Osmoregulatory Plasticity in Oreochromis niloticus-Nile Tilapia, and it’s Possible Estuary Use for Rivers Dispersion
Nile tilapia is a widely farmed species globally, due to its resistance to environmental challenges. Additionally, it is a generalist consumer and a prolific breeder. In Brazil, this fish is recognized as an invasive species because of its high dispersion and establishment rates, which compromise native biota. Estuarine waters (0.06 - 32‰) are being used in tilapia aquaculture, which is a promisor fish farming for euryahalinity species. The tilapia’s euryhalinity is well-know, but most protocols are based on gradual exposure. This research showed the tolerance of Nile tilapia on abrupt exposure to 0‰ (FW), and 10, 20, 30, 35 or 40‰, over in 6, 12 and 24 hours. An abrupt exposure resistance demonstrate that this fish can use estuaries to disperse for other watersheds. After experimental design the blood sample was taken after anesthesia was administered. Subsequently, fish was euthanized by spinal section, for muscle and gills samples collections. All samples were keep freezing until ionic, dehydration and enzyme measurement. Nile tilapia survived 24h until 30‰, 12h at 35‰ and 9h at 40‰. Osmolality increase and muscle dehydration decrease both above 20‰. While sodium levels keep regulated in all salinities, chloride, potassium and magnesium levels also increased above 20‰. These results prove that tilapia can easily tolerate abrupt exposure until 20‰ without an osmotic stressor. Gill carbonic anhydrase (C.A.) activity decreased at 30‰, and NKA did not change along salinities and times. These results complements plasma and dehydration averages, showing the large tolerance of this specie to salinity challenges, even in abrupt exposure. Then, these results can be used for discuss about the estuary fish farming system for tilapia, as this species may use estuarine waters to nearby rivers and also can harm the native ecosystem.
Introduction
Brazilian fish farming is the mainly activity through which non-native species can disperse into natural environment (about 87% involves non-native species) [1]. Tilapias and carps are the most commonly cultivated exotic species [2]. This high percentage can be linked with the economic benefits; since tilapia’s meat has many organoleptic characteristics which promotes high demand for the human diet add the low-cost in its farming [3].
Tilapia is considered an invasive species in several countries, as Brazil, because can tolerate several abiotic stressors parameters, not tolerated by most fishes, including native species. In addition, is an omnivorous species, has a short reproductive cycle (each 6 months), high offspring (200-1000 eggs per spawn), sexual reversion ability and good growth and development even under farming compared with other fishes. The physiological traits of Nile tilapia promote its farming in net tanks systems, also in estuary water (between freshwater – 0 to 0.06‰ and marine water - ~33‰). However, these systems combine with species’ high tolerance, could increase the risk of its invasion into other watershed, potentially leading a local ecological imbalance [4].
Nile tilapia belong to Cichlidae family (with freshwater secondary occupation) and Perciformes order (with marine ancestors) has an evolutionary history that helps explain, at least in part, the tilapia euryhalinity. Then, the specie can tolerate salinity changes because its genome retains osmoregulatory plasticity, allowing that different genes can expressed proteins for both active salt absorption (in freshwater) or secretion (in salt water) [5].
The blood osmolality and ionic levels evaluation, with muscle dehydration experiment, can offer insights into the euryhalinity capability perspective of species like Nile tilapia, as all fishes are osmoregulators. This means they must spend energy on ion transporters, to maintain internal ion levels with an opposite concentration of external environment [6]. Gill C.A. (carbonic anhydrase) and NKA (Na+/K+-ATPase) enzyme activities can reinforce these regulatory mechanisms at the molecular level over the long-term, as osmoregulatory proteins may be up or downregulated, or even not expressed [7].
