacteroidetes are hydrolytic in their activity and are profuse elaborators of short chain fatty acids and are capable of degrading complex carbohydrates. The environment of the cecum in chickens receiving MOS supplementation disfavors proliferation of Bacillus spp. and Clostridium spp. [23]. The effect of MOS on the immune system has been extensively studied with special reference to secretion and composition of intestinal mucosa which serves as a protective barrier against pathogens. MOS stimulates an increase in T-cells attributed to up-regulation of gene expression in cells of the intestinal crypts [24]. The performance improvements in poultry fed MOS as a dietary supplement have been assessed in meta analyses [25, 26]. A total of 44 trials were evaluated with MOS fed at levels ranging from 0.5 to 2 kg per metric ton of feed. Supplementation with MOS improved body weight by 2 percent enhanced feed conversion efficiency by 2.3 percent and reduced mortality by 22 percent compared with controls. A meta-analysis based on published articles evaluating MOS supplementation in turkeys demonstrated a significant improvement of 2 percent in body weight and reduction of 1.5 percent in feed conversion [27]. Analyses performed in swine indicating that piglets fed MOS supplementation, particularly those with a slow growth rate, can demonstrate increased growth, particularly during the first two week post-weaning [28]. Similar beneficial results were found in calves which demonstrated an improved rate of gain per day of 0.07kg and an overall weight gain of 3.42 kg or 15% [29]. In 2009 a second generation of a MOS-additive, termed Mannose-Rich Fraction (MRF) became available commercially as c. Concurrently, Alltech, the originator of the patented supplement developed an ELISA-linked mucin adherence assay (ELMAA). This allows quantification of the additive in pre-mixes and complete feed representing an advance in quality assurance and traceability. The benefits of MOS are evident with Actigen® although commercial experience confirms more intense action especially with regard to enhancing nutrient utilization, suppression of inflammation and performance parameters of piglets infected with porcine epidemic diarrhea, caused by a coronavirus [30]. In evaluating the immuno-modulatory action of the mannose-rich fraction, it was shown that gene expression of toll-like receptors such as TLR4, cytokines IL-12p35 and interferon in the ileum were down-regulated. The immune-mediators involved are the Th1-helper and Th2- helper pathways resulting in the anti-inflammatory response [31].The practical-field impact of supplementation with Actigen® in comparison to zinc bacitracin showed protection from necrotic enteritis applying a clostridial challenge model. The role of MOS in improving growth rate and feed conversion efficiency is supported by over 733 publications or platform presentations and the product has been in use for over two decades attesting to safety, efficacy and commercial acceptability.
Open Access Journal of Veterinary Science & Research
The application of symbiotic combining probiotics and prebiotics demonstrates benefits in growth rate equivalent to antibiotic supplementation and effects of combinations were superior to individual products [18].
Essential Oils
Essential oils are aromatic plant extracts subjected to varying degrees of purification and concentration. Most essential oils are complex mixtures of plant metabolites comprising phenylpropenes and terpenes [32]. The composition of a specific essential oil will vary according to the geographic location, time of harvest, and method of extraction although steam distillation is the most commonly employed process [33]. Controlled studies on the effects of the various essential oils were conducted during the first decade of the present century. Candidates evaluated included oregano, thymol, cinnamon, carvacrol and coriander. The ban on antibiotic growth promoters in the EU proposed in 2003 stimulated research on alternatives to sub- therapeutic administration of antibiotic compounds. The interest in essential oils related to the antibacterial action which is associated with the phenolic components. It was demonstrated that oregano and cinnamon essential oils demonstrated a higher antibacterial activity against Gram-positive genera than Gram-negative species although some reference strains and clinical isolates are susceptible to both essential oils. An important consideration is that susceptible species of bacteria do not develop resistance to the antibacterial activity of essential oils after 50 passages in the presence of low concentrations of the active ingredient. In this respect, cinnamon essential oil is superior to oregano in which some resistance was observed against Proteusmirabilis. This suggested that cinnamon is a more reliable compound for long-term administration [34]. The mode of action of essential oils in animal nutrition has yet to be defined although it is suggested that the compounds may enhance secretion of pancreatic amylase, trypsin and maltase in the intestinal tract resulting in more efficient digestion [35]. A second mechanism by which essential oils can enhance production, relates to antimicrobial activity. Essential oils containing high proportions of phenolic compounds including carvacrol, eugenol and thymol have a beneficial effect on the composition of the intestinal microbiota. It is hypothesized that the phenolic compounds can degrade the outer membrane of gram- negative bacteria releasing lipopolysaccharides thereby increasing the permeability of the cytoplasmic membranes [36]. Proliferation of Clostridium perfringens can be inhibited by administering a blend of essential oils including mixtures of carvacrol and thymol or thymol and eugenol [37]. Inactivation of Clostridium perfringens toxins by digestive enzymes is a possible mechanism to explain the beneficial effect of essential oil supplements in preventing necrotic enteritis. A similar result was obtained in trials conducted using a necrotic enteritis challenge model which showed a similar effect compared to bacitracin [38]. Essential oils appear to improve clinical response following administration of an oocyst vaccine to prevent coccidiosis. This is especially relevant for broilers produced according to drug-free or organic specifications [39]. Inconsistencies in results under field conditions arising from application of essential oils relates to the absence of uniformity, effective traceability and a dearth of analytical procedures. Variability in active ingredients in commercial complex mixtures may result in either heightened or suboptimal effects. There is little information on the potential deleterious interaction between essential oils, probiotic cultures, other nutrients or additives.
