The Effect of Coenzyme Q10 on Oxidative Stress and Endoplasmic Reticulum Stress in an Experimental Diabetes Model
Diabetes is the 8th leading cause of death worldwide. The greatest increase in rates has been seen in low and middle-income countries. Urbanization and lifestyle changes such as an increase in sedentary lifestyles, increase in the intake of foods with high energy but poor nutrition levels have caused a high incidence of the disease worldwide. Treating the mechanisms involved in the pathogenesis of diabetes with nutrients such as antioxidants can reduce further complications of the disease. This study tests whether hyperglycemia triggers endoplasmic reticulum stress in the liver and brain tissues of rats and the possible role of oxidative stress in this pathway. We investigated the relationship between oxidative and endoplasmic reticulum stress via the TWEAK pathway, and how the coenzyme Q10 affects further diabetic complications. We examined oxidative stress markers (Oxidized Glutathione, Reduced Glutathione) in the liver and brain tissues of rats in the control, diabetic, and diabetic antioxidant groups by ELISA method. We also determined endoplasmic reticulum stress markers (PERK, ATF6, GRP78) and the TWEAK molecule by the Western Blott method in the same experimental groups. Our results underlined that coenzyme Q10 had a significant and positive effect on oxidative and endoplasmic reticulum stress parameters, and TWEAK levels. We also found a more robust positive correlation between oxidative and endoplasmic reticulum stress parameters for the liver compared to the brain. However, a significant positive correlation was also seen between TWEAK and endoplasmic reticulum stress of the brain.
Introduction
Diabetes Mellitus is a multifactorial disease characterized by hyperglycemia resulting from deficiencies in insulin production or activity [1, 2]. According to the data of the International Diabetes Federation, while there were 285 million diabetes patients in 2010 and 425 million in 2017 in the world, the estimated expectation for 2045 is 629 million people. According to 2017 data, 4 million people die every year due to diabetes complications worldwide [3].
Several different mechanisms such as oxidative stress, inflammation, and endoplasmic reticulum stress play an important role in the pathogenesis of diabetes [4-
6]. In recent years, conducted studies have shown that endoplasmic reticulum stress plays an important role in the loss of function of pancreatic beta cells and the development of insulin resistance. Endoplasmic reticulum stress is the excessive accumulation of unfolded or misfolded proteins in the lumen of the endoplasmic reticulum that occurs in various physiological or pathological conditions where the capacity of the endoplasmic reticulum is exceeded [7, 8, 9, 10]. Intracellular signaling pathways called unfolded protein response (UPR) are activated to restore this disturbed balance in the cell and to get rid of the stress with minimum damage [1].
Protein kinase RNA-like ER kinase (PERK), Activating transcription factor 6 (ATF6) and Inositol-requiring kinase 1 (IRE1) proteins play a role in UPR signaling pathways. Under normal conditions, PERK, ATF6, and IRE1 proteins, which co-exist with the “glucose regulatory protein” (GRP78) and remain in an inactive state, become active by separating from GRP78 when stress exceeding the endoplasmic reticulum capacity is encountered [1]. Active UPR signaling pathways are responsible for restoring homeostasis of proper production, processing, and folding of proteins. They also reduce the protein load of the endoplasmic reticulum by stopping protein translation. GRP78 also helps to reduce endoplasmic reticulum stress by both adhering to misfolded proteins and helping to prevent misfolding during translocation in the endoplasmic reticulum [10].
Under conditions of prolonged and severe endoplasmic reticulum stress, the UPR is unable to maintain normal cellular functions and consequently cell death usually caused by apoptosis is triggered [1, 6, 11]. Increased cell death leads to loss of tissue function and causes various diseases related to endoplasmic reticulum stress [12, 13, 14, 15]. Apoptosis is thought to be the most important cause of pancreatic beta-cell loss, especially in type I and type II diabetes patients [16, 17].
TWEAK (TNF-like weak apoptosis inducer) is a member of the TNF family, which has been suggested to have a proapoptotic effect in various cells [18]. TWEAK binds to its receptor called Fn14 under various pathological conditions and stimulates many mechanisms including cellular growth, proliferation, migration, differentiation, apoptosis, angiogenesis, fibrogenesis, and inflammation. Although there are studies suggesting that there may be a relationship between TWEAK and endoplasmic reticulum stress, there is yet no study that completely explains the mechanism [19, 20].
Oxidative stress occurs with the disruption of the balance between free radicals accumulating in the body and antioxidant defense systems [21]. Oxidative stress causes serious damage to pancreatic β cells and has an important role in the pathogenesis of diabetes [22, 23]. Glutathione is one of the endogenous antioxidants that play a major role in reducing the oxidative stress which increases in the development of diabetes [24]. The ratio between the oxidized (GSSG) and reduced (GSH) forms of glutathione is used as one of the most important indicators of oxidative stress [25].
Coenzyme Q10 is a vitamin-like quinone found at high levels in various tissues. Coenzyme Q10 is also a very powerful antioxidant and plays an important role in protecting cells from the effects of oxidative stress. Studies indicate that patients with diabetes have significantly lower plasma coenzyme Q10 levels compared to normal healthy individuals. It has been suggested that exogenous supplementation of coenzyme Q10 potentially attenuates mitochondrial dysfunction induced by oxidative stress, thereby improving glycemic control in type II diabetes [26, 27, 28, 29]. In addition, recent experimental studies link insulin resistance to a decrease in coenzyme Q10 expression and show that supplementation with coenzyme Q10 can restore insulin sensitivity [30].