The CCs cells represents approximately 10% of the gill epithelium, and it can translocate primary or secondary osmoregulatory proteins, as like C.A. and NKA to their plasma membranes, or retain them in intracellular space. The C.A. is a cytoplasmic enzyme that catalyzes the dehydration carbonic acid, producing protons and bicarbonate. While chemical gradient of protons (H+ efflux promote by H+- ATPase) allow the sodium passive uptake, the bicarbonate/ chloride cotransporter increase intracellular chloride levels. This is crucial for to the function of basolateral NKA, with sodium internalization to blood. Freshwater osmoregulators must be able to salt absorption through this mechanism. To other hand, seawater osmoregulation involves a sodium paracellular pathway, allowed by potassium recirculation in the basolateral membrane [8].
This research examined whether Nile tilapia can tolerate abrupt salinity changes, and this repercussion on balance of native ecosystems.
Methodology
Committee of Ethica approved the experimental design (process number: 23075.081084/2011-67). Nile tilapia (9- 13 cm, n= 162) were acclimated for 5 days and submitted to 6, 12 and 24 hours at 0, 10 and 20‰; 6 and 12 hours at 30 and 35‰; and 6 hours at 40‰ (according preliminary survive tests).
After experimental design, all fish were anesthetized with benzocaine (prepared with 120 mg/L in ethanol, with 1:500 of water dilution, until movement lost ~3 min) for blood sample collect (vein punction, with heparin). The osmolality was measured with a micro-osmometer, and sodium and potassium levels by a flame photometer, while chloride and magnesium were determined via spectrophotometry.
After euthanasia by spinal section, a piece of muscle and the second left gill arch were collected for muscle water content or enzyme activities evaluation. Muscles samples were dried in an oven for calculate the dehydration: [(Wh - Wd)/ Wh] x 100, where Wh refers to wet muscle weight and Wd the dried muscle. For C.A. assay, the gill tissue was homogenized (1:10) in a buffer solution (pH of 7,4) containing mannitol (225mM), sucrose (75mM) and tri-phosphate (10mM), with subsequently centrifugation. The buffer(control) or the buffer with samples were put in a glass. Then, the decrease of the pH was obtained soon after the addition of carbon dioxide in water, also put on glass solution. Then, the pH was noted every 4 seconds, five times.
For NKA assay, gills were homogenized with 200µl of buffer solution cointaining sucrose (250mM), imidazole (50mM) and Na2EDTA (10 mM), along with 10µl of EDTA diluted in buffer, for centrifugation. The enzyme activity is measured by comparing ATP catalyze in the presence or absence of ouabain (NKA inhibitor), using absorbance readings taken over 10 minutes at 340nm.
Statistical analyzes were performing using one way – ANOVA (salinity x time, in samples of 0-30‰). T-test was used to compare time-dependent data. Significance level set at 5%.
Results and Discussion
The half-lethal salinity was achieve over 9h at 30‰ and over 6h at 35‰ and 40‰ salinity. Other studies have shown that the euryhaline Mozambique tilapia, O. mossambicus tolerated 6 hours at 30‰9 and 4 hours at seawater [9, 10]. As estuarine waters has 0.06 to 32‰, and a tidal cycle occurs approximately every 12 hours per day, these tilapia tolerance provides substantial insights into tilapia potential to invade other watersheds via estuaries. The estuary fish farming is increasing in several Brazilian states [2], creating an important concern about this system using tilapias.
Plasma osmolality in freshwater was measured at 252 mOsm/ kg of water, reaching over of 400 mOsm/ kg of water at salinities above 20%. At this salinity, osmolality in 24 hours was lower than 12 hours, showing a rapid osmolality adjustment (Figures 1A-C). In other fishes, osmolality also increases during abrupt exposure to 20‰ salinities, but normal (control) osmolality returns only after several weeks [11, 12, 13]. Muscle water content decreased at 20‰ in 24 hours (Figures 1D-F). This manner, osmotic muscle regulation closely follows osmolality, at least at 20‰, salinity threshold for mortality. Some studies showed a faster dehydration time (within 6 hours) at the same salinity 16.