Botanicals
The effect on the immune system and performance parameters following administration of herbs has been the subject of considerable speculation. Predictably research has been conducted in China [40] and India [41] where there is a long tradition of administering botanicals for both therapy and stimulation of growth. The action of many botanicals on the intestinal microbiota and the immune system is based on their content of essential oils which have stimulatory effects on tissues secreting digestive enzymes in addition to suppressing Gram- positive bacteria. A number of plants including Artemisia spp. contain artemisin in which demonstrates an inhibitory action against Eimeria oocysts, preventing sporulation and thereby preventing coccidiosis. The same mechanism has been demonstrated for tannins in pine bark [42]. Restraints associated with the application of essential oils, specifically with respect to consistency and quality, are even greater with botanicals. In addition, it is possible that at the levels of dietary inclusion which are effective, abnormal flavors can be induced in egg and poultry meat which degrades consumer acceptability. It is possible that Open Access Journal of Veterinary Science & Research
although botanicals are classified as GRAS, deleterious alkaloids or other components may prove toxic to poultry or result in residues in eggs and meat.
Organic Acids
Organic acids have been promoted as a potential replacement for growth promoting antibiotics in animal production especially after the EC announced a proposed ban on antibiotic growth promoters in 2013 (EC Regulation 1831/2003) [43]. Organic acids are also termed “volatile fatty acids”; “carboxylic acids” or “fatty acids”. The most commonly used organic acids are monocarboxylic, comprising a carboxyl group and a short chain extending from 1 to 6 carbon units. When included in feed, organic acids suppress pathogens, modify the microbiome, stimulate immunity and have the capacity to enhance the efficiency of digestion [44]. To be effective it is necessary to encapsulate short chain fatty acids to enable them to exert their activity in the distal intestinal tract. The action of organic acids on bacteria relates to their ability to transit the cell wall and then to disassociate in the internal cytoplasmic milieu of the bacterium. The ability to disassociate is a function of the pKa which is the pH at which half of the molecules will disassociate. Since most bacteria have a pH approaching neutrality, the five short chain fatty acids will liberate H+ which acidifies the cytoplasm and disrupts the acid-based regulatory mechanism of the cell. The anion portion of the molecule also interferes with metabolism through oxidative inhibition of enzymes, resulting in either stasis or death of the cell [45]. Since pH influences the inhibitory effect of organic acids, at low pH values the potential impact on pathogens is greater as denoted by values for minimum inhibitory concentration (MIC) of the organic acid. If an organic acid is to be effective in the terminal ileum and cecum, it is necessary to maintain a low pH value which is created by beneficial components of the microbiome including the lactic acid-producing flora. In this respect combination of prebiotics, probiotics and organic acids are synergistic [46]. It has been demonstrated that butyric and propionic acids inhibit virulence factors produced by pathogens including Salmonella and Listeria [47]. Sodium butyrate must be protected by encapsulation in order to exert any effect in the distal intestinal tract. Although non-protected sodium butyrate will reduce colonization with Salmonella in the crop, protection against Salmonella infection in the distal intestinal tract is only provided by vegetable-oil or gel encapsulation to prevent degradation of the additive in the proventriculus and ventriculus. Mixtures of organic acids including formic and propionic included in feed reduce the level of Salmonella and possibly other potential pathogens in the cecum of chickens [48]. Organic acids can alter the composition of the ileal and cecal microbiota when added in either feed or drinking water [49]. The level of Lactobacilli was increased significantly following administration of a combination of formic and propionic acids adherent to a silica-based carrier. E. coli was inhibited by formic acid without affecting Lactobacilli as indicated by a ten-fold difference in MIC value between coliforms and Lactobacilli [50]. Organic acids have the ability to modify the morphology of the intestinal mucosa. Formic acid included in diets increased the length of villi corresponding to the effect of a vilamycin, a commonly used