Interactions that are Important to Our Practice
Introduction: Our body hosts a complex microbial community [microbiota] responsible for its development, optimal functioning and health. We are constantly witnessing the emergence of new diseases caused by a large number of highly pathogenic viruses. Viral diseases are one of the most important predictors of subsequent bacterial infection. At the same time, we are seeing a worldwide increase in antibiotic use. Despite the fact that in the prevailing percentage terms it has no visible adverse effect, the alteration of the human microbiota can have serious short-term or, worse, long-term consequences for our health. The aim of the review is to trace the relationships between pathogenic viruses, bacteria and the human microbiota. Conclusion: Cocirculation of respiratory pathogens can lead to competitive or cooperative forms of interaction between them. Understanding how our interventions do not cause disruption of the balance of the human microbiota - reduced species diversity, altered metabolic activity, stimulation of selection and sharing of genes responsible for AMR will allow us to adjust strategies to treat infectious diseases. Knowing the interactions between commensal and opportunistic species will help us determine the time when we need to switch from non-pharmacological to pharmacological interventions, preserving the balance of the microbiota and that between it and the macroorganism, as well as reducing the rate of relapses. The right combination of pharmacological, non-pharmacological and dietary interventions in the personalized treatment of patients will allow us to reduce the unnecessary use of antibiotics and better control the risk in the course of diseases.
Introduction
The last decades have seen the emergence of new diseases caused by a large number of highly pathogenic viruses. Viral diseases are one of the most important predictors of subsequent bacterial infection. These infections can alter the structure of the respiratory microbiome increasing the number of pathogenic bacteria. Viral-bacterial interactions play a critical role in the pathogenesis of bacterial infection. At the same time, we are seeing an increasing rate of antibiotic resistance. The dilemma before us is whether we are on the cusp of a shift in treatment strategies for infectious diseases and whether harnessing the ability of the microbiota to influence host immunity will become imperative in our therapeutic strategies for infectious as well as other diseases.
Microbiota
Our body hosts a complex and dynamic microbial community (microbiota), responsible for its development, optimal functioning and health. It performs a barrier function on the epidermis, mucous membranes of the gastrointestinal tract, respiratory and urogenital systems. Its composition is not constant throughout our life and begins to form immediately after our birth. The most dynamic period is during the first three years. It is directly dependent on the type of birth, the environment, the season, socio-economic living conditions, nutrition, vaccinations, past illnesses, harmful habits, taking antibiotics, etc.
The microbiota is a collection of ecological niches with different composition and population density of microorganisms-bacteria, archaea, fungi and viruses and performs the following functions-trophic, metabolic and protective [1]. The metabolic activity of the intestinal microbiota forms an anaerobic environment that suppresses the virulence of the pathogen, the protective-production of bacteriocins that inhibit the growth or survival of the pathogen and the trophic - mobilization of bacteriophages that attack specific bacterial strains, minimizing their impact on the commensal microbiota. The initial categorization of the human microbiota was completed in 2012 [2]. Of the composition, the group of bacteria is best but still not sufficiently studied, but the study of the functions of fungi and viruses is also progressing [3, 4]. They are transitory and permanent. The vast majority of bacteria are non-pathogenic colonizers (commensals) that live symbiotically with the macroorganism and mutually provide conditions for their optimal condition. Their task is to suppress the colonization of opportunistic pathogens. To the former belong the opportunistic pathogens responsible for infectious diseases and a large part of the probiotics sold. They enter from the environment and do not undergo permanent colonization due to lack of suitable receptors or inability to compete for space. This phenomenon has been well studied in the gastrointestinal tract. In most cases, pathogens attempting to colonize mucosal surfaces elicit a strong immune response aimed at clearing the pathogen. The other part of the bacteria involved in the microbiota are the permanent members. They colonize the host in waves during ontogenesis and through various adaptation mechanisms enter into permanent relations with it. An example of this is Streptococcus saguinis, which is one of the first colonizers of the oral cavity - a gram-positive, non-spore-forming facultative anaerobe. The average time for its colonization is 9 months [5]. While the primary colonization can last for months, subsequent waves are shortened and in adult individuals can reach 2-4 weeks. These are the “true” commensal bacteria. They can be assumed to have the property of “memory”. The nasopharynx was thought to be a reservoir for sinus infections because conventional examination found a >90% concordance between pathogens of the osteomeatal complex and the nasopharynx. More recent gene sequencing of the microbiota from patients after functional endonasal sinus surgery, however, found a microbiome similar to the anterior nose, which temporarily became similar to that of the nasopharynx after surgery and then returned to the original niche equilibrium about 6 weeks [6].
The commensals stimulate various immune cells in the mucous membranes and help the maturation of gut- associated lymphoid tissues [7]. In the sequencing of the microbiota, commensal species with an immunomodulatory effect on the host have been identified. It is specific to individual groups of bacteria and is generally influenced by the species diversity of the microbiota, or alpha diversity. From the permanently resident types of bacteria, those that have an immunomodulating effect are called autobionts, and those that can cause diseases under certain conditions are called pathobionts. They are permanent members, but in limited populations. In eubiosis, auto- and pathobionts are in perfect balance with the host. With a normally functioning microbiota, they do not cause disease, but when the composition and/or is impaired, they can cause diseases of varying severity and even death. Microbes constantly move through the commensal-pathogen continuum depending on circumstances and co-infections. Pneumococcus, regardless of the fact that it is considered the main cause of pneumonia, in eubiosis is a member of the healthy nasopharynx, as well as Stapylococcus aureus and other microorganisms, and in dysbiosis normal commensals can cause severe diseases in immunocompromised [8, 9]. Autobionts are part of the normal microbiota that have evolved a way to colonize humans, actively participating in host immune regulation and health maintenance. With them, there is still much to learn about the mechanisms that provoke an immunomodulatory effect [10].
The significance of the microbiota in the course of the immune responses of the host is in the influence and in the maturation and functioning of different types of immune cells - IgA secreting plasma cells, Th17, Treg lymphocytes, invariant natural killer T cells (iNKT), NK cells, macrophages, dendritic cells (DC), etc. [11]. The immune effects controlled by the microbiota also play an autoregulatory role on itself. Induced by commensal bacteria, IgA secreting plasma cells are involved in the control of their number and composition [12, 13]. The gut microbiota has been found to control the immune status in an effector or regulatory direction through various stimuli [14]. Resident commensals on the skin provoke modulatory effects and induce local Th17 and Th1 responses to protect against bacterial infections [15]. Host genotype also plays an important role. In mouse models, it has been found that harmless commensals can become pathogenic [16].
The human microbiota is an indivisible whole, but in view of our area of interest, I will focus mainly on ecological niches in the upper respiratory tract (URT). The microbiota of the upper respiratory tract in healthy individuals consists of several genera. During the first year of life, the genera Staphylococcus, Streptococcus, Corynebacterium, Moraxella, Haemophilus and Alloicoccus/Dolosigranulum predominate, due to contact with the mother’s skin and milk, and the vaginal mucosa. After the third year, the most common are: Staphylococcus, Streptococcus, Corynebacterium, Prevotella, Veillonella, Propionibacterium and Fusobacterium. The commensals inhabiting the URT mainly inhabit the nasal cavity, naso- and oropharynx. Corynebacterium, Dolosigranulum, Streptococcus epidermidis and Streptococcus lugdunensis are thought to be primarily responsible for suppressing the possibility of disease caused by Streptococcus pneumoniae, Haemophilus influenzae, Staphylococcus aureus, Moraxella catharralis [17, 18, 19, 20, 21, 22]. About unlike URT, the microbiota of the gastrointestinal tract is dominated by two main genera, evolutionaly adapted to this niche. These are the gram- negative Bacteroides and the gram-positive Firmicutes [23]. Studies have shown that gut and commensal bacteria of the gastrointestinal tract participate in the activation of shared immune pathways regulating resistance to pulmonary infections [24, 25]. The upper and lower respiratory tract contain different ecological niches. URT are schematically divided into: front nose, paranasal sinuses, naso- and oropharynx and larynx [26]. They are characterized by the combination of taxonomically rich niches such as nose, naso- and oropharynx and scarce niches such as middle ear, sinuses and lower respiratory tract [27]. The nasal, nasopharyngeal and oropharyngeal microbiota consist of different types of aerobic and anaerobic microorganisms, the latter being predominant. Loss of the oral-nasopharyngeal distinction usually precedes respiratory tract infections [28]. Viral- bacterial interactions play a critical role in the pathogenesis of bacterial infection. When the microbial community becomes unbalanced, opportunistic pathogens can spread from the nose and nasopharynx to microbially poorer niches and cause disease. Immune regulation against viruses is modulated primarily by the gut microbiota [29].
