Intravenous Injection of Serratia marcescens Nuclease in Mice Induced Elevation of Synthetized and Secreted into the Serum Extracellular Ribonucleases
Deoxyribonucleases and ribonucleases are interest as potential anticancer compounds. Along with mammalian DNases and RNases the cancer suppressive effect was demonstrated for expressing the both activities nuclease from bacterium Serratia marcescens. The aim of the study was to examine how the appearing small amount of S.marcescens nuclease in blood can influence the level of serum DNase and RNase activities. Intravenous injection of S. marcescens nuclease in mice at the dose of 0.5μg /g of mouse was found to induce intensive rising RNA degrading activities in the serum. The induced activities differed from S.marcescens nuclease by the products of RNA degradation and requirement of magnesium cations. S.marcescens at the dose of 0.5μg /g of mouse did not both induce acute adverse effects and influenced on physical appearance and behavior in mice that evidenced a lack of the nuclease toxicity in vivo.
Introduction
The function of deoxyribonucleases and ribonucleases are interest as potential anticancer compounds due to their ability to degrade the released by tumor cells circulating extracellular DNA or RNA which presumably regulate the processes of tumor growth and metastasis [1, 2]. However, the mammalian RNases and DNases can be bound by the RNase or DNase inhibitors that may decrease their antitumor effectivity. Earlier the increased DNase activity was observed in mice serum resulted from the injection of nuclease that originated from Gram negative bacteria Serratia marcescens (further S. marcescens) [3]. The nuclease (EC 3.1.30.2) heads a family of homological nonspecific nucleases which are widely spread in the world [4]. S. marcescens nuclease is a well-studied enzyme [5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16]. Since the nuclease differs from mammalian DNases and RNases by the protein structure it cannot interact with mammalian RNase or DNase inhibitors. It is represented in two major isoforms, Sm1 and Sm2, differing by N-terminal three-peptide fragment and pIs [13]. The isoforms rapidly degrade both DNA and RNA in endonucleolytic manner and terminate the hydrolysis with producing di-, tri-, tetra-, penta – and hexanucleotides [10, 15]. Magnesium cations (further magnesium) activate these isoforms. In the presence of magnesium both isoforms degrade polyA, polyC and polyU. However, the rates of degradation are respectively 2-, 70- and 600-fold slower than the rate for RNA degradation. Both isoforms do not degrade PolyG [15]. In the presence of magnesium the ratio of enzymatic activity with RNA as substrate to that with DNA as substrate is constant and equal to 1.1-1.3 [10, 11]. In the absence of magnesium, the enzymatic activities diminish significantly, and the nuclease demonstrates its preference to DNA substrate in comparison with RNA substrate [8]. Due to the potent degrading activity toward both DNA and RNA the nuclease was studied as anti-rabies agent [17] and also as one that is similar to Pulmozyme® in their hydrolysis of DNA in sputum [18]. Earlier the nuclease was found to suppress the cancer growth. After intraperitoneal injection of the nuclease in to white mice the development of the intraperitoneal Ehrlich carcinoma slowed down by 50% [3]. Suppression of the carcinoma cells was also found after subcutaneous or intramuscular injection of the nuclease. For all modes of injection of the nuclease a rising DNase activity was observed in the serum for two hours after the injection. The increased DNase activity was referred to S.marcescens nuclease and explained by its filtration through the tissues barriers [3]. Nothing was reported of RNase activity in the serum as a result of the nuclease injection. Meanwhile, to test either the appearance of S.marcescens nuclease in the mammalian blood can influence the level of DNase and RNase activities in serum is interest and important. The aim of this study was to examine how the appearing in blood small amount of S.marcescens nuclease can influence the level of serum DNase and RNase activities.
Materials and Methods
All reagents were purchased from commercial sources, unless stated otherwise, and used without further purification. Cytochrome C, carbonic anhydrase and bovine serum albumin, poly A, poly C and highly polymerized DNA from Herring Testes (type XIV) were from Sigma, unspecified (total) RNA was from ICN.
Purification of S. marcescens nuclease was carried out as was described earlier [19]. The procedure was based on routine preparation of the fermentation medium and fractional salt precipitation of the crude nuclease, the dialysis, and cation exchange chromatography with NGC Discover chromatography system using UNO S12 column (Bio-Rad, USA). The DNA-degrading enzymatic activity in the collected fractions was tested by acid precipitation method as described further. The purity of the nuclease fractions was analyzed by denaturing sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) using 12% gel.