In this study, we aimed to examine whether hyperglycemia triggers endoplasmic reticulum stress in the liver and brain tissues, how the possible relationship with oxidative stress develops in this mechanism, and whether a powerful antioxidant coenzyme Q10 reduces endoplasmic reticulum stress in rats in a streptozotocin (STZ)-induced experimental diabetes model. In addition, we aimed to examine the possible connections of the TWEAK molecule with the UPR signaling pathways and to examine whether there is a relationship between them and endoplasmic reticulum stress.
Materials and Methods
Experimental Animals
The experimental protocol was approved by the Bezmialem Experimental Animals Local Ethics Committee. The experiment was carried out on 24 healthy adult male Sprague Dawley rats weighing 300-350 g. Rats were housed in polypropylene cages, maintained under standardized conditions (12 h light/dark cycle, 24°C, 35–60% humidity), and allowed free access to diet and purified drinking water ad libitum.
Experimental Design
Rats were randomly divided into three groups each consisting of eight animals. Group I: Control rats (distilled water 10 ml/kg), Group II: Diabetic rats, Group III: Diabetic rats treated with 10 mg/kg coenzyme Q10 (1% aqueous solution of Tween 80). An experimental diabetes model was developed by administering a single dose of 65 mg/ kg STZ solution (dissolved in citrate buffer, pH 4.5) (cat no. ab142155, Abcam, Cambridge, UK) intraperitoneally to rats in the diabetes and diabetic antioxidant group (31). Hyperglycemia was confirmed by elevated blood glucose levels at 72 h after injection. The animals with fasting blood glucose levels greater than 200 mg/dl were considered diabetic [31, 32]. The diabetic antioxidant group rats were administered 10 mg/kg “coenzymeQ10” (cat no. ab143173, Abcam, Cambridge, UK) intraperitoneally for 42 days [32]. Body weights and fasting blood glucose levels of the rats were measured regularly. At the end of the experiments, anesthesia was performed with 2% isoflurane inhalation [33, 34]. Blood taken from the tail veins of rats was stored at -80oC for ELISA studies. Liver and brain tissues from rats were stored at -80oC for Western Blot studies.
Western Blotting
The protein expressions of the endoplasmic reticulum stress proteins GRP78, ATF6, PERK, and the apoptotic marker TWEAK were assessed by western blotting analysis. Protein isolation from liver and brain tissues was performed with “Mammalian cell extraction kit (cat no. ab65399, Abcam, Cambridge, UK)”. The isolation buffer was prepared with “Extraction buffer - DTT - protease” chemicals as specified in the kit protocol. Tissues were homogenized in ice with this isolation buffer. Protein levels in the resulting homogenates were determined with the BCA kit (cat no. 21071, Intron Biotechnology Inc., S. Korea).
The procedure of western blotting was performed according to the protocol of Marie, et al. [35]. The proteins were separated by SDS-PAGE. Then proteins are transferred to the PVDF membrane. Membranes were probed by primary antibodies overnight at 4 °C. The primary antibodies were used as follows: anti-ATF6 (cat no. ab203119, Abcam, Cambridge, UK) diluted at 1:1000; anti-GRP78 (cat no. ab109659) diluted at 1:2000; anti-PERK (cat no. ab79483) diluted at 1:100; anti-TWEAK (cat no. ab37170) diluted at 1:1000. Then the membrane was incubated with a secondary antibody (1:2000; ab97069, Abcam, Cambridge, UK) at room temperature for 1 h. GAPDH was used as an internal control. The protein bands were visualized with Enhanced Chemiluminescence (ECL) (cat no. K22030, Abbkin Superlumia ECL Plus HRP Substrate Kit, USA) and their densitometry was done using Image J software (Total lab software, Wales, UK).
ELISA
GSSG (cat no. E1264Ra, Bioassay, Shanghai) and GSH (cat no. E1443Ra, Bioassay, Shanghai) levels in the blood taken from the tail veins of the rats on the 42nd day were determined by the ELISA method.
Statistical Analysis
Data analysis was conducted using SPSS statistical analysis software (version 20.0; SPSS Inc, Chicago, IL, USA). Results were expressed as means ± SD for continuous variables. ANOVA test was used to show the differences in protein expressions in liver and brain tissues of rats. Comparison of continuous variables of paired groups was done with Student’s t-test. Correlations between study variables were analyzed using the Pearson correlation method. p-values less than 0.05 were considered statistically significant.