Of the known respiratory viruses, those that most often attack our respiratory tract are: influenza virus, parainfluenza virus, RSV, adenoviruses, measles virus, rhinoviruses and coronaviruses [30]. The most frequently detected bacterial pathogens in the respiratory tract are: S. pneumoniae, S. pyogenes, H. influenzae, S. aureus, Neisseria meningitidis, Mycobacterium tuberculosis, Bordetella pertussis, M. catharralis, and in immunocompromised patients, Pseudomonas aeruginosa [31].
The barrier function of the microbiota in the nose and nasopharynx is expressed in а competition between commensal and potentially pathogenic species of bacteria, ensuring the regulation of pathogenic species, host responses and protection against infections. In eubiosis, commensal bacteria suppress the colonization of opportunistic pathogens by regulating the mucosal barrier and inhibit pathogen exposure through direct competition and indirect immune modulation. The mechanisms used are:
- Induction of IFN-λ secretion from the nasal mucosa. The commensal Staphylococcus epidermidis enhances resistance to influenza A virus infection in mice [32] and intestinal commensals modulate resistance to infection of the lung epithelium with the same virus [33].
- Secretion of antimicrobial peptide [AMP], bacteriocins and proinflammatory cytokines from the nasal mucosa. The commensal strain S. epidermidis blocks acquisition and limits the growth of S. aureus and M. catharralis in an AMP-dependent manner in mice [34, 35], and Haemophilus haemolyticus of H. influenzae [36].
- Influence on the adaptive immune response and generation of immune memory. The commensal Streptococcus mitis induced a cross-reactive immunity [antibodies and IL-17] to S. pneumoniae in mice [37]. The same was observed with Neisseria lactamica and Neisseria meningitidis.
- Production of antibiotics. The nasal commensal Staphylococcus lugdunensis produces lugdunin, an antibiotic with bactericidal activity against S. aureus and S. pneumoniae. Carriage of S. lugdunensis correlates with reduced S. aureus microbial counts [38].
- Inhibition of binding to the mucous membrane. Streptococcus salivarius limits the binding of S. pneumoniae independently of bacteriocin production [39]. The genus Corynebacterium directly competes with pathogens of the upper respiratory tract. Corynebacterium assolens inhibits the growth of S. pneumoniae via free fatty acids, and the cell-free environment of Corynebacterium striatum limits the adhesion of S. aureus [40, 41].
- Possibility of commensals of the genus Streptococcus to destroy formed biofilms (hypothesis) [42].
• For our practice, the study and understanding of virus-virus, virus-bacterial, virus-microbiota and interbacterial interactions is extremely important.
Bacteria exist not only as independent planktonic forms, but also in organized ecosystems [biofilm] within the human organism. The second group is more stable. Antoni van Leeuwenhoek (1632-1723) developed the concept of microorganisms that bind to the surface in the form of dental plaque. Characklis’ research on microbial slime taken from bodies of water sheds further light on the issue [43]. Up to 99% of the bacteria in the human body are thought to exist in the form of a bacterial biofilm [44]. It forms after individual planktonic forms merge and attach to the various surfaces through glucoconjugate bonds [45] then form an exopolysaccharide matrix [46]. By means of the towers and channels in the interior, waste substances are eliminated and the pH gradient is maintained. The oxygen tension gradient determines the increased metabolic activity at the surface of the matrix and the reduced/dormancy in the deeper layers. The cells in the biofilm are in constant contact with each other by means of chemical signaling [quorum sensing]. As the biofilm matures, individual cells are detached and directed to colonize another surface [47]. The bacterial biofilm can contain several strains of bacteria. It is characterized by its high degree of resistance to antibiotics and the host’s immune mechanisms-low susceptibility to opsonization and phagocytosis [48]. It is still being clarified whether biofilms can be considered as a driver of respiratory diseases [49].
Virome
The virome is a poorly understood component of the microbiota. Viral load carriage in healthy controls suggests a benign carriage, similar to commensal bacteria [50]. We have yet to determine the reasons why “healthy carriage” is replaced by a dominance of viral pathogens. Metagenomic analysis, in contrast to PCR-based analysis, showed the presence of many more viral sequences in children with unexplained fever. The virome is a poorly understood component of the microbiota. Viral load carriage in healthy controls suggests a benign carriage, similar to commensal bacteria [50]. We have yet to determine the reasons why “healthy carriage” is replaced by a dominance of viral pathogens. Metagenomic analysis, in contrast to PCR-based analysis, showed the presence of many more viral sequences in children with unexplained fever. This suggests us that the virome is a common cause of upper respiratory tract disease. During the same annual period, several respiratory viruses can circulate simultaneously. Co-circulation of viral pathogens can lead to multi-epidemics such as the combination of COVID-19, RSV and influenza, or the past viral infection can induce a refractory period during which the host is unlikely to be infected by another respiratory virus. To understand the interactions (interference), it is important to determine whether the viruses share the same environmental conditions (seasonality).
Interviral Interactions
Interviral interactions determine the individual risks of infection and the dynamics of the course and. The factors that would direct us to viral interference are:
- The ability of the interfering virus to induce a rapid IFN response due to the expression of interferon (IFN) stimulating genes (ISG) type I (IFN-α/β) and type III (IFN-λ) and provided temporary non-specific immunity to the host. Viral defense is expressed in the production of effectors that directly inhibit viral replication, as well as chemokines and cytokines [51, 52].
- The degree of sensitivity of the second virus to immune mediators
- The extent to which different viruses counteract the induction and antiviral effects of IFN and
- The pattern of innate immune response induced by the virus in the upper and lower respiratory tracts [31].
Viruses can simultaneously or sequentially infect the respiratory tract. Depending on whether the infection of the first virus will enhance or weaken the infection and replication of the second virus, we observe a positive (synergistic) or negative (antagonistic) interaction. The positive interaction is observed in:
- SARS-CoV-2, RSV and pandemic influenza virus A (pH1N1)[53]
- PIV1 and PIV2
- RSV and HMPV
- The surface antigen of the first virus serves as a receptor for the second (HbSAg of HBV for HDV). When blocking and/or reducing cell surface receptors and competition for cellular resources is severe, this is a possible option for evasion by other respiratory viruses.
Co-infection increases disease severity through excessive production of IFN and pro-inflammatory cytokines or through decreased secretion of non-inflammatory mediators such as interleukin (IL) 10 [54]. Knowledge of negative virus-virus interactions are much more important for practice. They are homologous and heterologous depending on whether the viruses belong to one or two different families. Blockade and/or reduction of cell surface receptors and competition for cellular resources are considered mechanisms of negative virus-virus interaction. Studying interference species in animal models has a potential limitation because their immune response to human respiratory viruses and immune evasion mechanisms in most cases differ.
