Microbial Fertilizer Improves Constitutive and Induced Resistance of Tomato against a Generalist Insect
Plants express various constitutive and induced resistance mechanisms that require substantial share of plant nutrients. Microbial fertilizers such as Phosphate Solubilizing Bacteria (PSB) are known to enhance nutrient availability to the plant, thereby helping in plant growth. However, it is still unknown whether such a growth compromises plant resistance. In present study, we evaluated the effect of PSB on early as well late responses in tomato (Solanum lycopersicum) after damage by Spodoptera litura. We also studied whether these responses translate to resistance mechanisms by analyzing response of insect larvae on damaged plants. Our results indicate that, at early hour both unsupplemented and plants supplemented by PSB showed equal susceptibility towards insect induced oxidative damage. From 48h onwards elevated resistance mechanisms were observed in PSB-supplemented plants, especially increase in trichome numbers and shoot biomass. Moreover, decreased feeding preference of larvae was also observed on the previously damaged PSB-supplemented plants. However, an increased consumption index does indicate increased palatability of the plant, possibly due to high phosphorus availability. Our results thus indicate that PSB administration improves growth as well as induced resistance in tomato.
Introduction
Plants encounter variety of insects in their natural environment, for which they have developed constitutive and induced defence system. Constitutive defenses are not as common in annual plants and where, there are small and unpredictable insect communities, due to their high cost [1, 2]. In effect, these plants rely on inducible defenses as a cost saving strategy [1, 3]. However, even inducible defenses may result in fitness costs such as decreased fruit production [4]. A major drawback of such defenses is that the plant remains susceptible in the long period between the initial damage and the peak of defense response [3]. Further, various environmental features are considered to affect allocation of plant resources to defensive compounds [5, 6]. In view of these evidences, nutrient accessibility becomes a vital factor in influencing the allocation of the available resources and hence the plant growth [5, 7]. In agricultural systems application of fertilizers is considered as a fundamental method of improving soil nutrient availability for plants [7, 8, 9, 10].
Phosphorus is one of the most essential macronutrients for plant growth and development [11]. However, in soil, most of it is present in bound form and thus is a major limitation for plant growth [12]. A large portion of soluble inorganic phosphate that is applied to soil as chemical fertilizer is immobilized after application, or may runoff and thus becomes unavailable to plants [13, 14]. A promising alternate to synthetic fertilizers is the use of Plant Growth Promoting Microorganisms (PGPMs) such as phosphate solubilizing bacteria (PSB). PSB play a central role in mineralization of most organic phosphorous compounds using phosphatase enzymes from inorganic and organic pools of the total soil phosphorus [15, 16]. However, they secrete organic acids such as acid phosphate, gluconic acid and malic acid, which affect plant’s physiology, growth and yield [17, 20]. Several reports also suggest that due to their presence in the plant rhizosphere, PGPMs might also help in inducing plant’s resistance to phytopathogens insect pests and nematode pests [17, 21, 22, 23, 24].
Tomato (Solanum lycopersicum L.) is the second most important vegetable crop in the world. It occupies about 3.7 million hectares agriculture area worldwide that produces approximately 100 million tons fresh fruit annually [25]. However, despite its global importance in diet, very few studies have dealt with the factors that affect its production in areas that are prone to stress conditions such as drought, salinity, nutrient deficiency, weeds and insect pests [26, 27]. Many studies suggest improved health and immunity in tomato when supplemented with PGPMs due to the increased availability of primary nutrients [28, 29, 30]. However, their effect on plant resistance mechanism is still not known. Therefore in present study, we evaluated the effect of PSB on the constitutive and induced resistance mechanisms in tomato during damage by Spodoptera litura. Biochemical (antioxidants and chlorophyll), morphological (trichomes) and physiological (dry weight) parameters were studied in plants with and without PSB supplementation. Furthermore, preference as well as feeding and performance of S. litura was also examined on damaged and undamaged plants treated with or without PSB in order to determine whether these responses may be translated as resistance mechanisms.