Results
Effect of Coenzyme Q10 Administration on Body Weight
As illustrated in Table 1, there was no significant difference between the body weight in all groups before the experiment (p=0.428). The body weight of diabetic rats showed a significant decrease after the administration of STZ (p=0.017). The weight loss of rats in the diabetic antioxidant group was less than that of the diabetic rats.
| Initial | 21st day | 42nd day | |
|---|---|---|---|
| Control | 296.3 ± 65.4* | 327.5 ± 67.5 | 335.7 ± 57.3 |
| Diabetic | 311 ± 45.2* | 263.3 ± 34.5 | 241 ± 29.9** |
| Coenzyme Q10 | 297 ± 49.8* | 235.3 ± 26.7 | 243.6 ± 34.4 |
Table 1: Effect of coenzyme Q10 on body weight. Values are expressed as mean ± SD; n=8. * p=0.428, ** p=0.017 as compared to init
Effect of Coenzyme Q10 Administration on Blood Glucose Levels
Among the diabetic rats, glucose level was significantly increased when compared to control rats (p=0.000). The glucose levels of the rats in diabetic antioxidant group were found to be statistically significantly lower than those in the diabetic group (p=0.009). Glucose levels of the diabetic antioxidant group at the end of the treatment were found to be statistically significantly lower than on the 21st day of treatment (p=0.001) (Table 2).
| Initial | 21st day | 42nd day | |
|---|---|---|---|
| Control | 94.6 ± 9.8 | 81.4 ± 5.6 | 104 ± 8.7 |
| Diabetic | 104 ± 8.7 | 455.6 ± 140.1 | 496.6 ± 68.3* |
| Coenzyme Q10 | 89.7 ± 12.1 | 536.5 ± 63.1 | 384.8 ± 93.6**# |
Table 2: Effect of coenzyme Q10 on blood glucose levels (mg/dl). Values are expressed as mean ± SD; n=8. * p=0.000 as compared to
Oxidative Stress Parameters
GSH values and GSH/GSSG ratio; were found to be lower in the diabetic group than in the control group (p=0.019, p=0.028), higher in the diabetic antioxidant group than in the diabetic group (p=0.003, p=0.003) and higher in the diabetic antioxidant group than in the control group (p=0.126, p=0.048). GSSG values were found to be higher in the diabetic group compared to the control group (p=0.343) and to be significantly lower in the diabetic antioxidant group than in the diabetic group and control group (p=0.063, p=0.106) (Table 3).
| Initial | 21st day | 42nd day | |
|---|---|---|---|
| Control | 94.6 ± 9.8 | 81.4 ± 5.6 | 104 ± 8.7 |
| Diabetic | 104 ± 8.7 | 455.6 ± 140.1 | 496.6 ± 68.3* |
| Coenzyme Q10 | 89.7 ± 12.1 | 536.5 ± 63.1 | 384.8 ± 93.6** # |
Table 3: Oxidative stress parameters. Values are expressed as mean ± SD; n=8. * p<0.05 as compared to control group, ** p<0.05 as
Brain Tissue Western Blotting Results
TWEAK, GRP78 and ATF6 proteins were found to be statistically significantly higher in the diabetic group than in the control group (p<0.001), and PERK is also higher but not significantly so (p=0.158). TWEAK, GRP78, ATF6, and PERK proteins were found to be statistically significantly lower in the diabetic antioxidant group than in the diabetes group (p<0.01). TWEAK, PERK, and ATF6 proteins were found to be statistically significantly lower in the diabetic antioxidant group than in the control group (p<0.05). GRP78 was found to be statistically significantly higher in the diabetic antioxidant group than in the control group (p=0.005) (Figure 1).
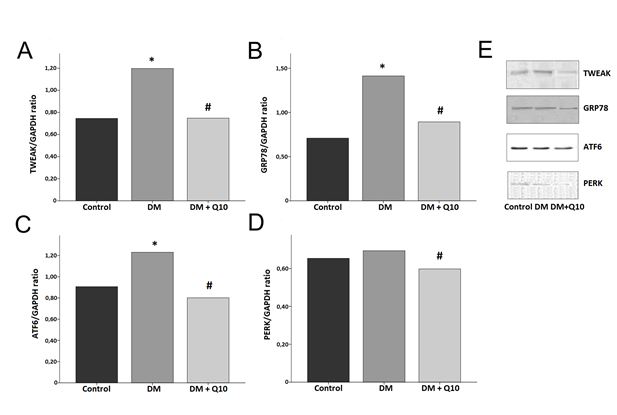
When we look at the correlation relationship between continuous variables of endoplasmic reticulum stress in brain tissue in our study, it is observed that there is a statistically significant positive correlation between PERK and GRP78 (r=0.455, p=0.025), between ATF6 and GRP78 (r=0.750, p=0.000), and between ATF6 and PERK (r=0.436, p=0.033) (Table 4).
| TWEAK | GRP78 | PERK | ATF6 | ||
|---|---|---|---|---|---|
| TWEAK | r | 1 | |||
| TWEAK | p | 0 | |||
| GRP78 | r | 0.56 | 1 | ||
| GRP78 | p | 0.004* | 0 | ||
| PERK | r | 0.187 | 0.455 | 1 | |
| PERK | p | 0.383 | 0.025* | 0 | |
| ATF6 | r | 0.622 | 0.75 | 0.436 | 1 |
| ATF6 | p | 0.001* | 0.000* | 0.033* | 0 |
Table 4: Brain tissue correlation findings.
Correlations between study variables were analyzed using the Pearson correlation method. The r value represents the correlation coefficient and the p value represents the statistical significance value. p-values less than 0.05 were considered statistically significant.