1. In the homologous interaction, presumably cross-
reactive immunity against the first virus prevents infection from the second virus
- Hierarchical model-pH1N1, H1N1 and H3N2.
- Time pattern-RSV, HMPV and PIV-taxonomically belong to the same family. 2. B. In the heterologous type of interference, the provoking of the non-specific immune response by the first virus reduces or prevents the infection and replication of the second virus:
- Coinfection with IAV in MDCK cells inhibits RSV replication by removing sialic acid from the cell surface and competing for viral protein synthesis [55, 56, 57].
- Live enterovirus vaccines (LEV) are used to prevent pathogenic enteroviruses in children.
- Oral administration of LEV in children reduces the detection of some unrelated respiratory viruses— influenza, PIV, RSV, HRV, and AdV [58, 59].
- Prior infection with IАV (H1N1) or IАV (H3N2) prevents subsequent infection with retroviruses.
- IBV and RSV [60].
- RSV c reduced HRV infection rate.
- HRV reduces SARS-CoV-2 replication.
- HRV reduced probability of co-detection IAV.
- RSV reduced probability of HMPV co-detection.
- IBV and Adv [61].
Influenza and SARS-CoV-2 viruses use a wider range of ways to evade IFN induction and signaling than RSV, human metapneumovirus (HMPV), and human rhinovirus (HRV).
Viral-Bacterial Interactions
Viral-bacterial interactions play a critical role in the pathogenesis of bacterial infection. Knowing the potential viral interactions with the pathobionts of the microbiota or exogenous pathogens will allow us to predict the severity of respiratory diseases and better adapt the therapeutic approach in these patients. There are three types of interactions:
- The virus potentiates bacterial colonization
- Proteases of respiratory tract bacteria cause structural changes leading to increased pathogenicity and tissue tropism of the virus and
- Bacteria enhance viral infection by activating host proteases [62, 63, 64, 65, 66].
The cooperation of viruses and bacteria can cause respiratory diseases that are more severe than those caused by either pathogen alone. Тhis also accounts for the fact that the majority of deaths during influenza epidemics are due to secondary bacterial infections [67, 68, 69, 70, 71]. Positive virus-bacterial interactions are extremely important to us:
- Facilitating bacterial colonization on airway surfaces through damage to the mucociliary system from a previous viral attack. When mechanical defenses are impaired, in primary ciliary dyskinesia, bacterial lung infection is more common [72]. RSV causes loss of cilia in human bronchial cells in vitro, and influenza virus damages the ciliated epithelium and bronchial epithelial lining [73, 74].
- A virus-induced change in the membrane potential of host cells - the most likely cause of increased bacterial adhesion. Viral glycoproteins expressed on host cell membranes can serve as receptors for bacteria. Hemagglutinin esterase [HA] of influenza virus on infected MDCK cells acts as a receptor for group B streptococci. The change in the glucoconjugate structure of murine nasopharyngeal mucosa caused by influenza infection is associated with changes in lectin binding patterns [75, 76]. As a possible mechanism for adherence of staphylococci to virus-infected cells in vivo, it is assumed that infected cells may be coated by a viral antibody serving as a receptor for staphylococcal protein A. Staphylococci appear to attack only those parts that are damaged by the virus. Damage to the respiratory epithelium by other viruses may also have a similar effect [31, 77].
- Suppression of the host’s defense mechanisms against bacterial attack by the viral infection - the non-specific humoral factors, the non-specific phagocytosis by neutrophils and macrophages at the beginning of the infection and the later specific antibody-mediated immune response. Influenza virus-induced polymorphonuclear dysfunction is an important condition for influenza virus-potentiated secondary pneumococcal diseases. After interaction with influenza virus in human neutrophils in vitro, a reduced chemotaxis and phagocytic activity towards staphylococci was observed [78] and a decrease in the bactericidal power of neutrophils and macrophages due to disruption of lysozyme production by both types of phagocytes [79, 80]. Influenza virus is the most studied example of a positive cooperation between a virus and bacteria. Influenza primarily causes upper respiratory tract infections, but when the lungs are affected it can be fatal due to pulmonary edema and hemorrhage. However, most deaths during influenza epidemics are due to secondary bacterial infections.
- Seasonal relationship in viral and bacterial infections- influenza, pneumococcal infection and meningococcal disease (winter months) [67, 69, 70].
Similar interactions with S. pneumoniae are found in RSV and PIV [71, 81]. Unlike most respiratory viruses, seasonal coronaviruses (HCoV-229E, HCoV-NL63, HCoV-OC43, and HCoV-HKU
- Аdenovirus и Moraxella catharralis
- RSV и Haemophilus influenzae
- RSV и Streptococcus pneumoniae
- Rhinovirus А и Streptococcus pneumoniae
- Rhinovirus А и Haemophilus spp
- Rhinovirus С и Streptococcus pneumoniae
- Rhinovirus С и Moraxella catharralis
- Influenza A virus и Streptococcus pneumoniae
- Influenza A virus и Staphylococcus aureus
- Influenza A virus и Neisseria meningitidis.
Bacterial-Bacterial Interactions
Sometimes our enemies’ competitors are our allies. Nasopharyngeal colonization with S. pneumoniae protects us from S. aureus, and reduction of the microbial count of S. pneumoniae after pharmacological intervention leads to increased presence of S. aureus. Decreased colonization by S. pneumoniae also results in increased colonization by H. influenzae, S. aureus, N. meningitidis, and M. catarrhalis [86, 87, 88, 89]. Therefore, we need to carefully consider our antibiotic interventions. Another interesting phenomenon is that pneumococcal vaccines (PCV and PPSV) due to limited coverage induce replacement of vaccine serotypes of S. pneumoniae with non-vaccine serotypes in the nose and nasopharynx and increased carriage of non-typing Haemophilus influenzae. Although they reduced disease caused by the vaccine serotypes, overall colonization rates did not change. The imbalance of the microbiota following acute respiratory infections enables opportunistic pathogens to spread to adjacent poorer ecological niches and cause disease. Cocolonization and polymicrobial interactions between pneumococci and other respiratory colonizers, non- typing H. influenzae and M. catarrhalis lead to an increased risk of acute otitis media [90].
Hypothesis
My hypothesis is based on the importance of maintaining at a constant level the ratios X (surface area), Y (amount of nutrients) and Z (number of microorganisms).
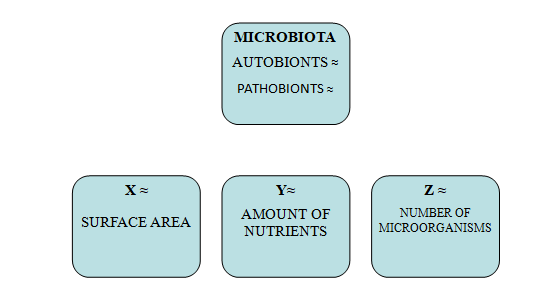
The human microbiota is a complex community of autobionts and pathobionts, and its main function is to protect us against colonization by opportunistic pathogens. Interactions, direct or indirect, between microorganisms and the host include competition for nutrients, ability to attach to surfaces, and induction of host metabolite production [91, 92, 93, 94]. The microbial composition is influenced by epidemiological factors, comorbidities, environment [pollution, radiation], diet, socioeconomic conditions - housing, health care, hygiene, family size (correlated with increased carriage of M. catharralis, S. aureus and antibiotic-resistant strains of S. pneumonia), lifestyle (tobacco smoking, travel and physical activity), vaccinations and intake of antimicrobial drugs [95, 96].