feed-additive antibiotic growth promoter in the EU prior to 2016. Both the formic acid and antibiotic treatments were significantly superior with respect to villus morphology than control-fed broilers at 42 days of age. A similar effect was noted when a combination of formic and propionic acids was added to the diets of broilers resulting in enhanced growth and superior feed conversion efficiency compared to non-supplemented controls. The beneficial effect from administering organic acids was attributed in part to raising the metabolizable energy value of diets through enhancing the digestibility of crude protein, ether extract and crude fiber [51]. Proteolysis is enhanced by increased secretion of pepsin possibly as a result of stimulation of gastrin and cholecystokinin release. Stimulation of the exocrine pancreas increases secretion evidenced by an elevation of the concentrations of trypsinogens and peptidases in the duodenal lumen. Organic acids may have a positive effect on tissue immunity. Combinations of organic acid in combination with phytase enhanced levels of immuno-globulin G as both a primary and secondary response [52]. Based on the diverse actions of organic acids, structured trials demonstrated improved growth rate and feed conversion efficiency equivalent to growth promoting antibiotics [43]. Under controlled conditions using a necrotic enteritis challenge model, it was demonstrated that a combination of medium chain fatty acids incorporated in feed was Open Access Journal of Veterinary Science & Research
equivalent to bacitracin in maintaining growth rate and preventing mortality compared to non-medicated controls [53]. Medium chain fatty acids including caproic, capric and lauric acids are more effective than the short- chain compounds in modifying the microbiome of the distal intestinal tract [54]. Despite the effects on reducing the level of pathogens in the terminal intestinal tract, improvement in mucosal absorption area, stimulation of digestive enzymes and an indirect effect on the immune system, field results with organic acids singly or in combination have been inconsistent. This may be due to confounding factors associated with environmental effects including biomass, response to coccidiosis vaccines, suboptimal ventilation and litter management or immune suppression due to sub-clinical viral infections such as bursal disease, reovirus and Marek’s disease.
Recombinant Enzymes
Enzymes are biologically active proteins capable of degrading specific chemical bonds and releasing nutrients for digestion and absorption. Feed additive enzymes accelerate chemical reactions in the intestine which would otherwise proceed very slowly or not at all. Enzymes used in the feed industry are commonly produced by bacteria such as Bacillus subtilis and fungi such as. Trichoderma reesei and Aspergillus niger using a submerged fermentation process. Supplementing monogastric diets with exogenous enzymes to enhance performance has been applied since the1950s. Enzyme supplementation has commonly focused on degrading feed components resistant to endogenous enzymes (i.e. β-glucanase, xylanase, mannanase, pectinase and galactosidase), inactivating anti-nutritional factors (phytase) and supplementing endogenous enzymes that may be present in insufficient amounts (proteases, lipases and amylases). Typical responses have been strongest in poultry followed by swine. The contribution of enzymes alone to growth rate is the result of improved digestibility of nutrients. The primary benefits of enzymes in programs to replace antibiotic Extrapolating laboratory-generated data into quantifiable field results frequently represents a challenge, especially due to the confounding effects associated with commercial production. The fact that organic acids and combinations of prebiotics and probiotics in the form of symbiotic have similar action, it is logical to presume that combinations of acidifiers, oligosaccharides and active probiotics will have more consistent and reproducible effects in their ability to replace antibiotic growth promoters, which are now banned in most industrialized countries. It is stressed that there is no single compound which can replace growth promoting antibiotics. Alternative feed supplements must be administered in a comprehensive approach incorporating adequately balanced diets, suppression of coccidiosis and enhanced management of litter moisture, which in turn is based on ventilation rate, biomass and control over watering systems. Programs such as Seed, Feed and Weed [56] demonstrate how alternative feeding methods can be approached in this comprehensive manner to obtain similar results without the use of antibiotics.