The purified nuclease was then dialyzed against distilled water and lyophilized. After that 1mg of lyophilized nuclease was dissolved with 0.2 ml of physiological saline, 0.85% water solution of NaCl, and then centrifuged at 4oC for removing the pellet. Concentration of the nuclease solution was calculated on the basis of absorption of the supernatant at 280 nm and the molar extinction coefficient of 47 292 M-1/ cm2 [20].
The nuclease activity was assayed by the method based on estimation of the amount of acid soluble low molecular nucleotide fragments produced by the nuclease [11] when after addition of the tested aliquot to 9-fold volume of the assay mixture the concentration of Tris-HCl buffer, pH 8.5, was 50 mM, MgSO4 - 6 mM (unless stated otherwise), and appropriate substrate (DNA or RNA or poly A or poly C) - 1 mg/ml of the incubation mixture. The incubation was performed at 37°C for 15 min – 5 h so that not more than 15-50% of the substrate had been converted to acid-soluble products. The hydrolysis was stopped with addition of chilled 4% perchloric acid (unless stated otherwise). The precipitate was removed by centrifugation. The absorption of supernatant was monitored at 260 nm. Experiments with animals were performed using unbreded white mice weighing 20g, per five mice in each experimental group. The mice of one group were injected with physiological saline instead the nuclease. Their serum was used as a control one. The other mice were injected with the nuclease solution, by 0.05 ml into the tail venous. Before the injection the prepared as described above nuclease solution was additionally two-fold diluted with the physiological saline. The serums were obtained immediately after the injection and two hours later the nuclease injection. For the serum preparation the blood was taken from the heart right after beginning of the mice euthanizing with carbon dioxide, however prior to cardiac arrest. One more group of the nuclease injected mice was used for determination of the short term toxicity. These mice were observed for 2 weeks after the nuclease injection and then were sacrificed and subjected to necropsy for the organ examination.
For serum preparation in sufficient quantity the taken blood in each group of mice were pooled together and serum isolation was carried out in accordance with the routine method. To form a clot the blood was incubated for 60 min at 25°C. Then the clot was removed and the blood cells were separated by 10 min centrifugation at 1300 g. Serum samples were stored at -20°C.
For analyzing the products of nucleic acids degradations RNA- or DNA were prior degraded at 37°C for respectively 15 or 30 min with the nuclease or the serum samples. The incubation mixture also contained 50 mM Tris-HCl buffer, pH 8.5. DNA degradation was carried out in the presence of 6 mM MgSO4. RNA degradation was carried out both in the presence and in the absence of magnesium. Before addition in the incubation mixture the tested materials were diluted with water so that the activity per 1 ml of the diluted solution was similar to the activity of respectively diluted nuclease. The nuclease was 10 000 –fold- or 2 500-fold diluted towards the substrate DNA or RNA respectively. The serum sample obtained right after the nuclease injection was 300-fold diluted toward the DNA substrate and 160- or 36-fold diluted towards the RNA substrate in the presence- or the absence of magnesium respectively. The serum sample obtained two hours later the nuclease injection was, respectively, 7- fold diluted towards the DNA- and RNA substrate in the presence of magnesium or 10-fold diluted towards the RNA substrate in the absence of magnesium. The control serum sample was not diluted towards the DNA- and RNA substrate in the presence of magnesium or 2.5- fold diluted towards the RNA substrate in the absence of magnesium. The degradation was stopped by freezing. The mixtures containing the products of degradations were analyzed with agarose gel electrophoresis when PCR Marker (Sigma) containing eight double- stranded DNA- fragments from 2000 - to 50 base pairs (bp) long was used as the polynucleotide Ladder.
Results and Discussion
Characterization of the Isolated Nuclease
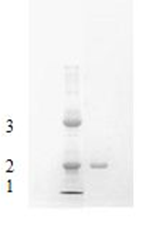
Using the earlier published protocol for purification of S. marcescens nuclease [19] we obtained the purified enzyme exhibiting high level of activity. Homogeneity of the used for biological examination nuclease sample was verified by SDS-PAGE that indicated a single protein band with an apparent molecular mass near 29 kDa (Figure 1). Earlier we demonstrated high purity of the isolated nuclease fraction with the full-scan MALDI-TOF mass spectrometry [19]. As the isoforms were similar in their key biochemical and biological properties [14, 20, 21] they were not separated from each other for further examination. The enzymatic activity of the dissolved nuclease fraction was 389333.3 U/ml per h with RNA as substrate and 294666.6 U/ml per h with DNA. So in the presence of magnesium the ratio of enzymatic activity with RNA as substrate to that with DNA as substrate was 1.3 that well correlated with previously published results [10, 11]. The nuclease concentration before the injection was 0.21 mg/ml.