Sequencing patients at disease onset will help us identify elevated viral carriage compared to healthy controls, and with laboratory platforms to capture potential pathogens, we will detect bacterial carriage. Through rapid NAATs, we will isolate the viral agents: SARS-CoV-2, influenza, RSV, etc. These infections are usually difficult to distinguish based on clinical manifestations alone. Viral coinfection triggers respiratory bacterial infections in the nose and nasopharynx, causing destabilization preceding the infection itself and dysbiosis. Whole genome sequencing (WGS) is more sensitive but more expensive in detecting pathogens after antimicrobial drug treatment, and fluorescence in situ hybridization (FISH) has greater sensitivity in detecting bacterial pathogens in biofilm [97]. The factors determining a person’s susceptibility to infection will also give us an opportunity to determine the risk and an easier choice for treatment. These are age, sex, health status, diet, exposure to the pathogen, co-infection, current state of immunity and the individual’s genome. Various studies have shown that microbial diversity increases up to a certain age, and comorbidity is generally associated with a more severe course and more frequent complications. In disease, the amount of autobionts decreases, and that of pathobionts and exogenous pathogens increases.
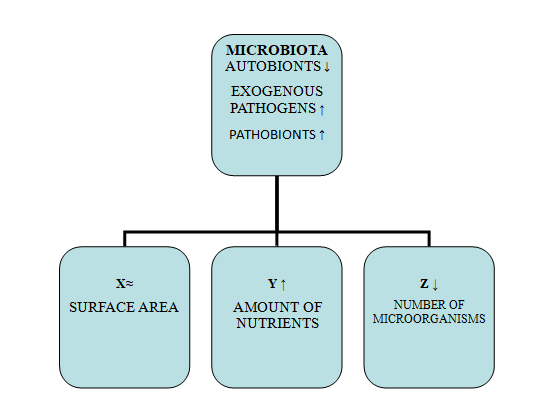
Another important reason for the occurrence of dysbiosis is antibiotic therapy. A 65% increase in the use of antibiotics between 2000 and 2015 has been observed worldwide [98]. They are also the most common cause of decreased resistance to colonization and a drastic change in the composition and diversity of the microbiota. Studies in mouse models have demonstrated disruption of the microbial community of the gastrointestinal tract (change in number and diversity) during antibiotic therapy. Broad-spectrum antibiotics do not differentiate between commensal and pathogenic bacteria and can lead to a 30% reduction in the microbiota [99, 100]. Depletion of commensal bacteria by antibiotic treatment results in excess nutrients that can be used by pathobionts and external pathogens. Separately, pathogens, through the released toxins, provide themselves with an additional nutritional advantage necessary for their survival [101]. Another major problem is the increasing rate of antimicrobial resistance (AMR) in recent decades, which is defined as a critical threat to public health. My hypothesis is based on the possibility of limiting the attachment and entry of viruses into cells and preventing, as far as possible, positive interactions virus-virus, virus-bacteria and between bacteria through controlled administration of probiotics, local vaccines and dietary components with the aim of artificial colonization.
At the first symptoms of an acute viral infection, the use in the form of sprays of agents containing sulfated polysaccharides-carrageenans, silver ions and eucalyptus oil will help us to reduce the attachment and entry of the virus into the target cells of the nasopharynx by means of:
- Inhibition of viral attachment by i-carrageenan (Proven in influenza A and human coronavirus OC43)
- Limiting bacterial colonization of the surface of the airways, probably in a manner similar to the cell-free environment of Corynebacterium striatum limiting adhesion
- Reducing the possibility of virus-induced changes in the membrane potential of host cells and increased bacterial adhesion and
- Hindering the use of the viral glycoproteins expressed on the membranes of the host cells for bacterial receptors [102, 103].
Probiotics, Local Vaccines and Dietary Components
The use of probiotics, local vaccines and dietary components for artificial colonization will help us partially compensate for the impaired functions of the microbiota, hinder the sharing of genes responsible for AMR transmission and limit the possibility of active invasive infection. The amount of nutrients is relatively constant in the human body. Probiotics are defined by Fuller as live organisms contained in food that, when ingested, can alter the gut microbiota and stimulate the immune system after ingestion. Admitted in sufficient quantity, they have a certain benefit for the health of humans and animals. They have already proven to be an alternative to antibiotics in poultry [104]. The main properties of probiotics are: acid resistance, specificity, lack of side effects, reduction of pathogenic microbial numbers and viability during storage. By using transient strains to compete for adhesion and nutrients with those with pathogenic potential [pathobionts and exogenous pathogens], we will be able to some extent to prevent the invasion and overgrowth of the latter [105]. Early use of preparations such as topical vaccines containing α hemolytic streptococci AНS [Streptococcus sanguis, Streptococcus mitis, Streptococcus salivarius and Streptococcus oralis] or oral supplements can also help us to strain and deplete energy resources in the upper respiratory tract and gastrointestinal tract. In this way, we will reduce the opening opportunity for both pathobionts and external bacterial pathogens to take advantage of the increased opportunities for nutrient utilization and surface attachment due to the reduction in autobiont microbial numbers. AНS have an inhibitory effect on H. influenzae and S. aureus, Streptococcus oralis alone on S. pneumoniae and Streptococcus salivarius on S. pyogenes and S. pneumoniae. Commensals of the genus Streptococcus promote cross- reactive immunity through the expression of a capsular antigen with genetic and antigenic similarities to those of S. pneumonia [106]. Administration of an intranasal vaccine with Bacillus subtilis to enhance local immunity of the nasal mucosa holds promise [107].
Diet influences the microbial composition of the microbiota. Long-term dietary patterns are part of the factors determining the enterotype of the intestinal microbiota [108]. The Bacteroides enterotype is associated with reduced microbial activity and genetic diversity, insulin resistance and risk of obesity and non-alcoholic steatosis, and the Prevotella enterotype with a diet with a predominance of plant carbohydrates [109]. The changes that occur from the type of diet play a role in the health-disease balance. The microbiota in infants is much richer even than that of children fed with adapted milk with added Dolosigranulum and Corynebacterium [27]. The genus Lactobacillus is a common inhabitant of the gastrointestinal tract of humans and animals and, due to its probiotic characteristics, is recognized as safe [110]. These lactic acid bacteria with a protective function were described by Döderlein and are used in the food industry. Their surface protein layer has an antimicrobial inhibitory effect expressed in competition for binding sites on the surface of the host’s epithelial cells, and for viruses in antagonism to entry and replication, but not to attachment. The genus contains hydrogen peroxide- producing (HP+) and hydrogen peroxide-non-producing (HP-) strains. Depending on the current balance, hydrogen peroxide-producing (HP+) lactobacilli function in a protective, indifferent or abdicating state (patients with infection). The ability of Lactobacillus to adhere to epithelial surfaces is critical for maintaining persistent colonization in the gut and other mammalian tissues. Lactobacillus delbrueckii subsp. is a well-studied member of this genus. It is used in the industrial fermentation of milk. Bulgrian kiselo mljako not yogurt thanks to Lactobacillus bulgaricus is a natural probiotic with a strong beneficial effect in humans. Kiselo mljako is a widespread food among the Bulgarian population. A dairy-rich diet is associated with a richer and more diverse microbiota. A study shows that the nasal microbiota in dairy farmers is more complex and protects against infection by out-competing S. aureus colonization [111]. Lactobacillus acidophilus is another studied representative found in the human gut and a component of many probiotics. Qiu, et al. reported that dietary supplementation with Bacillus subtilis significantly increased Lactobacillus and Bifidobacterium counts in the ileum and cecum, and decreased coliforms and Clostridium perfringens in the cecum.