Materials and Methods
Plant
Seeds of S. lycopersicum var. PH-4 were obtained from Indian Agricultural Research Institute (IARI), New Delhi, India. The seeds were initially germinated on glass beads in 10 x10cm plastic containers inside the plant growth chamber at 27±1ºC, 65±5% RH and 16: 8h light/dark photoperiod. The plants of stage 11 as per BBCH scale Lancashire, et al. [31] were transferred to 6 inch earthen containing garden soil with vermicompost in 3:1 ratio inside the insect free enclosure at Sri Venkateswara College. For plant growth in pots, 1 Kg of soil was mixed with 10ml of a commercial PSB formulation Phosphophix® (International Panaacea Pvt. Ltd, New Delhi, India) containing Bacillus megaterium, Pseudomonas fluorescence and Bacillus spp. Equal number of control plants was given soil mixed with 10ml of double distilled water instead of PSB formulation.
Insect
S. litura was obtained from laboratory cultures maintained in Animal Plant Interactions lab at Sri Venkateswara College, University of Delhi. The insects were periodically supplemented with field collected population to avoid inbreeding. Insects were maintained at a constant temperature of 27±1ºC with 75±5% RH and 16:8h light/dark photoperiod on castor leaves sterilized with K2MnO4. The adults were provided with 10% honey solution with multivitamins.
Induction of Plants
The experiments were conducted on plant stage 25 according to BBCH scale. Single third instar S. litura was introduced in a 2.5cm plastic clip cage on the third true leaf of the plant. Undamaged treatments of PSB supplemented and control plants received empty clip cages on its third true leaf for the undamaged treatment set. The damaged and undamaged plants were arranged randomly in the insect free enclosure. Insects were allowed to feed on the plant for 48h, unless mentioned otherwise. Based on the preliminary Rhodamine B dye tracking study, fifth leaf was used for the study responses. Following induction, plants were studied for morphological, physiological and biochemical parameters at various time points.
Trichome Number
Glandular and non-glandular trichomes were counted on the adaxial and abaxial surface of the leaf according to O’Neal, et al. [32]. Two spots of 1cm2 each were used to count the trichomes and their mean values were used for analysis.
Fresh and Dry Weight
Fresh and dry weight study was done on plants of stage 41 according to BBCH scale in a set of 20 replicates. Insect induction of each set of the four sets of plant was done for 48h which was followed by uprooting and separation into root and shoot parts. The root part was washed with distilled water to remove loose soil and carefully blotted with filter paper to remove surface moisture only. The plant parts were then weighed separately and placed individually in paper bags for drying in oven at 60°C. The dry weight of the plants was observed after 48h.
Antioxidant Estimation
Extraction of antioxidants from each of the four set of plants was done after 3h and 9h of S. litura damage. The single leaflet was removed, weighed and homogenized in 2ml extraction buffer containing 0.6% polyvinyl propylidine, 10mM EDTA, 10mM cystein and 100mM Tris HCl in ice cold conditions. The homogenized leaf was centrifuged in 10,000rpm for 10min in 4°C. The pellet was discarded and supernatant was immediately stored on -20°C until further use. The leaf extract thus obtained was used to study the enzymes Superoxide dismutase (SOD), Glutathione reductase (GR) and Polyphenol oxidase (PPO).
SOD was analysed as per Verma, et al. [33] using 10µl of leaf extract with 100mM sodium phosphate buffer (pH 7.4), 20mM methionine (prepared in 0.1ml of 1% (v/v) Triton X-100), 10mM Hydroxylamine hydrochloride and 100µM EDTA. After incubation at 37°C for 5min in dry bath, 100µM riboflavin was added to the mixture and reaction was initiated by incubated under 20W fluorescent light at room temperature. The reaction was stopped using Greiss reagent and the absorbance was taken at 543nm against no enzyme and no riboflavin control. SOD was calculated using the following equation:
$$\text{SOD units / ug of protein} = \frac{\left[ \text{O.D. of control} / \left( \text{O.D. of sample} - 1 \right) \right]}{10\text{ug of protein}}$$
GR activity was studied at 340nm used in the reaction mixture containing 100mM Tris-HCl buffer (pH 7.8), 0.5mM Glutathione disulphide (GSSG), 2mM EDTA and 0.2mM NADPH along with the leaf extract, as per Rao, et al. [34]. Enzyme activity was studied for 3min duration with an interval of 12 seconds using UV-Vis spectrophotometer (Shimadzu UV-1800, Shimadzu, Japan).