Liver Tissue Western Blotting Results
GRP78 and PERK proteins were found to be statistically significantly higher in the diabetic group than in the control group (p<0.05), and were found to be statistically significantly lower in the diabetic antioxidant group than in the diabetic group (p<0.01). TWEAK and ATF6 proteins were found to be statistically significantly lower in the diabetic group than in the control group (p<0.001), and were found to be statistically significantly higher in the diabetic antioxidant group than in the diabetic group (p<0.01) (Figure 2).
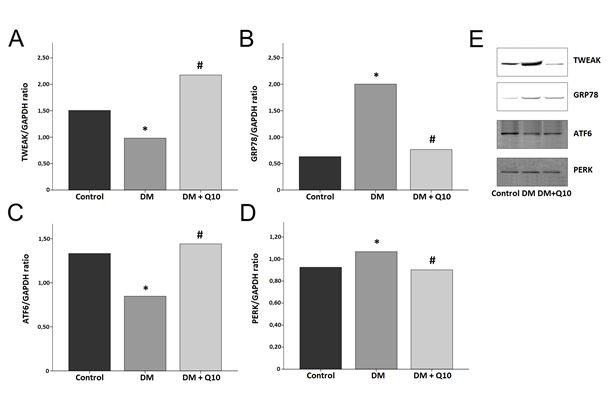
When we look at the correlational relationship between continuous variables of endoplasmic reticulum stress in liver tissue in our study, it is seen that there is a positive correlation between ATF6 and TWEAK (r=0.719, p=0.000), a positive correlation between PERK and GRP78 (r=0.588, p=0.003), and a negative correlation between ATF6 and GRP78 (r= -0.470, p=0.020) (Table 5). In addition, when we look at the correlation between oxidative stress parameters and endoplasmic reticulum stress parameters in the liver tissue, there is a negative correlation between GSH and GRP78 (r= -0.476, p=0.019), a negative correlation between GSH and PERK (r= -0.586, p=0.003) (Table 6).
| TWEAK | GRP78 | PERK | ATF6 | |||
|---|---|---|---|---|---|---|
| TWEAK | r | 1 | ||||
| TWEAK | p | 0 | ||||
| GRP78 | r | -0.221 | 1 | |||
| GRP78 | p | 0.298 | 0 | |||
| PERK | r | 0.027 | 0.588 | 1 | ||
| PERK | p | 0.899 | 0.003* | 0 | ||
| ATF6 | r | 0.719 | -0.47 | -0.028 | 1 | |
| ATF6 | p | 0.000* | 0.020* | 0.895 | 0 |
Table 5: Liver tissue correlation findings.
Correlations between study variables were analyzed using the Pearson correlation method. The r value represents the correlation coefficient and the p value represents the statistical significance value. p-values less than 0.05 were considered statistically significant.
| TWEAK | GRP78 | PERK | ATF6 | ||
|---|---|---|---|---|---|
| GSH | r | 0.151 | -0.476 | -0.586 | 0.343 |
| GSH | p | 0.482 | 0.019* | 0.003* | 0.101 |
| GSSG | r | -0.387 | -0.054 | -0.136 | -0.201 |
| GSSG | p | 0.062 | 0.802 | 0.527 | 0.347 |
| GSH / GSSG | r | 0.37 | -0.321 | -0.39 | 0.32 |
| rate | p | 0.075 | 0.127 | 0.06 | 0.127 |
Table 6: Liver tissue correlation findings (oxidative stress and endoplasmic reticulum stress).
Correlations between study variables were analyzed using the Pearson correlation method. The r value represents the correlation coefficient and the p value represents the statistical significance value. p-values less than 0.05 were considered statistically significant.
Discussion
Diabetes mellitus, one of the leading causes of death globally, is a chronic endocrine disease that causes hyperglycemia resulting from deficiencies in insulin production or activity, as well as disruptions in carbohydrate, fat, and protein metabolism [1, 2, 3]. Acute and chronic complications, which occur as a result of failure to control hyperglycemia in the advanced stages of diabetes, negatively affect the quality of life of patients and lead to serious organ failure. The most affected organs in diabetes are the pancreas, brain, kidney, and liver [1, 36].
Although many different mechanisms play a role in the pathogenesis of diabetes, oxidative stress is one of the conditions that have the most impact on it [4]. Recent studies suggest that endoplasmic reticulum stress has an important effect on the pathogenesis of diabetes too. There are studies showing that endoplasmic reticulum stress triggers diabetes, especially because it accelerates beta cell death, increases insulin deficiency, and causes insulin resistance [37, 38]. One of the major causes of endoplasmic reticulum stress is oxidative stress [24]. Oxidative stress, which develops with the increase of the effect of free radicals in the cell, may cause stress development by interacting with redox signal mediators that play a role in the protein folding process in the endoplasmic reticulum lumen [39, 40].
Antioxidant defense systems prevent damage caused by oxidative stress and endoplasmic reticulum stress. Antioxidant molecules are nucleophilic and reductive in nature and react with oxidants which in turn are usually electrophilic, giving them one or two electrons. Coenzyme Q10 acts as an antioxidant and as an indirect stabilizer of calcium channels in the prevention of lipid peroxidation and reduction of calcium overload [41, 42, 43, 44].
Based on this, we aimed to investigate whether hyperglycemia in liver and brain tissues triggered endoplasmic reticulum stress in rats by an experimental diabetes mellitus model and reveal whether coenzyme Q10 would have a preventing effect by suppressing oxidative stress. Another aim of our study was to examine the possible functions of the TWEAK molecule in the UPR signaling pathways and its relationship with endoplasmic reticulum stress.