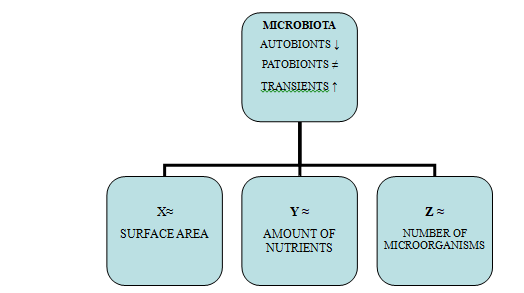
Negative Interactions
Nutritional interventions and probiotics will allow us to benefit from the negative interactions provided by their components:
- Lactobacillus rhamnosus GG, Bifidobacterium Spp, Lactobacillus acidophilus and Streptococcus thermophilus significantly reduce nasal colonization with potential pathogens including - S. aureus, S. pneumoniae and β hemolytic streptococci, thereby reducing respiratory infections. [112, 113, 114]
- Newborns given prebiotics and probiotics have a significantly lower incidence of respiratory tract infections. Those taking Lactobacillus plantarum had a significant reduction in respiratory tract infections [115, 116, 117].
- S. salivarius and S. oralis significantly reduced the abundance of S. aureus and increased the total number of beneficial microorganisms [118].
- Bifidobacterium infantis has an anti-inflammatory effect by modifying the IL-10/IL-12 ratio. It has an immunomodulating effect by increasing mucosal dendritic cells and decreasing Th1 and Th7 helper T cells.
- Lactobacillus саsеi expressing the S. pneumoniae protein antigen PspA induces antibodies that protect mice from disease [119].
- Bifidobacterium longum when administered orally in mice induces reactive oxygen species on alveolar macrophages, enhancing protection against lung infection caused by Klebsiella pneumonia [120].
Conclusion
Cocirculation of respiratory pathogens can lead to competitive or cooperative forms of interaction between them. Probably in the coming years, if not decades, we will be able to answer themselves the following two questions: - Which of the species represented in the probiotics offered on the market will not only be transient, but will also adapt to the receptors on the epithelial surfaces - Whether frequent and long-term intakes of probiotics will not make the human microbiota “lazy” and whether in 20 years we will not be looking for the reasons that made their use meaningless, as is the case with antibiotics. We must not forget that we are carriers of 20-25,000 genes, and up to 10 million useful genes have been detected in our feces. The commensals presented in probiotics are artificially cultured and it is not known whether they fully adapt to the ecological niches normally occupied by them and whether they carry the same genes. Replacing our microbiota can lead to unpredictable macroorganism responses.
The diet participates in the formation of the composition of the microbiota, and the changes that occur due to the type of diet play an important role in the health-disease balance. Providing a greater diversity and abundance of competing microorganisms for food resources and adhesion receptors on epithelial surfaces will likely allow us to help restore resistance against microbiota colonization [121].
The use before the autumn winter season and at the beginning of the disease of preparations such as topical vaccines containing α hemolytic streptococci ANS [Streptococcus sanguis, Streptococcus mitis, Streptococcus salivarius and Streptococcus oralis] or oral supplements can help us to increase the microbial diversity and reduce the possibility of both pathobionts , as well as external bacterial pathogens to take advantage of the increased opportunities for nutrient utilization and surface attachment in URT infection. We need to understand how our interventions do not cause a disturbance in the balance of the human microbiota - reduced species diversity, altered metabolic activity, stimulating the selection and sharing of genes responsible for AMR and from there reducing the rate of relapses. Knowing the interactions between commensal and opportunistic species, and the reasons they jump from one group to the other, will help us determine the time to move from non-pharmacological to pharmacological interventions, preserving the balance of the microbiota and that between it and the macroorganism. The correct combination of pharmacological, non-pharmacological and dietary interventions in the personalized treatment of patients will allow us to better control the risk in the course of the diseases.
References
-
Costello EK, Lauber CL, Hamady M, Fierer N, Gordon JI, et al. (2009) Bacterial community variation in human body habitats across space and time. Science 326(5960): 1694-1697.
-
Human Microbiome Project Consortium (2012) Structure, function and diversity of the healthy human microbiome. Nature 486(7402): 207-214.
-
Iliev ID, Funari VA, Taylor KD, Nguyen Q, Reyes CN, et al. (2012) Interactions between commensal fungi and the C-type lectin receptor Dectin-1 influence colitis. Science 336(6086): 1314-1317.
-
Reyes A, Semenkovich NP, Whiteson K, Rohwer F, Gordon JI (2012) Going viral: Next-generation sequencing applied to phage populations in the human gut. Nat Rev Microbiol 10(9): 607-617.
-
Caufield PW, Dasanayake AP, Li Y, Pan Y, Hsu J, et al. (2000) Natural history of Streptococcus sanguinis in the oral cavity of infants: evidence for a discrete window of infectivity. Infect Immun 68(7): 4018-4023.
-
Hauser LJ, Ir D, Kingdom TT, Robertson CE, Frank DN, et al. (2016) Investigation of bacterial repopulation after sinus surgery and perioperative antibiotics. Int Forum Allergy Rhinol 6(1): 34-40.
-
Macpherson AJ, Harris NL (2004) Interactions between commensal intestinal bacteria and the immune system. Nat Rev Immunol 4(6): 478-485.
-
Donkor ES (2013) Understanding the pneumococcus: transmission and evolution. Front Cell Infect Microbiol 3: 7.
-
Mechergui A, Achour W, Baaboura R, Ouertani H, Lachal A, et al. (2014) Case report of bacteremia due to Neisseria mucosa. APMIS 122(4): 359-361.
-
Clemente JC, Ursell LK, Parfrey LW, Knight R (2012) The impact of the gut microbiota on human health: Аn integrative view. Cell 148(6): 1258-1270.
-
Honda K, Littman DR (2012) The microbiome in infectious disease and inflammation. Annu Rev Immunol 30: 759-795.
-
Wei M, Shinkura R, Doi Y, Maruya M, Fagarasan S, et al. (2011) Mice carrying a knock-in mutation of Aicda resulting in a defect in somatic hypermutation have impaired gut homeostasis and compromised mucosal defense. Nat Immunol 12(3): 264-270.
-
Kawamoto S, Tran TH, Maruya M, Suzuki K, Doi Y, et al. (2012) The inhibitory receptor PD-1 regulates IgA selection and bacterial composition in the gut. Science 336(6080): 485-489.
-
Olszak T, An D, Zeissig S, Vera MP, Richter J, et al. (2012) Microbial exposure during early life has persistent effects on natural killer T cell function. Science 336(6080): 489- 493.
-
Naik S, Bouladoux N, Wilhelm C, Molloy MJ, Salcedo R, et al. (2012) Compartmentalized control of skin immunity by resident commensals. Science 337(6098): 1115- 1119.
-
Bloom SM, Bijanki VN, Nava GM, Sun L, Malvin NP, et al. (2011) Commensal Bacteroides species induce colitis in host-genotype-specific fashion in a mouse model of inflammatory bowel disease. Cell Host Microbe 9(5): 390-403.
-
Clark SE (2020) Commensal bacteria in the upper respiratory tract regulate susceptibility to infection. Curr Opin Immunol 66: 42-49.
-
Huxley EJ, Viroslav J, Gray WR, Pierce AK (1978) Pharyngeal aspiration in normal adults and patients with depressed consciousness. Am J Med 64(4): 564- 568.
-
Bassis CM, Erb-Downward JR, Dickson RP, Freeman CM, Schmidt TM, et al. (2015) Analysis of the upper respiratory tract microbiotas as the source of the lung and gastric microbiotas in healthy individuals. mBio 6(2): e00037.
-
Segal LN, Alekseyenko AV, Clemente JC, Kulkarni R, Wu B, et al. (2013) Enrichment of lung microbiome with supraglottic taxa is associated with increased pulmonary inflammation. Microbiome 1(1): 19.
-
Bosch AATM, De Steenhuijsen Piters WAA, Van Houten MA, Chu MLJN, Biesbroek G, et al. (2017) Maturation of the infant respiratory microbiota, environmental drivers, and health consequences. A prospective cohort study. Ame J Respir Crit Care Med 196(12): 1582-1590.