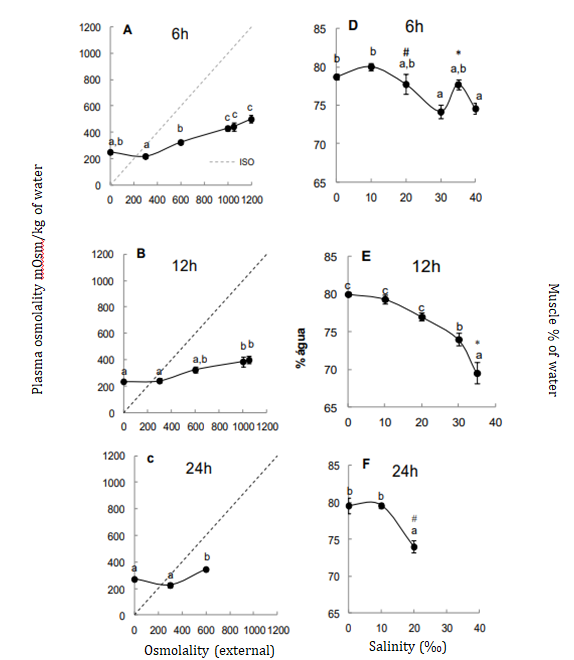
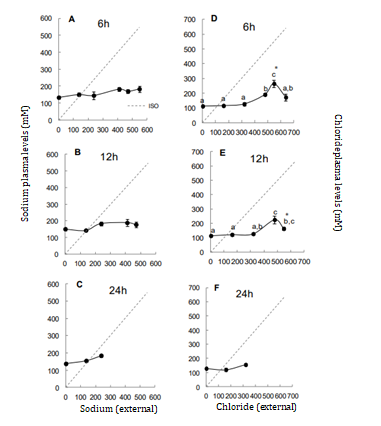
Sodium levels remained regulated across all times and salinities, averaging ±148 mM, which demonstrate sodium’s essential role in maintaining other cellular activities. The highest chloride levels increased above 20‰ reinforcing that higher salinities, over time, acting as stressors to the fish, limiting their survival tolerance (Figure 2). Osmolality and chloride levels serve as indicators of osmotic stress, reflecting the need for a rapid modulatory response that not all euryhaline species can achieve during abrupt exposure [14]. Addittionally, salinities changes are related to the number or density CCs, which likely influence in chloride influx [15].
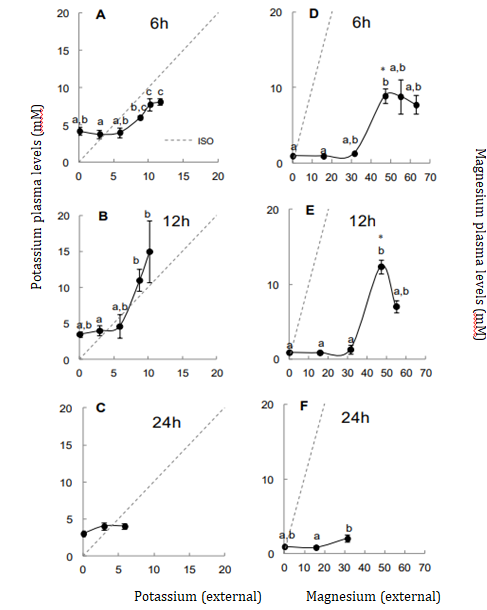
Potassium and magnesium levels also increased above 20% (Figure 3). The elevated potassium levels may indicate cell lysis, which reinforce the ionic-osmotic deregulation observed. Tilapias exposed to abrupt salinities changes showed increases in these ions even at 18‰ [16].