Examination of Short -Term Effect of the Nuclease
Since the nuclease solution at the concentration of 0.2–25 µg/ml was known to express a weak cytotoxic effect towards malignant and non-malignant cells [21] as well as intramuscular, intraperitoneal, intravenous injection of 0.2 µg of the nuclease per 1 g of mouse did not cause any visible changes in the animals’ behavior or in tissue morphology at the places of the nuclease injection [17] the dose of 10.5 µg of the nuclease per mouse ( about 0.5 µg per 1 g of mouse) was selected for the examination. The chosen dose was supposedly harmless and close to the previously used one for the study of suppression of the cancer growth (0.25µg /g of mouse) [3]. However, in order to avoid serious toxicological effects in mice after the intravenous nuclease injection and also to obtain information regarding the dose that should be used further, determination of short-term adverse effects of the administered in a single dose nuclease was carried out. For two weeks the once injected and non-injected mice were monitored daily for any changes in physical appearance and behavior that suddenly might occur due to the unknown long-term toxic effects of the nuclease. No deaths occurred right after – or two hours later intravenous injection of the nuclease at the dose of 0.5µg/g of mouse. No changes were observed in motor function, such as disturbances of gait or muscle tone, and level of arousal such as hyperactivity and lethargy, or autonomic functions, such as salivation, lacrimation, urination, and defecation, or psychological status, likewise aggression, biting, self- mutilation.
No mice showed significant behavioral changes such as changes in motor function, level of arousal, autonomic functions, and psychological status that might indicate the neurotoxicity effect. Physical appearance, such as skin, fur, and eyes was normal. Food and water consumption remained normal. The emaciation was no found at the end of observation. The lack of effect on appetite means that there was no disruption of carbohydrate, protein or fat metabolism levels. During fourteen days of observation the mice weight was similar compared to the control. No mortality or physiological and behavioral changes were observed in the mice during the time of observation. Additionally, no gross pathological findings were observed. No histological changes were observed in the liver, kidney, spleen, thyroid or other organs in mice which were exposed to the nuclease injection.
Thus intravenous injection of the nuclease at the dose of 0.5µg /g of mouse did not induce acute adverse effects on physical appearance and behavior in mice that showed a lack of the nuclease toxicity in vivo at the selected dose. This result agreed with previously accrued information of the nuclease harmlessness at the dose of 0.25µg /g of mouse [3] and the absence of injurious effect of the nuclease in vivo as a result of its intracerebral, subcutaneous, intramuscular, intraperitoneal and intravenous injection in mice [17].
Evaluation of DNase and RNase Activity in the Serum Samples before and after the Nuclease Injection into the Mice
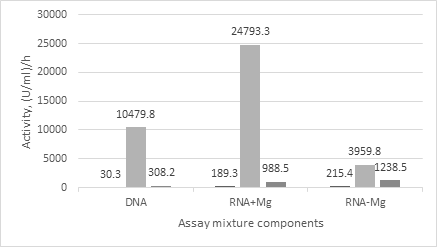
Both activities, as DNase and RNase, were found in the control serum (Figure 2). Interestingly that RNase activity was similar as in the presence and in the absence of magnesium, and was about 5-fold higher than the DNase activity.
After intravenous injection of the nuclease both activities in the serum sharply increased, by 350 –fold towards DNA and – 130- fold towards RNA in the presence of magnesium. The ratio of enzymatic activities in serum towards RNA and DNA substrates in the presence of magnesium was 2.4 that is higher than the ratio in the nuclease itself. In the absence of magnesium RNase activity also increased compared the control serum (by about 18-fold) but was lower than the activity in the presence of magnesium. Two hours later the nuclease injection, both RNase and DNase activities in the serum reduced, in the presence of magnesium respectively by 25- and 30-times, in the absence – by about 3- fold. The RNase activity in the absence of magnesium became close to the activity in the presence one. At the same time, they stayed by about 5-10 –fold higher than the activities in the control serum. The ratio of RNase and DNase activities in the resulted serum was higher than 3.0.