-
Teo SM, Mok D, Pham K, Kusel M, Serralha M, et al. (2015) The infant nasopharyngeal microbiome impacts severity of lower respiratory infection and risk of asthma development. Cell Host Microbe 17(5): 704-715.
-
Ivanov II, Honda K (2012) Intestinal commensal microbes as immune modulators. Cell Host & Microbe 12(4): 496-508.
-
Brown RL, Sequeira RP, Clarke TB (2017) The microbiota protects against respiratory infection via GM-CSF signaling. Nat Commun 8(1): 1512.
-
Cait A, Hughes MR, Antignano F, Cait J, Dimitriu PA, et al. (2017) Microbiome–driven allergic lung inflammation is ameliorated by short-chain fatty acids. Mucosal Immunol 11(3): 785-795.
-
Man WH, Steenhuijsen PWAA, Bogaert D (2017) The microbiota of the respiratory tract: gatekeeper to respiratory health. Nat Rev Microbiol 15(5): 259-270.
-
Flynn M, Dooley J (2021) The microbiome of the nasopharynx. J Med Microbiol 70(6): 001368.
-
Man WH, Steenhuijsen C, Piters WAA, Houten MA, Chu MLJN et al. (2019) Loss of microbial topography between oral and nasopharyngeal microbiota and development of respiratory infections early in life. Am J Respir Crit Care Med 200(6): 760-770.
-
Ichinohe T, Pang IK, Kumamoto Y, Peaper DR, jh H et al. (2011) Microbiota regulates immune defence against respiratory tract influenza a virus infection. Proc Natl Acad Sci USA 108(13): 5354-5359.
-
(1998) Topley and Wilson’s Microbiology and Microbial Infection. In: Mahy BWJ, et al. (Eds.), Virology , 9th (Edn.), Arnold, London, United Kingdom, 1.
-
(1998) Topley and Wilson’s Microbiology and Microbial Infection. In: Hausler WJ, et al. (Eds.), Bacterial Infections, 9th (Edn.), Arnold, London, United Kingdom, 3.
-
Kim HJ, Jo A, Jeon YJ, An S, Lee KM, et al. (2019) Nasal commensal Staphylococcus epidermidis enhances interferon-λ- dependent immunity against influenza virus. Microbiome 7(1): 80.
-
Bradley KC, Finsterbusch K, Schnepf D, Crotta S, Llorian M, et al. (2019) Microbiota-driven tonic interferon signals in lung stromal cells protect from influenza virus infection. Cell Rep 28(1): 245-256.e4.
-
Liu Q, Liu Q, Meng H, Lv H, Liu Y, et al. (2019) Staphylococcus epidermidis contributes to healthy maturation of the nasal microbiome by stimulating antimicrobial peptide production. Cell Host Microbe 27(1): 68-78.
-
Janek D, Zipperer A, Kulik A, Krismer B, Peschel A (2016) High frequency and diversity of antimicrobial activities produced by nasal Staphylococcus strains against bacterial competitors. PLoS Pathog 12(8): e1005812-e1005820.
-
Latham RD, Gell DA, Fairbairn RL, Lyons AB, Shukla SD, et al (2017) An isolate of Haemophilus haemolyticus produces a bacteriocin-like substance that inhibits the growth of nontypeable Haemophilus influenzae. Int J Antimicrob Agents 49(4): 503-506.
-
Engen SA, Valen RH, Becattini S, Jarrossay D, Blix IJ, et al. (2014) The oral commensal Streptococcus mitis shows a mixed memory Th cell signature that is similar to and cross-reactive with Streptococcus pneumoniae. PLoS One 9(8): e104306-e104309.
-
Zipperer A, Konnerth MC, Laux C, Berscheid A, Janek D, et al. (2016) Human commensals producing a novel antibiotic impair pathogen colonization. Nature 535(7613): 511-516.
-
Manning J, Dunne EM, Wescombe PA, Hale JDF, Mulholland EK, et al. (2016) Investigation of Streptococcus salivarius- mediated inhibition of pneumococcal adherence to pharyngeal epithelial cells. BMC Microbiol 16(1): 225.
-
Bomar L, Brugger SD, Yost BH, Davies SS, Lemon KP (2016) Corynebacterium accolens releases antipneumococcal free fatty acids from human nostril and skin surface triacylglycerols. mBio 7(1): e1725-1815.
-
Ramsey MM, Freire MO, Gabrilska RA, Rumbaugh KP, Lemon KP (2016) Staphylococcus aureus shifts toward commensalism in response to Corynebacterium species. Front Microbiol 7: 1230.
-
Santagati M, Scillato M, Patane F, Aiello C, Stefani S (2012) Bacteriocin-producing oral streptococci and inhibition of respiratory pathogens. FEMS Immunol Med Microbiol 65(1): 23-31.
-
Characklis WG (1973) Atteched microbial growths, ІІ. Frictional resistance due to microbial slimes. Water Res 7(9): 1249-1258.
-
Potera C (1999) Forging a link between biofilms and disease. Science 283(5409): 1837-1839.
-
Lim DJ, Cotticchia JM, Ueno K, Heiselman FA, Bakaletz LO (1991) Glycoconjugates in the chinchilla tubotympanum. Ann Otol Rhinol Laryngol 100(11): 933-943.
-
Ramadan HH, Sanclement JA, Thomas JG (2005) Chronic rhinosinusitis and biofilms. Otolaryngol Head Neck Surg 132(3): 414-417.
-
Costerton JW, Stewart PS, Greenberg EP (1999) Bacterial biofilms: a common cause of persistent infections. Science 284(54): 1318-1322.
-
Wood AJ, Fraser JD, Swift S, Patterson-Emanuelson EA, Amirapu S, et al. (2012) Intramucosal bacterial microcolonies exist in chronic rhinosinusitis without inducing a local immune response. Am J Rhinol Allergy 26(4): 265-270.
-
Subtil J, Bajanka-Lavado M, Rodrigues J, Duarte A, Reis L, et al. (2018) Cross-sectuional study of adenoidal biofilms in pediatric population and its clinical implications. Otolar Polska 73(1): 1-5.
-
Wylie K, Mihindukulasuriya K, Sodergren E, Weinstock G, Storch G (2012) Sequence analysis of the human virome in febrile and afebrile children. PLOS One 7(6): e27735.
-
Wong A, Guevara LAB, Goult E, Briga M, Kramer SC, et al. (2023) The interactions of SARS-CoV-2 with cocirculating pathogens: Epidemiological implications and current knowledge gaps. PLOS Pathogens 19(3): e1011167.
-
Kitchen A, Shackelton LA, Holmes EC (2011) Family level phylogenies reveal modes of macroevolutionin RNA viruses. Proc Nat Acad Sci USA 108(1): 238-243.
-
Xiao-Ying C, Wang Q, Guang-Yu L, Zhi-Wei C, Chuang- Xing L, et al. (2014) Respiratory virus infections among children in South China. J Med Virol 86(7): 1249-1255.
-
I-Chueh H, Li W, Sui J, Marasco W, Choe H, et al. (2008) Influenza A virus neuraminidase limits viral superinfection. J Virol 82(10): 4834-4843.
-
Laurie KL, Horman W, Carolan LA, Chan KF, Layton D, et al. (2018) Evidence for viral interference and crossreactive protective immunity between influenza B virus lineages. J Infect Dis 217(4): 548-559.
-
Chan KF, Carolan LA, Korenkov D, Druce J, McCaw J, et al. (2018) Investigating viral interference between influenza A virus and human respiratory syncytial virus in a ferret model of infection. J Infect Dis 218(3): 406- 417.
-
Schneider WM, Chevillotte MD, Rice CM (2014) Interferonstimulatedgenes: a complex web of host defenses. Annu RevImmunol 32: 513-545.