Mohamed et al. administered 20 mg/kg of coenzyme Q10 to diabetic rats for 2 weeks and showed that coenzyme Q10 statistically significantly reduced blood glucose levels [45]. Similarly, in our study, we found that the glucose levels of the rats in the diabetic antioxidant group on the 42nd day of treatment were statistically significantly lower than the diabetic counterpart, same as seen on the 21st day of the treatment. Similar studies conducted by Fatemeh, et al. and Motawi, et al. revealed that coenzyme Q10 statistically significantly reduced glucose levels [46, 47]. These studies show that the glucose-lowering effect of coenzyme Q10 occurs by increasing the glucose utilization of cells and suppressing gluconeogenesis in the liver [45, 46, 47].
Glutathione is considered the most abundant molecule among the endogenous antioxidants, responsible for preventing the damage caused by oxidative stress in the body. The ratio between the oxidized (GSSG) and reduced (GSH) forms of the glutathione molecule is one of the best indicators of redox and it decreases under oxidative stress conditions [24, 25, 48].
Maheshwari et al. administered 10 mg/kg coenzyme Q10 to diabetic rats for 42 days. In this study, they showed that coenzyme Q10 statistically significantly reduced malondialdehyde (MDA) levels, which are oxidative stress indicators in the kidney tissue, and statistically significantly increased the levels of superoxide dismutase, catalase, and GSH levels, which are important antioxidants [32]. Another study showed that coenzyme Q10 treatment inhibited lipid peroxidation caused by hyperglycemia and decreased oxidative stress by increasing the activity of antioxidant enzymes such as SOD, catalase, and GSH [49]. Our results are parallel with the studies above.
In our study, we found a positive correlation between oxidative and endoplasmic reticulum stress markers, more significantly in the liver than in the brain. These findings support the thesis that there may be a relationship between oxidative and endoplasmic reticulum stress, as mentioned in the literature [12, 24, 39].
The endoplasmic reticulum is an important organelle in all eukaryotic cells that is responsible for the correct folding of proteins after transcription and reaching their target site [50]. Excessive accumulation of unfolded or misfolded proteins in the endoplasmic reticulum lumen causes endoplasmic reticulum stress. To correct this situation, a complex mechanism called UPR is initiated. The UPR mechanism is a homeostatic response to repair the disturbed balance between folded protein production and unfolded protein production. PERK, ATF6, and GRP78 proteins play an important role in the UPR mechanism [12, 13, 14].
Hengjiang, et al. determined that PERK, ATF6, and GRP78 levels in the kidney tissues of diabetic mice were statistically significantly higher than in the control group. In the same study, they showed that PERK, ATF6, and GRP78 levels in mice with astragaloside IV were statistically significantly lower than in diabetic mice. They also showed that the levels of caspase 3, caspase 9, and caspase 12, which are effective on the apoptotic pathway, were statistically significantly higher in mice in the diabetic group compared to those in the control group, and the levels of the same molecules in the antioxidant group were statistically significantly lower than in the diabetic group [51]. Another study conducted by Feng et al. showed that PERK, GRP78, and CHOP levels in the brain hippocampus tissues of diabetic rats were statistically significantly higher than in the control group; however, administration of vitamin D as an antioxidant statistically lowers these UPR markers [33]. There were also some studies to underscore that the increased levels of p-PERK, CHOP, and ATF6 in different tissues of diabetic rats, and the administration of variable antioxidants lowers the levels of these markers [52, 53].
In our study, ATF6 and GRP78 levels in the brain tissues of rats in the diabetic group were found to be statistically significantly higher than those in the control group. The PERK levels of the diabetic group were found to be higher than those of the control group, although it was not statistically significant. The PERK, ATF6, and GRP78 levels of the rats in diabetic antioxidant group were found to be statistically significantly lower compared to the diabetic counterpart. Our results are parallel with the aforementioned literature.
Consequently, we evaluated the PERK and GRP78 levels of the liver tissues. These UPR markers increased in the diabetic group as compared to the control group, however, the antioxidant molecule Q10 administration to the diabetic antioxidant group decreased the levels of PERK and GRP78 as compared to the diabetic group.
The UPR mechanism against endoplasmic reticulum stress is responsible for restoring homeostasis in the cell [1]. In cases where the UPR is not sufficient, the endoplasmic reticulum-mediated signal cascade in the cell is initiated and apoptosis is triggered. Cell death leads to loss of tissue function and may be the cause of various diseases related to endoplasmic reticulum stress. Apoptosis is thought to be the most important cause of pancreatic beta-cell loss, especially in type I diabetes patients [12, 13, 54].
TWEAK, which is claimed to have a proapoptotic effect, binds to its receptor called Fn14 under various pathological conditions and stimulates many mechanisms [55, 56]. Studies show that TWEAK/Fn14 receptor binding in the brain causes neuronal apoptosis in the pathogenesis of stroke and also contributes to an increase in the permeability of the blood- brain barrier in cerebral ischemia. In addition, the findings in these studies that show increased TWEAK and Fn14 expressions in the ischemic brain of stroke patients and in animal models of cerebral ischemia also support this opinion [57, 58]. The TWEAK/Fn14 interaction during cerebral ischemia activates the transcription factor NF-kβ pathway. This interaction also induces the expression of MMP-9, proinflammatory cytokines and chemokines, recruitment of neutrophils, disruption of the blood-brain barrier, and neurodegeneration. In light of these studies, a hypothesis has been proposed that claims TWEAK-Fn14 antagonists may limit brain damage [59, 60].