Thus comparative analysis of DNase and RNase activities in the serum samples showed that before the nuclease injection the level of own, secreted in blood enzymes for degradation of both DNA and RNA is rather low. After the nuclease injection the level of DNase and RNase activities in the serums increased many times that was expected and clear, because the nuclease displayed the both activities by itself. However significant degradation of RNA by the serum samples in the absence of magnesium and inequality in the ratios of RNase to DNase activities at the serum samples compared to the nuclease itself allowed us to suspect that the nuclease injection in mice caused uprising the own murine enzymes, notably the serum RNases.
In order to check the idea, at first we analyzed the products of RNA- or DNA degradations with both the nuclease as well the serum samples by agarose gel electrophoresis. Then we comparatively analyzed the rates of poly A and poly C degradations by the serum samples because the secreted murine RNases are known to differ both from each other and from S.marcescens nuclease by their preference to purine and pyrimidine substrates [15].
Analysis of the Products of RNA- or DNA Degradations by the Nuclease and the Serum Samples
Analyzing the products of DNA and RNA degradations (Figures 3a-c) by the serum samples (lines 8,9, 11,12, 14,15) and the nuclease (lines 5,6) we applied the tested solutions to the gel without dilution (lines 5, 8, 11, 14) and diluted 10 times (lines 6, 9, 12, 15) in order to obtain a more accurate picture. As controls we used the substrates at the beginning (line 2) and at the end (line 3) of incubation at 37o C as well the serum samples or the nuclease (lines 7,10,13 or 4), applying them to the gel with the same dilution and in
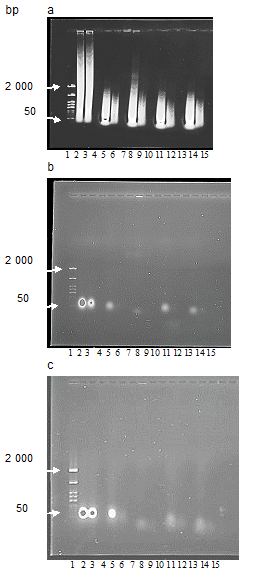
the same amounts as used for the degradation of DNA and RNA. As was seen except for the control serum neither other serum samples nor the nuclease solution displayed any polynucleotide contamination when were checked in the same amounts as used for the degradation of DNA and RNA. In addition, as the view of substrate samples at the beginning (line 2) and at the end (line 3) of incubation at 37°C was similar we conclude that the incubation of DNA or RNA at 37°C by itself did not changed the substrate composition.
The DNA substrate Figure 3 a lines 2,3 was represented with polynucleotide fragments of varying size. Degradation of DNA with the nuclease or the serum samples resulted in disappearing polynucleotide fragments which were much longer than 2 000 base pairs (lines 5,6, 8, 9, 11, 12, 14, 15), and in accumulating the products of shorter than 50 base pairs which were lack in the initial DNA substrate (lines 2, 3). However, when DNA substrate was incubated with the nuclease (lines 5,6) or the serums of the nuclease injected mice (lines11,12,14,15) the longest polynucleotide products were placed close to the marker’s band of 2000 base pairs.
But when DNA substrate was incubated with the control serum (lines 8,9) the largest polynucleotide products were longer than 2 000 base pairs. Degradation of RNA with the nuclease or the serum samples in the presence of magnesium were mainly displayed by the quenching luminescence Fig.3 b, lines 5, 8, 11, 14) if to compare with the RNA substrate as in the beginning (line 2) and in the end (line 3) of incubation. However, the products resulted from incubation with the control serum (line 8) were located lower than the appropriate zone of the other tested materials and differed from others by rather weak luminescence. Whereas the products of degradation by the nuclease (line 5) and serums of the nuclease injected mice (lines 11, 14) were located close to the spots of RNA substrate (lines 1, 2). In the absence of magnesium, the picture of products resulted from RNA degradation by the tested materials (Figure 3c) significantly differed from that obtained in the presence of magnesium. Firstly, the products of RNA degradation by the nuclease (line 5) distinguished from other ones by bright luminescence which was like to intensity of undegraded RNA (lines 2,3). As RNase activity in S.marcescens nuclease is known to be halted in the absence of magnesium [8] the bright luminescence of the RNA substrate (Fig.3 c line 5) after its incubation with the nuclease in the absence of magnesium connected with the lack of RNA degradation. Secondly, other tested materials resulted from degradation by both the control serum (line 8) and serums of the nuclease injected mice (lines 11, 14) were located lower than the undegraded RNA (lines 2, 3) or the products of RNA incubation with the nuclease (line 5). Their luminescence was rather weak and dispersed compared the undegraded RNA (lines 2, 3, 5) and also the analogical samples resulted from incubation in the presence of magnesium (Fig.3 b, lines 11,14). Besides that, localization of the products of RNA degradation by the control serum was similar independently on the presence of magnesium in the assay mixture (Fig.3 b and c, line 8) that was another difference between the control serum and the serums of the nuclease injected mice (Fig.3 b and c, lines 11, 14).