growth promoters have been indirect in action, mediated through the intestinal microflora. The microbiota of the terminal intestine and ceca of poultry can be modified by enhanced digestion of crude protein and fiber in the duodenum and anterior intestinal tract. Incomplete digestion favors the proliferation of potentially deleterious pathogens such as Clostridium spp. to the detriment of beneficial lactose-producing bacteria. Novel enzyme complexes, produced through solid state fermentation, provide additional benefits since they are produced from a single microorganism and as such produce a range of enzymes which are synergistic in their action. Studies have shown that enzyme complexes are more efficient at releasing energy and amino acids from feeds compared to commercial blends of individual enzymes. Complete digestion obviates the accumulation of incompletely degraded feed which serves as a substrate for proliferation of potential pathogens [55].
Conclusion
Open Access Journal of Veterinary Science & Research
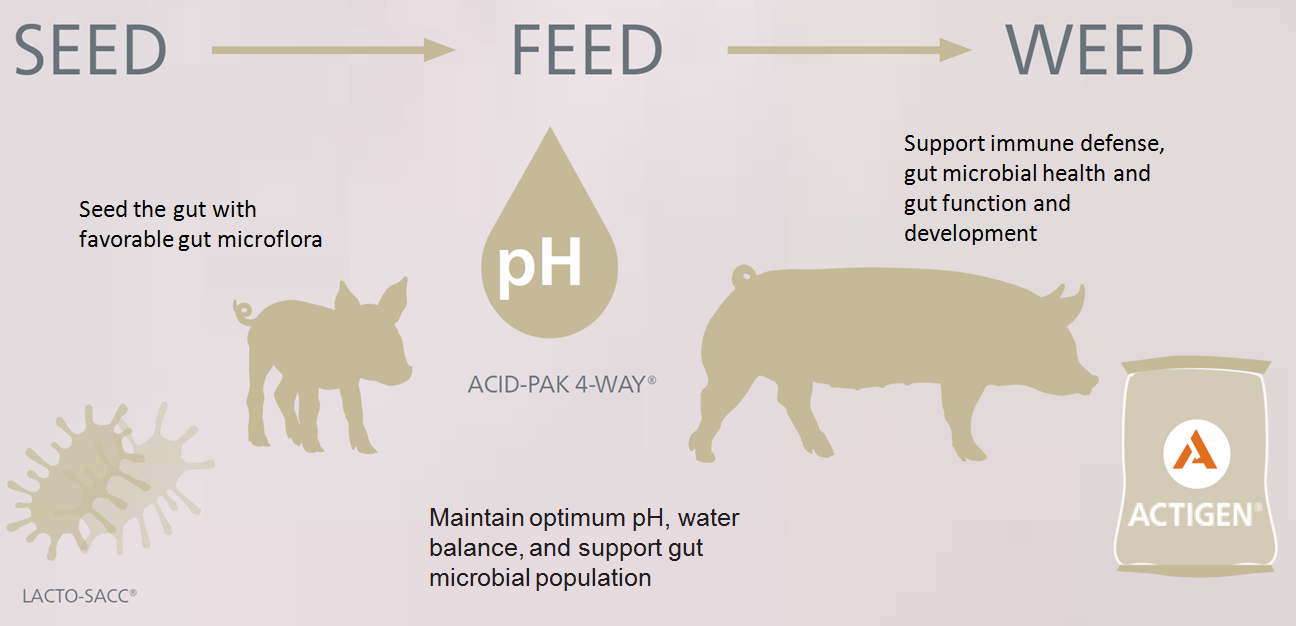
When determining if a feed supplement is appropriate to promote animal health and performance, and offer an alternative to antibiotics as appropriate, McCartney [57] has developed a five-point test one can use:
- Will consumers accept the new ingredient?
- Is the product safe for animals and people (in consumption, administration, etc.)?
- Is it pellet stable?
- Is the mode of action understood?
- Are responses consistent? Positively responses to these five questions either with an individual intervention or with a program can allow producers to arrive at possible replacement therapies that are consumer acceptable without being detrimental to animal performance.