-
Voroshilova MK (1989) Potential use of nonpathogenic enteroviruses for control of human disease. Prog Med Virol 36: 191-202.
-
Chumakov MP, Voroshilova MK, Antsupova AS, Boĭko VM, Blinova MI, et al. (1992) Live enteroviral vaccines for the emergency nonspecific prevention of mass respiratory diseases during fall-winter epidemics of influenza and acute respiratory diseases [in Russian]. Zh Mikrobiol Epidemiol Immunobiol (11-12): 37-40.
-
Shinjoh M, Omoe K, Saito N, Matsuo N, Nerome K (2000) In vitro growth profiles of respiratory syncytial virus in the presence of influenza virus. Acta Virol 44(2): 91-97.
-
Piret J, Boivin G (2022) Viral interference between respiratory viruses. Emerg Infect Dis 28(2): 273-281.
-
Nickbakhsh S, Mair C, Matthews Llouise, Reeve R, Johnson PCD, et al. (2019) Virus-Virus interactions impact the population dynamics of influenza and common cold. Proc of Nat Acad of Science 116(52): 27142-27150.
-
Rott R, Klenk HD, Nagai Y, Tashiro M (1995) Influenza viruses, cell enzymes and pathogenicity. Am. J. Respir. Crit. Care Med 152: S16-S19.
-
Tashiro M, Ciborowski P, Klenk H, Pulverer G, Rott R (1987) Role of staphylococcus protease in the development of influenza pneumonia. Nature 325: 536- 537.
-
Tashiro M, Ciborowski P, Reinacher M, Pulverer G, Klenk HD, et al. (1987) Synergistic role of staphylococcal proteases in the induction of influenza virus pathogenicity. Virology 157(2): 421-430.
-
Akaike T, Molla A, Ando M, Araki S, Maeda H (1989) Molecular mechanism of complex infection by bacteria and virus analyzed by a model using serratial protease and influenza virus in mice. J Virol 63(5): 2252-2259.
-
Kilbourne ED (1987) Influenza. Plenum Publishing Corp, New York, US.
-
Schwarzmann SW, Adler JL, Sullivan RJ, Marine WM (1971) Bacterial pneumonia during the Hong Kong influenza epidemic of 1968–69. Experience in a city- county hospital. Arch Intern Med 127: 1037-1041.
-
Taubenberger JK, Reid AH, Fanning TG (2000) The 1918 influenza virus: a killer comes into view. Virology 274(2): 241-245.
-
Cartwright KAV, Jones DM, Smith AJ, Stuart JM, Kaesmarski EB, et al. (1991) Influenza A and meningococcal disease. Lancet 338(8766): 554-557.
-
Kim PE, Musher DM, Glezen WP, Rodriguez-Barradas MC, Nahm WK, et al. (1996) Association of invasive pneumococcal disease with season, atmospheric conditions, air pollution and isolation of respiratory viruses. Clin Infect Dis 22(1): 100-106.
-
Reynolds HY, Root EK (1991) Bronchiectasis and broncholithiasis. In: Wilson JD, Braunwald E, Isselbacher KJ, Petersdorf RG, Martin JB, et al. (Eds.), Harrison’s Principles of Internal Medicine, New York, US, pp: 1069- 1071.
-
Tristam D, Hicks W, Hard R (1998) Respiratory syncytial virus and human bronchial epithelium. Arch. Otolaryngol. Head Neck Surg 124(7): 777-783.
-
Walsh J, Dietlein L, Low F, Burch G, Mogabgab W (1960) Bronchotracheal response in human influenza Type A, Asian strain, as studied by light and electron microscopic examination of bronchoscopic biopsies. Arch Intern Med 108(3): 376-388.
-
Hirano T, Kurono Y, Ichimiya I, Suzuki M, Mogi G (1999) Effects of influenza A virus on lectin-binding patterns in murine nasopharyngeal mucosa and on bacterial colonization. Otolaryngol Head Neck Surg 121(5): 616- 621.
-
Sanford BA, Shelokov A, Ramsay MA (1978) Bacterial adherence to virus infected cells: a cell culture model of bacteria super-infection. J Infect Dis 137(2): 176-181.
-
Austin RM, Daniels CA (1978) The role of protein A in attachment of staphylococci to influenza-infected cells. Lab Investig 39(2): 128-132.
-
Larson HE, Parry RP, Gilchrist C, Luquetti A, Tyrrell DAJ (1977) Influenza viruses and staphylococci in vitro: some interactions with polymorphonuclear and epithelial cells. Br J Exp Pathol 58(3): 281-292.
-
Warr GA, Jakab GJ, Chan TW, Tsan MF (1979) Effects of viral pneumonia on lung macrophage lysosomal enzymes. Infect Immun 24(2): 577-579.
-
Pang G, Clancy R, Gong M, Ortega M, Ren ZG, et al. (2000) Influenza virus inhibits lysozyme secretion by sputum neutrophils in subjects with chronic bronchial sepsis. Am J Respir Crit Care Med 161(3 Pt 1): 718-722.
-
Fiore AE, Iverson C, Messmer T, Erdman D, Lett SM, et al. (1998) Outbreak of pneumonia in a long term care facility: antecedent human parainfluenza virus 1 infection may predispose to bacterial pneumonia. J Am Geriatr Soc 46(9): 1112-1117.
-
Makela MJ, Puhakka T, Ruuskanen O, Leinonen M, Saikku P, et al. (1998) Viruses and bacteria in the etiology of the common cold. J Clin Microbiol 36(2): 539-542.
-
Klenk HD, Rott R, Orlich M (1977) Further studies on the activation of influenza virus by proteolytic cleavage of the haemagglutinin. J Gen Virol 36(1): 151-161.
-
Scheiblauer H, Reinacher M, Toshiro M, Rott R (1992) Interaction between bacteria and influenza A virus in the development of influenza pneumonia. J Infect Dis 166(4): 783-791.
-
Moore H, Jacoby P, Taylor A, Harnett G, Bowman J (2010) The interaction between respiratory viruses and pathogenic bacteria in upper respiratory tract of asymptomatic aboriginal and nonaboriginal children. Ped Infect Dis J 29(6): 540-545.
-
Regev-Yochay G, Dagan R, Raz M, Carmeli Y, Shainberg B, et al. (2004) Association between carriage of Streptococcus pneumoniae and Staphylococcus aureus in Children. JAMA 292(6): 716-720.
-
Bogaert D, De Groot R, Hermans PWM (2004) Streptococcus pneumoniae colonisation: the key to pneucoccal disease. Lancet infect Dis 4(3): 144-154.
-
Watson k, Carville K, Bowman J, Peter J, Victor RT, et al. (2006) Upper respiratory tract bacterial carriage in Aboriginal and non- Aboriginal children in semi-arid area of Western Australia. Pediatr Infect Dis J 25(9): 782- 790.
-
Chan WY, Cohen J, Brown J (2016) The new first-line defense: The potential of nasopharyngeal colonization in vaccine strategies. Vaccine development and therapy 6: 47-57.
-
Revai K, Mamidi D, Chonmaitree T (2008) Association of nasopharyngeal bacterial colonization during upper respiratory tract infection and the development of acute otitis media. Clin Infect Dis 46(4): 34-37.
-
Mansbach JM, Hasegawa K, Piedra PA, Avadhanula V, Petrosino JF, et al. (2020) Haemophilus-dominant nasopharyngeal microbiota is associated with delayed clearance of respiratory syncytial virus in infants hospitalized for bronchiolitis. J Infect Dis 94: 107-109.
-
Britton RA, Young VB (2014) Role of the intestinal microbiota in resistanceto colonization by Clostridium difficile. Gastroenterology 146(6): 1547-1553.