Analysis of Poly A or Poly C Degradations by the Serum Samples
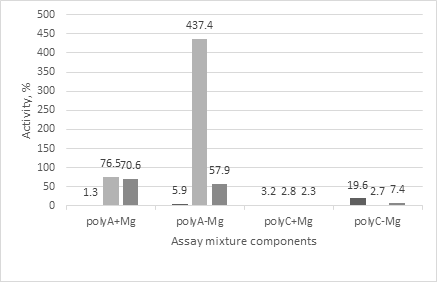
Comparative analysis of degradation of poly A and poly C with the serums showed (Figure 4) that only degradation of poly C in the absence of magnesium by the control serum was relatively high. Nevertheless, the rate of such degradation corresponded to 19.6% of the rate of RNA degradation, taken as 100 %. The nuclease injection was followed with the increasing rates of both RNA and poly C degradations. However, as in the presence and in the absence of magnesium the rates of poly C degradation were not higher than 7% of the RNA ones. Degradation of poly A with the control serum looked rather weak independently on the presence of magnesium. After the nuclease injection it has more than 10 times increased. However, only once the rate of poly A degradation was significantly higher than of RNA one, right after the nuclease injection and in the absence of magnesium in the assay mixture. For the other variants it was lower than the rate of RNA degradation (57. 9 – 76. 5 % of the rate in RNA) but approximately 1.5 –fold higher than the possible rate of poly A degradation with S. marcescens nuclease [15].
In this way the comparative analysis of degradation of polynucleotides by the serum samples as well the nuclease showed that all tested serum samples contained the murine own nucleases. At that, as after the nuclease injection the products of DNA degradation by the serum samples differed from the control one and better resembled the products resulted from the degradation with S. marcescens nuclease, we assume that the increased DNase activity in the serum right after the nuclease injection and two hours later mainly conditioned by the presence in the blood of the nuclease itself.
And in the opposite we believe that the portion of own mice RNases in the RNase activity of the serum samples from the nuclease injected mice is significant. So, as in S.marcescens nuclease the ratio of RNase to DNase activity has to be 1.1- 1.3 [10, 11] the portion of the nuclease itself in the RNase activity of serum obtained after the nuclease injection can be calculated. The RNase activity attributed to S.marcescens nuclease should be not more than 15000 U/ml per h right after the nuclease injection or not more than 400 U/ml per h two hours later. In reality it was much higher (Figure 2). So much, that it could not simply be a sum of RNase activities of the nuclease itself and the control serum. This means for us that some of the own mice RNases appeared in the serum after the nuclease injection, i.e. upon its influence and were secreted into the blood from the tissues.
Such secreted RNases could be RNase 2 and RNase 3 which belong to mammalian RNase A superfamily and are stored in the eosinophil secondary granules [22]. As known RNase3 release is induced in response to the interaction of allergens which are often represented by proteins. Like RNase 3 in mice and RNase 2 in Mammalia [23] the serum sample resulted from the nuclease injection demonstrated the requirement of magnesium for the catalysis that correlated with our results. As we found, the rate of RNA degradation in the absence of magnesium approximately 6-fold lower than the rate in the presence one (Figure 2). Also the serum sample resulted from the nuclease injection expressed the high rate of poly A degradation. The rates of polyA degradation by the serum samples were much higher, more than 1.5-fold-, than it could be due to the presence of nuclease itself [15] and approximately 70-fold higher compared the control serum (Figure 4). This allowed us to conclude that some of the presumably expressed RNases could rapidly degrade poly A. For example, RNase 2 is known to degrade poly A successfully. Unfortunately, murine RNases are characterized worse than human ones. At present almost all canonical and non- canonical RNases of mammalian RNase A superfamily are known in mice [22, 24, 25, 26, 27]. However, in mice and rats a striking diversity of RNases 2 and 3 counterparts is observed; up to 13 new eosinophil murine members were identified [28]. This diversity and lack of information on murine RNases make it difficult to reach a definitive conclusion. Moreover, our data is also not enough too to do final conclusions. The obtained results appeared to be unexpected for us. Nevertheless on the basis of above presented information one can conclude that intravenous injection of S.marcescens nuclease in mice induced the instant releasing the own mice RNases into blood. And the instant secreting the own mamalian RNases into blood serum upon the action of S.marcescens nuclease could represent a novel therapeutic approach in particular to suppression of malignancy.