References
-
CNN Health (2016) Antibiotics in the fast food meat supply.
-
Emborg HD, Ersboll AK, Heur OE, Wegener HC (2001) The effect of discontinuing the use of antimicrobial growth promotors on the productivity in the Danish broiler production. Prev Vet Med 50(1-2): 53-70.
-
Engster HM, Marvil D, Stewart-Brown B (2002) The effect of withdrawing growth promoting antibiotics from broiler chickens: A long term commercial industry study. J Applied Poultry Science 11: 431-436.
-
Graham JP, Boland JJ, Silbergeld E (2007) Growth promoting antibiotics in food animal production: An economic analysis. Public Health 122(1): 79-87.
-
Miller GY, Liu X, Paul E, McNamara, Bush EJ (2005) Farm-Level Impacts of Banning Growth-Promoting Antibiotic Use in U.S. Pig Grower/Finisher Operations. Journal of Agribusiness 23(2).
-
Brorsen BW, Brorsen, Lehenbauer T (2002) Economic Impacts of Banning Subtherapeutic Use of Antibiotics in Swine Production. Journal of Agricultural and Applied Economics 34(3): 489-500.
-
Hayes DJ, Jensen HH (2003) Lessons from the Danish Ban on Feed-Grade Antibiotics. CARD Iowa State University.
-
Miller GY, McNamara PE, Bush EJ (2003) Productivity and Economic Effects of Antibiotics Used for Growth Promotion in Pork Production. Journal of Agricultural and Applied Economics 35: 469-482.
-
Gulberg E, Albrecht LM, Karlsson C, Sandegren L, Andersson DI (2014) Selection of a multidrug resistance plasmid by sublethal levels of antibiotics and heavy metals. mBio 5: e0918-1014.
-
Shane SM, Gyimah J, Harrington KS, Snyder TG (1985) Etiology and pathogenesis of necrotic enteritis. Vet Res Communications 9(4): 269-287. Open Access Journal of Veterinary Science & Research
-
Syngai GG, Gopi IR, Bharali R, Dey S, Lakshmanan GMA, et al. (2016) Probiotics-the versatile functional food ingredients. J Food Sci Technol 53(2): 921-933.
-
Hardy H, Harris J, Lyon E, Beal J, Foey AD (2013) Probiotics, prebiotics and immunomodulation of gut mucosal defenses: homeostasis and immunopathology. Nutrition 5(6): 1869-1912.
-
Hemaiswarya S, Raja R, Ravikumar R, Carvalho IS (2013) Mechanism of the action of probiotics. Brazil Archiv Biol Technol 56(1): 113-119.
-
Tellez G, Pixley C, Wolfenden RE, Layton SL, Hargis BM (2012) Probiotics/direct fed microbials for Salmonella control in poultry. Food Research International 45: 628-633.
-
From C, Pukall R, Schumann P, Hormazabal V, Granum PE (2015) Toxin-Producing Ability among Bacillus spp. Outside the Bacillus cereus Group. Applied and Environmental Microbiology 71(3): 1178-1183.
-
Smith JS (2014) A review of avian probiotics. J Avian Medicine and Surgery 28(2): 87-94.
-
Spring P, Wenks C, Connolly A, Kiers A (2015) A review of 733 published trials on Bio-Mos®, a mannan oligosaccharide and Actigen®, a second generation mannose rich fraction, on farm and companion animals. J Applied Animal Nutrition 3: 1- 11.
-
Gagia F, Mattarelli P, Biavati B (2010) Probiotics and prebiotics in animal feeding for safe food production. Int J Food Microbiology 141(1): 515-528.
-
Vinogradov E, Petersen B, Bock K (1998) Structural analysis of intact polysaccharide mannan from Saccharomyces cerevisiae yeast using 1H and 13C NMR spectroscopy at 750 MHz. Carbohydrate Research 307: 177-183.
-
Firon N, Ofek I, Sharon N (1983) Carbohydrate specificity of the surface lectins of Escherichia coli, Klebsiella pneumonia and Salmonella typhimurium. Carbohydrate Research 120: 235-249.
-
Corrigan A, de Leeuw M, Penaud-Frezet S, Dimova D, Murphy RA (2015) Phylogenetic and functional alterations in bacterial community compositions in broiler ceca as a result of mannan oligosaccharide supplementation. Applied and Environmental Microbiology 81(10): 3460-3470.