In our study, we aimed to reduce TWEAK-Fn14 binding in the brain tissues of diabetic rats with the coenzyme Q10 antioxidant. The findings revealed that the TWEAK levels of the diabetic antioxidant group were statistically significantly lower than those of the diabetic group. Although there are some studies suggesting a relationship between TWEAK and endoplasmic reticulum stress, none explains the whole mechanism in question (19). In our study, we examined the possible relationship between the TWEAK molecule with UPR signaling pathways which might explain their possible role in endoplasmic reticulum stress. Our results showed a statistically significant positive correlation between TWEAK and UPR proteins (ATF6, GRP78). We also observed a positive relationship between TWEAK and PERK. These findings may link the TWEAK molecule to endoplasmic reticulum stress by the UPR mechanism.
In the liver tissue there was a decline in the level of TWEAK molecules as compared to the control group. Tiller et al. claimed that there was no TWEAK receptor on adipocytes and mature liver cells, and therefore TWEAK could not affect these cells [61]. This claim supports our findings.
In consideration of the aforementioned factors, it can be concluded that the use of coenzyme Q10 for a certain period has a glucose-lowering effect. Coenzyme Q10 reduces oxidative stress and endoplasmic reticulum stress (especially in brain tissue) and increases antioxidant levels. Consequently, these findings suggest that coenzyme Q10 has a positive effect on reducing diabetic brain damage. In our study, the correlation between oxidative stress parameters and endoplasmic reticulum parameters supports the positive relationship between them. The relationship between TWEAK and endoplasmic reticulum stress has not been fully illuminated, but the finding of a positive correlation in our study may lead to a new way of addressing this issue.
Acknowledgments
This study is supported by the research fund of University Istanbul-Cerrahpasa (Project number: 30528).
References
-
Van der Kallen CJH, Van Greevenbroek MMJ, Stehouwer CDA, Schalkwijk CG (2009) Endoplasmic reticulum- stress-induced apoptosis in the development of diabetes: is there a role for adipose tissue and liver? Apoptosis 14: 1424-1434.
-
Haziran (2015) TUE Physician Guidelines Medical Information to Support the Decisions of TUE Committees, Diabetes Mellitus.
-
IDF Diabetes Atlas, Eighth edition, 2017.
-
Ighodaro OM (2018) Molecular pathways associated with oxidative stress in diabetes mellitus. Biomed Pharmacother 108: 656-662.
-
Gomes BF, Accardo CM (2019) Immunoinflammatory mediators in the pathogenesis of diabetes mellitus. Einstein (Sao Paulo) 17(1): 1-5.
-
Demirtas L, Guclu A, Erdur FM, Akbas EM, Ozcicek A, et al. (2016) Apoptosis, autophagy & endoplasmic reticulum stress in diabetes mellitus. Indian J Med Res 144(4): 515-524.
-
Maamoun H, Abdelsalam SS, Zeidan A, Korashy HM, Agouni A (2019) Endoplasmic Reticulum Stress: A Critical Molecular Driver of Endothelial Dysfunction and Cardiovascular Disturbances Associated with Diabetes. Int. J. Mol. Sci 20(7): 1658.
-
Ghosh R, Negron KC, Papa FR (2019) Endoplasmic reticulum stress, degeneration of pancreatic islet b-cells, and therapeutic modulation of the unfolded protein response in diabetes. Mol Metab 27(suppl): S60-S68.
-
Engin F (2016) ER stress and Development of Type 1 Diabetes. J Investig Med 64(1): 2-6.
-
Papa FR (2012) Endoplasmic reticulum stress, pancreatic β-cell degeneration, and diabetes. Cold Spring Harb Perspect Med 2(9): 1-18.
-
Oakes SA, Papa FR (2014) The role of endoplasmic reticulum stress in human pathology. Annu Rev Pathol 10: 173-194.
-
Hasnain SZ, Prins JB, McGuckin MA (2016) Oxidative and endoplasmic reticulum stress in β-cell dysfunction in diabetes. J Mol Endocrinol 56(2): R33- R54.
-
Hampton RY (2002) ER-associated degradation in protein quality control and cellular regulation. Curr Opin Cell Biol 14(4): 476-482.
-
Gardner BM, Walter P (2011) Unfolded proteins are IRE1-activating ligands that directly induce the unfolded protein response. Science 333(6051): 1891-1894.
-
Gardner BM, Pincus D, Gotthardt K, Gallagher CM, Walter P (2013) Endoplasmic reticulum stress sensing in the unfolded protein response. Cold Spring Harb Perspect Biol 5(3): 1-15.
-
Tomita T (2017) Apoptosis of pancreatic β-cells in type 1 diabetes. Bosn J Basic Med Sci 17(3): 183-193.
-
Tomita T (2016) Apoptosis in pancreatic β-islet cells in type 2 diabetes. Bosn J Basic Med Sci 16(3): 162-179.