-
Lawley TD, Walker AW (2013) Intestinal colonization resistance. Immunology 138(1): 1-11.
-
Sorbara MT, Pamer EG (2019) Interbacterial mechanisms of colonizationresistance and the strategies pathogens use to overcome them. Mucosal Immunol 12(1): 1-9.
-
Kamada N, Kim YG, Sham HP, Vallance BA, Puente JL, et al. (2012) Regulated virulence controls the ability of a pathogen tocompete with the gut microbiota. Science 336(6086): 1325-1329.
-
Cleary D, Devine V, Morris D, Osman K, Gladstone R, et al. (2018) Pneumococcal vaccine impacts on the population genomics of nontypeable Haemophilus influenzae. Microb Genomics 4(9): e000209.
-
Stepinska M, Olszewska-Sosinska O, Lau-Dworak M, Zielnik-Jurkiewicz B, Trafny E (2013) Indentification of intracellular bacteria in adenoid and tonsil tissue specimens: The efficienc of culture versus fluorescent in situ hybridization FISH. Curr Microbiol 68(1): 21-29.
-
Klein EY, Van Boeckel TP, Martinez EM, Pant S, Gandra S, et al. (2018) Global increase and geographic convergence in antibiotic consumption between 2000 and 2015. Proc Natl Akad Sci, USA, 115(15): E3463- E3e70.
-
Neuman H, Forsythe P, Uzan A Avni O, Koren O (2018) Antibiotics in early life : dysbiosis and the damage done. FEMS Microbiol Rev 42(4): 489-499.
-
Francino MP (2015) Antibiotics and the human gut microbiome: dysbiosis and accumulation of resistance. Front Microbiol 6: 1543.
-
Fletcher JR, Pike CM, Parsons RJ, Rivera AJ, Foley MH, et al. (2021) Clostridoides difficile exploits toxin- mediated inflammation to alterthe host nutritional landscape and exclude competitors from the gut microbiota. Nat Commun 12(1): 462.
-
McKim JM, Willoughby Sr JA, Blakemore WR, Weiner ML (2019) Clarifying the confusion between poligeenan, degraded carrageenan, and carrageenan: A review of the chemistry, nomenclature, and in vivo toxicology by the oral route. Crit Rev Food Sci Nutr 59(19): 3054-3073.
-
Hebar A, Koller C, Seifert JM, Chabicovsky M, Bodenteich A, et al. (2015) Non-clinical safety evaluation of intranasal iota-carrageenan. PLoS One 10(4): e0122911.
-
Soren S, Mandal GP, Roy B, Samanta I, Hansda RN (2023) Assessment of Bacillus subtilis based probiotics on health and productive perfomance of poultry: A review. Indian J Anim Health 62(2): 132-138.
-
McFarland LV (2008) Antibiotic-associated diarrhea: epidemiology, trends and treatment. Future Microbiol 3(5): 563-578.
-
Skov Sorensen UB, Yao K, Yang Y, Tettelin H, Kilian M (2016) Capsular polysaccharide expression in commensal Streptococcus species: genetic and antigenic similarities to Streptococcus pneumoniae. mBio 7(6): e01844-16.
-
Yang Y, Jing Y, Yang J, Yang Q (2018) Effects of intranasal administration with Bacillus subtilis on immune cells in the nasal mucosa and tonsils of piglets. Exp Ther Med 15(6): 5189-5198.
-
Costea PI, Hildebrand F, Arumugam M, Backhed F, Blaser MJ, et al. (2018) Enterotypes in the landscape of gut microbial community composition. Nat Microbiol 3(1): 8-16.
-
Wu GD, Chen J, Hoffmann C, Bittinger K, Chen YY, et al. (2011) Linking long-term dietary patterns with gut microbial enterotypes. Science 334(6052): 105-108.
-
Walter J, Heng NCK, Hammes WP, Loach DM, Tannock GW, et al. (2003) Identification of Lactobacillus reuteri genes specifically induced in the mouse gastrointestinal tract. Appl Environ Microbiol 69(4): 2044-2051.
-
Shukla SK, Ye Z, Sandberg S, Reyes I, Fritsche TR, et al. (2017) The nasal microbiota of dairy farmers is more complex than oral microbiota, reflects occupational exposure, and provides competition for staphylococci. PLoS One 12(8): e0183898.
-
Konstantinov SR, Smidt H, Vos WMD, Bruijns SCM, Singh SK et al. (2008) S layer protein A of Lactobacillus acidophilus NCFM regulates immature dendritic cell and T cell functions. Proc Natl Acad Sci USA 105 (49): 19474- 19479.
-
Gluck U, Gebbers JO (2003) Ingested probiotics reduce nasal colonisation with pathogenic bacteria (Staphylococcus aureus, Streptococcus pneumoniae and β hemolytic streptococci). Am j Clin Nutr 77(2): 517-520.
-
Bernstein JM, Haase E, Scannapieco F, Dryja D, Wolf J, et al. (2006) Bacterial interference of penicillin- sensitive and resistant Streptococcus pneumoniae by Streptococcus oralis in adenoid organ culture: Implications for the treatment of recurrent upper respiratory tract infections in children and adults. Ann Otol Rhinol Laryngol 115(5): 350-356.
-
Di Pierro F, Donato G, Formia F, Adami T, Careddu D, et al. (2012) Preliminary pediatric clinical evaluation of the oral probiotic Streptococcus salivarius K12 in preventing recurrent pharyngitis and/or tonsillitis caused by Streptococcus pyogenes and recurrent acute otitis media . Int J Gen Med 5: 991-997.
-
Luoto R, Ruuskanen O, Waris M, Kalliomäki M, Salminen S, et al. (2014) Prebiotic and probiotic supplementation prevents rhinovirus infections in preterm infants: a randomized, placebo-controlled trial. J Allergy Clin Immunol 133(2): 405-413.
-
Panigrahi P, Parida S, Nanda NC, Satpathy R, Pradhan L, et al. (2017) A randomized synbiotic trial to prevent sepsis among infants in rural India. Nature 548(7668): 407-412.
-
De Grandi R, Drago L, Bidossi A, Bottagisio M, Gelardi M, et al. (2019) Putative microbial population shifts attributable to nasal administration of Streptococcus salivarius 24SMBc and Streptococcus oralis 89a. Probiotics Antimicrob Proteins 11: 1219-1226.
-
Campos IB, Darrieux M, ferreira DM, Miyaji NM, Silva DA, et al. (2008) Nasal immunization of mice with Lactobacillus casei expressing the pneumococcal surface proteinA: induction of antibodies, complement deposition and partial protection against Streptococcus pneumoniae challenge. Microbes Infect 10(5): 481-488.
-
Vieira AT, Rocha VM, Tavares L, Garcia CC, Teixeira MM, et al. (2016) Control of Klebsiella pneumonae pulmonary infection and immunomodulation by oral treatment with commensal probiotic Bifidobacterium longum 51A. Microbes Infect 18(3): 180-189.
-
Logan LK, Weinstein RA (2017) The epidemiology of carbapenem-resistant Enterobacteriaceae: the impact and evolution of global menace. J Infect Dis 215(S1): S28-S36.
- 4th Branchial Cleft Sinus Anomaly Presenting as Recurrent Thyroid Abscess in A Child: A Case Report
- Parotid Duct Injury Repaired Using an Angiocatheter Stent: A Case Report
- Organization and Functionality of the Referral and Counter-Referral System for ENT Disorders in District Hospitals of N’Djamena, Chad: A Cross-Sectional Analytical Study
- Facial Metastases from a Gastrointestinal Stromal Tumor: A Case Report
- Panorama of Ent Cancers and Literature Review: Epidemiological Profile and Therapeutic Management
- Could Antimicrobial Resistance Prove to Be Both a Threat and an Opportunity for us?