-
Singh KM, Shah TM, Deshpande S, Jakhesara SJ, Koringa PG, et al. (2012) High throughput 16S rRNA gene-based pyro sequencing of fecal microbiota of high FCR and low FCR broiler growers. Mol Biol Rep 39(12): 10595-10602.
-
Paredes CJ, Alasker KV, Papoutsakis ET (2005) A comparative genomic view of clostridial sporulation and physiology. Nat Rev Microbiol 3(12): 969-978.
-
Uni Z (2007) Physiological dynamics at the intestinal brush border. In: Lyons TP (eds), Nutritional biology in the feed and food industries. Proceedings of Alltech’s 23rd Annual Symposium, Nottingham University Press, pp. 131-138.
-
Hooge DM (2011) Meta-analysis summary of broiler chicken trials with dietary Actigen® (2009-2011). Int J Poultry Science 10(10): 819-824.
-
Hooge DM, Kiers A, Connolly A (2013) Meta-analysis, summary of broiler chicken trials with dietary Actigen® (2009-2012). Int J Poultry Science 12(1): 1- 8.
-
Hooge DM (2004) Turkey pen trials with dietary mannan oligosaccharide: meta-analysis, 1993-2003). Int J Poultry Science 3(3): 179-188.
-
Miguel JC, Rodriguez-Zas SL, Pettigrew JE (2004) Efficacy of a mannan oligosaccharide (Bio-Mos®) for improving nursery pig performance. J Swine Health Prod 12(6): 296-307.
-
Hooge DM (2006) MOS may boost calf gain: Mannan oligosaccharides had beneficial effects on dairy calf weight gain and total starter feed intake, but mortality needs to be further evaluated. (mannan oligosacchides). Feedstuffs. Miller Publishing Co 2006. High Beam Research 7
-
Che TM, Song M, Liu Y, Johnson RW, Kelley KW et al. (2012) Mannan oligosaccharide increases serum concentrations of antibiotics and inflammatory mediators in weanling pigs infected with porcine reproductive and respiratory syndrome virus. J Animal Science 90(8): 2784-2793.
-
Munyaka PM, Echeverry H, Yitbarek A, Camelo-Jaimes G, Sharif S, et al. (2012) Local and systemic innate immunity in broiler chickens supplemented with Open Access Journal of Veterinary Science & Research yeast-derived carbohydrates. Poultry Science 91: 2164-2172.
-
Brenes A, Roura E (2010) Essential oils in poultry nutrition: Main effects and modes of action. Animal Feed Science and Technology 158(1-2): 1-14.
-
Greathead H (2003) Plants and plant extracts for improving animal productivity. Proc Nutr Soc 62(2): 279-290.
-
Becerril R, Nerin C, Gomez-Lus R (2012) Evaluation of bacterial resistance to essential oils and antibiotics after exposure to oregano and cinnamon essential oils. Food borne Pathogens and Disease 9(8): 699- 705.
-
Jang IS, Ko HY, Ha JS, Kim JY, Kang SY, et al. (2004) Influence of essential oil components on growth performance and the functional activity of the pancreas and small intestine in broiler chickens. Asian Australasian J Animal Science 17(3): 394-400.
-
Helander IM, Alakomi HL, Latva-Kala K, Matilla- Sandholm T, Pol I, et al. (1998) Characterization of the action of selected essential oil components on Gram-negative bacteria. J Agric Food Chem 46(9): 3590-3595.
-
Mitsch P, Zitterl-Eglseer K, Kohler B, Gabler C, Losa R, et al. (2004) The effect of two different blends of essential oil components on the proliferation of Clostridium perfringens in the intestine of broiler chickens. Poultry Science 83(4): 669-675.
-
Sims MD, Williams PG, Frehner M, Losa R (2004) Crina® poultry and BMD alleviate the effects of a Clostridium perfringens challenge in commercial broilers. Poultry Science 83: 1787-1788.
-
Oviedo-Rondon EO, Hume ME, Hernandez C, Clemente-Hernandez A (2006) Intestinal microbial ecology of broilers vaccinated and challenged with mixed Eimeria species, and supplemented with essential oil blends. Poultry Science 85(5): 854-860.