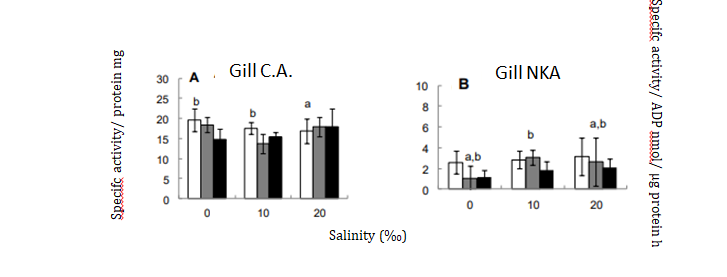
Osmotic stress promote deregulation in osmoregulatory proteins functions, as gene expression regulation is a long- term process (i.e.; weeks or months) and relies on the “reserves” of these molecules. Gills C.A. activity decreased just after 24h at 20‰, while NKA keep regulated (Figure 4). While NKA maintain membrane potential (for cell survival) C.A. activity is essential mainly in freshwater, due it salt absorption role, but is also crucial for maintaining acid-base balance, so its abrupt reduction may introduce additional stress factors for the fish [17]. Other studies showed that both in Nile tilapia 10 and Mozambique tilapia [13], typically show increased osmoregulatory enzyme activity as result as a pre-adaption to higher salinities. Mozambique tilapia also shows enzyme activity decrease at seawater, indicating fish limits of salinity stress [12].
This research proved that the tilapia is extremely euryhaline even under abrupt exposure of salinities changes, tolerating a tidal cycle without difficulty. This manner, this species really has the potential to invade, disperse and establish itself in other watersheds through estuaries salinities. This findings warning about new fish farming in estuaries, which could increase the density and genic flow of this invasive species. The higher density of invasive species can hybridize with native species, which results in the loss of genetic patrimony of the ecosystem, in addition with changes in trophic chain and new parasites bring by the exotic fish species, that can lead native biodiversity lost and even extinctions. Even with these issues, several governmental programs and the Law 5989/09 stimulates the use of exotic species for national fish farming [18].
Net tanks have a higher human intervention, observed mainly in fishing, which is made nearby or directly in natural water’s body. Is common to find not-cultivated species in net tanks, which suggest the opposite way for farming species, easing the invasive specie dispersion [19].
Conclusion
- Nile tilapia exhibits high euryahalinity, even under abrupt exposure to salinities changes;
- The physiological performance of Nile tilapia in the short-term (ionic and osmotic regulation) demonstrates that it can survive in estuarine waters long enough to spread to other watersheds;
- Nile tilapia used in this research showed greater tolerance compared to other studies with Nile tilapia more euryhaline tilapia species. These results may be related to hybrization species in fish farming;
- Abrupt exposure does not allows for observation of gene regulation of osmoregulatory proteins, indicating that abrupt salinity changes do not enable the species to mount a long-term physiological response;
- Other suggestions of studies can have an ecological approach, for example, comparing native or non-native species diversity and richness in several river watersheds nearly estuary fish farming.
References
-
Gu DG, Wang JW, Xu M, Mu XD, Wei H, et al. (2022) Does aquaculture aggravate exotic fish invasions in the rivers of southern China? Aquaculture 547: 737492.
-
Valenti WC, Barros HP, Moraes VP, Bueno GB, Cavalli RO (2021) Aquaculture in Brazil: past, presente and future. Aquaculture Reports 19.
-
Banha F, Gago J, Margalejo D, Feijão J, Casals F, et al. (2024) Angler’s preferences, perceptions and practices regarding non-native freshwater fish. Reviews in Fish Biology and Fisheries 34: 385-404.
-
Lomeu EAC, Valle AA, Azevedo JS (2024) Fishing Grounds and the Invasive Species Impact: a Case Study in a Coastal Brazilian Watershed, Braz arch biol Technol.
-
Pan J, Chen L, Ji Y, Huang Y, Bu X, et al. (2023) A crucial role in osmoregulation against hyperosmotic stress: Carbohydrate and inositol metabolism in Nile tilapia (Oreochromis niloticus). Aquaculture reports.
-
Seal AP, Breves JP (2022) Endocrine and osmoregulatory responses to tidally-changing salinities in fishes. General and Comparative Endocrinology 326 :114071.
-
Kültz D, Gilmour KM (2020) Iono-and osmoregulation. In: Suzanne C, Evans DH (Eds.), The Physiology of Fishes. 5th (Edn.), Boca Raton: CRC Press, pp: 256.