Funding
The paper has been supported by the Kazan Federal University Strategic Academic Leadership Program (PRIORITY-2030).
Conflict of Interest
The authors declare no competing interests. The submitted work was carried out in the absence of any personal, professional or financial relationships that could potentially be construed as a conflict of interest.
Submission
The authors declare that the article is not submitted elsewhere and has not been published anywhere.
Research Involving Humans and Animals
The experimental protocol was designed according to the guidelines of the current European Directive (2010/63/ EU) on the care and protection of animals used for scientific purposes and approved by the Ethical Committee of the Kazan Federal University (Russian Federation, Kazan, accession number 459/06/04/2020).
References
-
Alekseeva L, Sen’kova A, Zenkova M, Mironova N (2020) Targeting Circulating Sines and Lines with DNase 1 provides Metastases Inhibition in Experimental Tumor Models. Mol Ther Nucleic Acids 20: 50-61.
-
Garcia-Olmo D, Garcia-Olmo DC, Dominguez-Berzosa C, Guadalajara H, Vega L, et al. (2012) Oncogenic Transformation Induced by Cell-Free Nucleic Acids Circulating in Plasma (Genometastasis) remains after the Surgical Resection of the Primary Tumor: A Pilot Study. Expert Opin Biol Ther 12(Suppl1): S61-S68.
-
Gabdullina GK (1980) The Effect of Serratia marcescens Nuclease on Cells and The Growth of Erlich Ascites Tumor. Dissertation, Institute of Oncology Problems by R.E.Kavetskii, Academy of Science of Ukrainian SSR.
-
Chen C, Krause K, Pettitt B (2009) Advantage of Being a Dimer for Serratia marcescens Endonuclease. J Phys Chem B 113(2): 511-521.
-
Benedik MJ, Strych U (1998) Serratia marcescens and its Extracellular Nuclease. FEMS Microbiology Letters 165(1): 1-13.
-
Trifonova E, Saveleva A, Romanova A (2015) Transgenic Expression of Serratia marcescens Native and Mutant Nucleases Modulates Tobacco Mosaic Virus Resistance in Nicotiana Tabacum L. Russian Journal of Genetics 51: 715-719.
-
Miller M, Tanner J, Alpaugh M, Benedik M, Krause K (1994) 2.1 A Structure of Serratia Endonuclease Suggests a Mechanism for Binding to Double-Stranded DNA. J Nature Struct Biol 1(7): 461-468.
-
Romanova J, Gubskaya V, Nuretdinov I, Zainutdinova E, Filimonova M (2017) Analysis of the Mechanism of Mg2+ Action on the RNase Activity of Serratia marcescens. Bio Nano Science 7(2): 276-283.
-
Friedhoff P, Kolmes B, Gimadutdinow O, Wende W, Krause KL, et al. (1996) Analysis of the Mechanism of the Serratia Nuclease Using Site-directed Mutagenesis. Nucleic Acids Res 24(14): 2632-2639.
-
Nestle M, Roberts WK (1969) An Extracellular Nuclease from Serratia marcescens. I. Purification and Some Properties of the Enzyme. J Biol Chem 244(19): 5213- 5218.
-
Leshchinskaya I, Balaban N, Egorova G (1974) Isolation and Characterization of Highly Purified Preparation of Nuclease from Serratia marcescens. Biochemistry, Moscow 39: 95-100.
-
Biedermann K, Jepsen P, Riise E, Svendsen I (1989) Purification and Characterization of a Serratia marcescens Nuclease Produced by Escherichia Coli. Carlsberg Res Commun 54: 17-27.
-
Pedersen J, Filimonova M, Roepstorff P, Biedermann K (1993) Characterization of Serratia marcescens Nuclease Isoforms by Plasma Desorption Mass Spectrometry. Biochim Biophys Acta 120291): 13-21.