-
Gong J, Yin F, Hou Y, Yin Y (2014) Review: Chinese herbs as alternatives to antibiotics in feed for swine and poultry production: Potential and challenges in application. Canadian J Animal Science 94(2): 223- 241.
-
Dhama K, Latheef SK, Mani S, Hari AS, Karthik K, et al. (2015) Multiple beneficial applications and modes of action of herbs in poultry health and production-a review. Int J of Pharmacology 11(3): 152-176.
-
Muthamilselvan T, Kuo T-F, Wu Y-C, Yang, W-C (2016) Herbal remedies for coccidiosis control: A review of plants, compounds and anticoccidial actions. Evidence Based Complementary and Alternative Medicine.
-
Khan SH, Iqbal J (2016) Recent advances in the role of organic acids in poultry nutrition. J Applied Animal Research 44: 359-369.
-
Broom LJ (2015) Organic acids for improving intestinal health of poultry. World’s Poultry Science Journal 71: 630-642.
-
Booyen F, Haesbrouck F, Vanparys A, Volf J, Mahu M, et al. (2008) Coated fatty acids alter virulence properties of Salmonella Typhimurium and decrease intestinal colonization of pigs. Veterinary Microbiology 132(3-4): 319-327.
-
Khodambashi, Emami N, Graystone EN, Broom LJ (2015) Organic acid based products promote broiler gut health and performance. Proc 2015 Poultry Science Assn.
-
Sun Y, O’Riordan MXD (2013) Regulation of bacterial pathogenesis by intestinal short-chain fatty acids. Advances in Appl Microbiology 85: 93-118.
-
Iba AM, Berchieri A (1995) Studies on the use of a formic acid-propionic acid mixture (Bio-add®) to control experimental Salmonella infection in broiler chickens. Avian Pathology 24: 303-311.
-
Nava GM, Attene-Ramos MS, Gaskins HR, Richards JD (2009) Molecular analysis of microbial community structure in the chicken ileum following organic acid supplementation. Vet Microbiology 137: 345-353.
-
Nakai SA, Siebert KJ (2003) Validation of bacterial growth inhibition models based on molecular properties of organic acids. Int J Food Microbiol 86(3): 249-255.
-
Ghazala AA, Atta AM, Elkloub K, Mustafa MEL, Shata RFH (2011) Effect of dietary supplementation of organic acids on performance, nutrients digestibility and health of broiler chicks. Int J Poultry Science 10: 176-184. Open Access Journal of Veterinary Science & Research
-
Emami NK, Naeini SZ, Ruiz-Feria (2013) Growth performance, digestibility, immune response and intestinal morphology of male broilers fed phosphorus deficient diets supplemented with microbial phytase and organic acids. Livestock Science 157(2-3): 506-513.
-
Karnezos TP, Dvorak R, Novak C (2016) Performance of broilers fed diets supplemented with medium chain fatty acids or medium chain fatty acids plus a probiotic and challenged with Clostridium perfringens. Proceedings of the 2016 International Poultry Symposium M-130.
-
Van Immerseel F, De Buck J, De Smet I, Pasmans F, Haesebrouck F, et al. (2004) Interactions of butyric acid –and acetic acid-treated Salmonella with chicken primary cecal epithelial cells in vitro. Avian Diseases 48: 384-391.
-
Ao T (2011) Using Exogenous Enzymes to Increase the Nutritional Value of Soybean Meal in Poultry Diet, Soybean and Nutrition. In. Hany El-Shemy (ed), In Tech.
-
Collett Stephen Dr (2013) Seed Feed Weed.
-
Connolly A (2010) A primacy in science and education-Part 3. Antibiotic-free animal production.
- The Digital Stethoscope: Harnessing AI in Veterinary Medicine Without Losing Our Healing Touch
- Meningoencephalomyelitis of Unknown Etiology: Short-Term Effect of Two Treatment Protocols on Cerebrospinal Fluid
- Safety and Efficacy of the HomeoPet Cough in Domestic Pets –A Clinical and Correction Analysis Based Upon User Response Survey
- Non Human Animals Responses to Social Loss
- Owner Reported Clinical Outcomes of a Homeopathic Proprietary Preparation for the Treatment of Upper Respiratory and Nasal Disorders in Companion Animals
- Effects and Diagnostic Approach of Ultrasound in Veterinary Practice: A Systematic Review