-
Boulamery A, Jego SD (2017) Regulation of neuroinflammation: What role for the tumor necrosis factor-like weak inducer of apoptosis/fn14 pathway? Immunology 8: 1534-1540.
-
Vendrell J, Chacon MR (2013) TWEAK: A new player in obesity and diabetes. İmmunology 4(498): 1-6.
-
Yildirim ZK, Sumnu A, Bademler N, Kilic E, Sumnu G, et al (2016) Soluble TNF-like weak inducer of apoptosis as a new marker in preeclampsia: A pilot clinical study. Dis Markers, pp: 5930589.
-
Tibaut M, Petrovic D (2016) Oxidative stress genes, antioxidants and coronary artery disease in type 2 diabetes mellitus. Cardiovasc Hematoll Agents Med Chem 14(1): 23-38.
-
Gerber PA, Rutter GA (2017) The Role of oxidative stress and hypoxia in pancreatic beta-cell dysfunction in diabetes mellitus. Antioxid. Redox Signal 26(10): 501- 518.
-
Keane KN, Cruzat VF, Carlessi R, Bittencourt PIH, Newsholme P (2015) Molecular events linking oxidative stress and inflammation to insulin resistance and 𝛽-cell dysfunction. Oxid Med Cell Longev 2015:181643.
-
Zeeshan HMA, Lee GH, Kim HR, Chae HJ (2016) Endoplasmic reticulum stress and associated ROS. Int J Mol Sci 17(327): 1-20.
-
Diez CE, Miguel V, Mennerich D, Kietzmann T, Perez PS, et al. (2015) Antioxidant responses and cellular adjustments to oxidative stress. Redox Biol 6: 183-197.
-
Shen Q, Pierce JD (2015) Supplementation of coenzyme Q10 among patients with type 2 diabetes mellitus. Healthcare 3(2): 296-309.
-
Sourris KC, Harcourt BE, Tang PH, Morley AL, Huynh K, et al. (2012) Ubiquinone (coenzyme Q10) prevents renal mitochondrial dysfunction in an experimental model of type 2 diabetes. Free Radic Biol Med 52(3): 716-723.
-
Genova ML, Lenaz G (2011) New Developments on the functions of coenzyme Q in mitochondria. Biofactors 37(5): 330-354.
-
Alam A, Rahman M (2014) Mitochondrial dysfunction in obesity: Potential benefit and mechanism of Co-enzyme Q10 supplementation in metabolic syndrome. J Diabetes Metab Disord; 13(60): 1-11.
-
Fazakerley DJ, Chaudhuri R, Yang P, Maghzal GJ, Thomas KC, et al. (2018) Mitochondrial CoQ deficiency is a common driver of mitochondrial oxidants and insulin resistance.
-
Sonmez H, Syuer S, Gungor Z, Kokoglu E, Isbir T (1997) Sialidase activities and sialic acid levels of the liver and serum from streptozotocin induced diabetic rats. Diabetes Research 32: 101-106.
-
Maheshwari R, Balaraman R, Kumar Sen A, Shukla D, Seth A (2017) Effect of concomitant of coenzyme Q10 with sitagliptin on experimentally induced diabetic nephropathy in rats. Ren Fail 39(1): 130-139.
-
Feng G, Yue H, Wang L, Ding C, Wu L, et al. (2017) Vitamin D supplement ameliorates hippocampal metabolism in diabetic rats. Biochem Biophys Res Commun 490(2): 239-246.
-
Afrin R, Arumugam S, Soetikno V, Thandavarayan RA, Pitchaimani V, et al. (2015) Curcumin ameliorates streptozotocin-induced liver damage through modulation of endoplasmic reticulum stress-mediated apoptosis in diabetic rats. Free Radic Res 49(3): 279- 289.
-
Marie FM, Lucie P, Wurood AH, Tobin G, Daniel G (2019) Radiation induces changes in toll-like receptors of the uterine cervix of the rat. Plos One 14(4): 1-13.
-
Skyler JS, Bakris GL, Bonifacio E, Darsow T, Eckel RH, et al. (2017) Differentiation of diabetes by pathophysiology, natural history, and prognosis. Diabetes; 66(2): 241-255.
-
Clark AL, Urano F (2016) Endoplasmic reticulum stress in beta cells and autoimmune diabetes. Curr Opin Immunol 43: 60-66.
-
Kim MK, Kim HS, Lee IK, Park KG (2012) Endoplasmic reticulum stress and insulin biosynthesis: A review. Exp Diabetes Res 2012: 509437.
-
Malhotra JD, Kaufman RJ (2007) Endoplasmic reticulum stress and oxidative stress: A vicious cycle or a double- edged sword? Antioxid. Redox Signal 9(12): 2277-2293.
-
Moron EB, Jimenez ZA, Maranon A, Iannantuoni F, Lopez IE, et al. (2019) Relationship between oxidative stress, ER stress, and inflammation in type 2 diabetes: The battle continues. J Clin Med 8: 1-22.
-
Bonakdar RA, Guarneri E (2005) Coenzyme Q10. Am Fam Physician; 72(6): 1065-1070.
-
Raziner AE (2019) Coenzyme Q10. Methodist Debakey Cardiovasc J 15(3): 185-191.
-
Camacho JDH, Bernier M, Lluch GL, Navas P (2018) Coenzyme Q10 supplementation in aging and disease. Front Physiol 9: 1-11.