-
Mohamed NA, Saad M, Shukry M, et al. (2021) Physiological and ion changes of Nile tilapia (Oreochromis niloticus) under the effect of salinity stress. Aquaculture Reports.
-
Weng CF, Chiang CC, Gong, HY, Chen MC, Lin CJF, et al. (2002) Acute changes in gill Na+-K+-ATPase and creatine kinase in response to salinity changes in the euryhaline teleost, tilapia (Oreochromis mossambicus). Physiological and Biochemical Zoology 75(1): 29-36.
-
Güner Y, Ozden O, Çagirgan H, Altunok M, Kizak V (2005) Effects of salinity on the osmoregulatory functions of the gills in Nile Tilapia (Oreochromis niloticus). Turkey Journal of Veterinary and Animal Science 29: 1259-1266.
-
Souza BLR, Freire CA (2009) The Handling of salt by the neotropical cultured freshwater catfish Rhamdia quelen. Aquaculture 289: 167-174.
-
Wang PJ, Lin CH, Hwang LY, Huang CH, Lee TH, et al. (2009) Differential responses in gills of euryhaline tilapia, Oreochromis mossambicus, to various hyperosmotic shocks. Comparative Biochemistry and Physiology, Part A 152(4): 544-551.
-
Kammerer BD, Cech JJ, Kultz D (2010) Rapid changes in plasma cortisol, osmolality, and respiration in response to salinity stress in tilapia (Oreochromis mossambicus). Comparative Biochemistry and Physiology, Part A 157(3): 260-265.
-
Gutierre SMM (2011) Ferramentas fisiológicas para avaliação do potencial invasor de peixes dulcícolas. (Master degree defense) Universidade Federal do Paraná, pp: 73.
-
Prodocimo V, Souza CF, Pessini C, Fernandes LC, Freire CA (2008) Metabolic substrates are not mobilized from the osmoregulatory organs (gills and kidney) of the estuarine pufferfishes Sphoeroides greeleyi and S. testudineus upon shortterm salinity reduction. Neotropical Ichthyology 6(4): 613-620.
-
Ouattara NG, Bodinier C, Nègre SG, Cotta HD, D’Cotta H, et al. (2009) Changes in gill ionocyte morphology and function following transfer from fresh to hypersaline waters in the tilapia Sarotherodon melanotheron. Aquaculture 290: 155-164.
-
Karsi A, Yavuzcan YH (2005) Secondary stress response of Nile tilapia, Oreochromis niloticus, after direct transfer to different salinities. Tarim Bilimleri Dergisi 11: 139-141.
-
Silva DLAC (2012) Análise de Investimento em piscicultura: produção de tilápia do Nilo (Oreochromis niloticus) em tanques-rede. Apresentação oral. SOBER - Sociedade Brasileira de Economia, Administração e Sociologia Rural. Grupo de Pesquisa: Agropecuária, Meio Ambiente e Desenvolvimento Sustentável.
-
Ayroza LMS (2009) Criação de tilápias do Nilo, Oreochromis niloticus, em tanques-rede, na usina hidrelétrica de Chavantes, Rio Paranapanema, SP/PR. (Thesis). Universidade Estadual Paulista, pp: 104.
- Genetic Improvement of Nile Tilapia (Oreochromis niloticus): Advances in Selective Breeding and Genomic Approaches for Sustainable Aquaculture
- Microplastics, Contaminants, and Waste Hotspots: Divergences and Faults in Prioritizing Control Efforts
- Creating a Healthier, More Vibrant Open and Closed Aquatic Environment. A Submersible, Centrifugal Magnetically Affixed Current Changing Aquarium Pump
- An Attempt to Assess Alpha Diversity and Sample Size: Using the Ostracod Assemblages off Kumamoto Port, Japan
- Assessment of the Efficiency of Common Fishing Gears and Crafts Used at Mohananda River of Chapai Nawabganj, Bangladesh
- Fish Productivity and Biodiversity Status of Sundarban Mangrove in Bangladesh