-
Filimonova MN, Gubskaya VP, Nuretdinov IA, Benedik MJ, Bogomolnaya LM, et al. (1997) Isoforms of Serratia marcescens Nuclease: The Role of Mg2+ in the Hydrolysis Mechanism. Biochemistry 62(9): 983-988.
-
Filimonova MN, Garusov AV, Smetanina TA, Andreeva MA (1996) Isoforms of Serratia marcescens Nuclease. Comparative Analysis of the Substrate Specificity. Biochemistry 61(10): 1274-1278.
-
Romanova J, Filimonova M (2012) The Effects of Addition of Mononucleotides on Sma Nuc Endonuclease Activity. Scientific World Journal 2012: 454176.
-
Khismatullina N, Ivanov A, Martynova E, Mironov A, Zainutdinova E, et al. (2014) Nuclease Composition with Anti-Rabies Activity. J Pure Appl Microbiol 8(2): 499- 504.
-
Vafina G, Zainutdinova E, Bulatov E, Filimonova MN (2018) Endonuclease from Gram-Negative Bacteria Serratia marcescens Is as Effective as Pulmozyme in the Hydrolysis of DNA in Sputum. Front Pharmacol 9: 1-8.
-
Vafina G, Bulatov E, Zainutdinova E, Filimonova M (2016) A One-Step Protocol for Chromatographic Purification of Non-Recombinant Exogenous Bacterial Enzyme: Nuclease of Serratia marcescens. Bio Nano Science 6: 335-337.
-
Filimonova M, Baratova L, Vospel’nikova N, Zheltova A, Leshchinskaya I (1981) Endonuclease Serratia marcescens. Enzyme Characteristic. Biokhimiya 46: 1660-1666.
-
Malanyeva A, Salikhova T, Rizvanov A, Filimonova M (2020) The Nuclease from Gram-Negative Bacteria Serratia marcescens is Weakly Cytotoxic at Therapeutic Doses. Open Access Journal of Veterinary Science & Research 5: 1-8.
-
Martin L, Koczera P, Simons N, Zechendorf E, Hoeger J, et al. (2016) The Human Host Defense Ribonucleases 1, 3 and 7 Are Elevated in Patients with Sepsis after Major Surgery—A Pilot Study. Int J Mol Sci 17(3): 294-299.
-
Bollenbach T, Schuster G, Stern D (2004) Cooperation of endo- and exoribonucleases in chloroplast mRNA turnover. Prog Nucleic Acid Res Mol Biol 78: 305-337.
-
Kanwar S, Mishra P, Meena KR, Gupta S, Kumar R (2016) Ribonucleases and their Applications. Journal of Advanced Biotechnology and Bioengineering 4: 1-10.
-
Samuelson L, Wiebauer K, Howard G, Schmid RM, Koeplin D, et al. (1991) Isolation of the murine ribonuclease gene Rib-1: structure and tissue specific expression in pancreas and parotid gland Nucleic Acids Res 19(24): 6935-6941.
-
Sporn M, Lazarus H, Smith J, Henderson W (1969) Studies on nuclear exoribonucleases.3. Isolation and properties of the enzyme from normal and malignant tissues of the mouse. Biochemistry 8(4): 1698-1706.
-
Cho S, Beintema J, Zhang J (2005) The ribonuclease A superfamily of mammals and birds: identifying new members and tracing evolutionary histories. Genomics 85(2): 208-220.
-
Lu L, Li J, Moussaoui M, Boix E (2018) Immune Modulation by Human Secreted RNases at the Extracellular Space. Frontiers in Immunology 16(9): 1012.
- The Digital Stethoscope: Harnessing AI in Veterinary Medicine Without Losing Our Healing Touch
- Meningoencephalomyelitis of Unknown Etiology: Short-Term Effect of Two Treatment Protocols on Cerebrospinal Fluid
- Safety and Efficacy of the HomeoPet Cough in Domestic Pets –A Clinical and Correction Analysis Based Upon User Response Survey
- Non Human Animals Responses to Social Loss
- Owner Reported Clinical Outcomes of a Homeopathic Proprietary Preparation for the Treatment of Upper Respiratory and Nasal Disorders in Companion Animals
- Effects and Diagnostic Approach of Ultrasound in Veterinary Practice: A Systematic Review