-
Acosta MJ, Fonseca LV, Desbats MA, Cerqua C, Zordan R, et al. (2016) Coenzyme Q biosynthesis in health and disease. Biochim Biophys Acta 1857(8): 1079-1085.
-
Amin MM, Asaad GF, Abdel Salam RM, El-Abhar HS, Arbid MS (2014) Novel CoQ10 antidiabetic mechanisms underlie its positive effect: Modulation of insulin and adiponectine receptors, tyrosine kinase, PI3K, glucose transporters, sRAGE and visfatin in insulin resistant/ diabetic rats. Plos one 9(2): 1-12.
-
Samimi F, Baazm M, Eftekhar E, Rajabi S, Goodarzi MT, et al. (2019) Possible antioxidant mechanism of coenzyme Q10 in diabetes: Impact on Sirt1/Nrf2 signaling pathways. Res Pharma Sci 14(6): 524-533.
-
Motawi TK, Darwish HA, Hamed MA, El-Rigal NS, Naser AFA (2017) A therapeutic insight of niacin and coenzyme Q10 against diabetic encephalopathy in rats. Mol Neurobiol 54(3): 1601-1611.
-
Lu SC (2013) Glutathione synthesis. Biochim Biophys Acta 1830 (5): 3143-3153.
-
Modi K, Santani DD, Goyal RK, Bhatt PA (2006) Effect of coenzyme Q10 on catalase activity and other antioxidant parameters in streptozotocin-induced diabetic rats. Biol Trace Elem Res 109(1): 25-33.
-
Ruggiano A, Foresti O, Carvalho P (2014) ER-associated degradation: Protein quality control and beyond. J Cell Biol 204(6): 869-879.
-
Hengjiang G, Cao A, Chu S, Wang Y, Zang Y, et al. (2016) IV attenuates podocyte apoptosis mediated by endoplasmic reticulum stress through upregulating sarco/endoplasmic reticulum Ca2+-ATPase 2 expression in diabetic nephropathy. Front Pharmacol 7: 1-18.
-
Shuqin S, Yang S, Ana N, Wanga G, Xua Q, et al. (2019) Astragalus polysaccharides inhibits cardiomyocyte apoptosis during diabetic cardiomyopathy via the endoplasmic reticulum stress pathway. Journal of Ethnopharmacology 238: 1-12.
-
Tian N, Gao Y, Wang X, Wu X, Zou D, et al. (2018) Emodin mitigates podocytes apoptosis induced by endoplasmic reticulum stress through the inhibition of the PERK pathway in diabetic nephropathy. Drug Des, Devel Ther 12: 2195-2211.
-
Marasco MR, Linnemann AK β-cell autophagy in diabetes pathogenesis. Endocrinology 159(5): 2127-2141.
-
Laso VF, Sastre C, Barbero NM, Egido J, Ventura JLM, et al. (2017) TWEAK blockade decreases atherosclerotic lesion size and progression through suppression of STAT1 signaling in diabetic mice. Scientific Reports 7: 1-15.
-
Sanz AB, Nino MDS, Ortiz A (2011) TWEAK, a multifunctional cytokine in kidney injury. Kidney International 80(7): 708-718.
-
Inta I, Frauenknecht K, Dorr H, Kohlhof P, Rabsilber T, et al. (2008) Induction of the cytokine TWEAK and its receptor Fn14 in ischemic stroke. J Neurol Sci 275(1-2): 117-120.
-
Potrovita I, Zhang W, Burkly L, Hahm K, Lincecum J, et al. (2004) Tumor necrosis factor-like weak inducer of apoptosis-induced neurodegeneration. J Neurosci 24(38): 8237-8244.
-
Polavarapu R, Gongora MC, Winkles JA, Yepes M (2005) Tumor necrosis factor-like weak inducer of apoptosis increases the permeability of the neurovascular unit through nuclear factor-κb pathway activation. J Neurosci 25(44): 10094-10100.
-
Haile WB, Echeverry R, Wu J, Yepes M (2010) The interaction between tumor necrosis factor-like weak inducer of apoptosis and its receptor fibroblast growth factor-inducible 14 promotes the recruitment of neutrophils into the ischemic brain. J Cereb Blood Flow Metab 30(6): 1147-1156.
-
Tiller G, Posovszky PF, Laumen H, Finck A, Skurk T, et al. (2009) Effects of TWEAK (TNF superfamily member 12) on differentiation, metabolism, and secretory function of human primary preadipocytes and adipocytes. Endocrinology 150(12): 5373-5383.
- Superposition of Cryo-EM and AlphaFold Predictions of Dengue Antigen-Antibody Complexes
- Jugular-Applied Coherent Low-Level Laser Therapy Enhances Systemic Mitochondrial Metabolic Function and Antioxidant Response
- Role of OMC32 Polypeptide in Acrosin-Mediated Exocytosis during the Bovine Sperm Acrosome Reaction
- Association of Galectin-3 but not Laminin in Tamoxifen-Induced Growth Suppression in Breast Cancer MCF-7 Cells
- Effect of Different Wavelengths of Light on the Rate of Photosynthesis
- Nutritional, Therapeutic, and Environmental Effect of Oyster Mushrooms: An Editorial