For PPO analysis, the insect induction was given for 48h. 0.1g of leaf was flash freezed in liquid nitrogen and homogenized with 50mM Sodium phosphate buffer (pH 6.0). The sample was vortexed for 5min and centrifuged at 10000rpm for 15min at 4°C. Supernatant was transferred to a new eppendorf tube and centrifuged at 10000rpm for 30min at 4°C. The final supernatant was used for PPO estimation. 100mM phenyl methyl Sulfonyl fluoride (PMSF) was added before analysis. 30µl of leaf extract sample was pipetted in a well of the 96 microliter well plate in a 200µl reaction mixture containing 5.75mM L-DOPA and 100mM sodium phosphate buffer (pH 8.0). The reaction was performed on ice in dark conditions. The kinetic rate reading of the reaction was taken at 490nm at 1 minute interval for 10 minutes using the ELISA plate reader (Electronics Corporation of India, India) and was calculated as per Nguyen, et al. [35].
Chlorophyll A and B Estimation
For this set of experiment the larvae were allowed to feed on leaves for seven days in the clip cage. Fifth leaf of the damaged and undamaged plants was removed and 0.1g leaf was homogenized in 10ml of 80% acetone. The extract was centrifuged at 3000rpm for 10min at 4°C. The supernatant was used for absorbance at 663 and 645nm for calculating Chl a and b according to the equations given by Arnon, et al. [36].
Larval Preference Assay
Fifth leaf of each of the four treated plant sets (after 48h of insect damage) was removed and their leaf area was measured as per O'Neal, et al. [32]. One leaf per treatment set was placed at equal distance in a 20cm diameter Petri dish. Four S. litura of third instar were released at the centre of the plate for 24h. The insects were satiated with water for 12h before the experiment. Leaf area was measured again after feeding to calculate the amount of leaf consumed from each set.
Insect Performance Assays
Fifth leaves of each set were placed individually in containers lined with moist filter paper. Four containers, each for the leaves of one set, were kept without larvae to determine leaf moisture loss. Initial weight of the leaves was observed before placing them separately in 9cm Petriplates lined with Whatmann No.1 filter paper. Each of these plates received one newly moulded final larvae, which were satiated with water overnight. Larval weight gain and the weight of the remaining leaf were observed after 24h of feeding. The leaf weight at the end of each experiment was subtracted from its initial weight to calculate the amount ingested. Change in leaf weight due to loss of moisture was determined from leaves in containers without larvae and deducted from the experimental and control leaves. Nutritional indices were calculated as per Waldbauer, et al. [37] as follows:
The Waldbauer indices were calculated as follows: Consumption Index (CI) = [(leaf mass ingested)/(larval mass gain x number of days)], Approximate Digestibility (AD) = [(leaf mass ingested- frass mass)/(leaf mass ingested)], Efficiency of Conversion of Digested food (ECD) = [(larval mass gain)/(leaf mass ingested- frass mass)] and Efficiency of Conversion of Ingested food (ECI)= [(larval mass gain)/(leaf mass ingested)].
Statistical Analysis
Data were analyzed using SPSS 23 (IBM Chicago, IL, USA). Data normality and homogeneity of variance was determined using one-sample Kolmogorov-Smirnov test and Levene’s test, respectively. Trichome number and PPO data was normalized by log transformation whereas SOD and GR data was normalised using square root transformation. Trichome number and dry weight data was analysed by using one way ANOVA with LSD as post hoc treatment. Larvae preference was analysed using non-parametric χ2 Friedman’s test.
Results
Trichome number was higher on the adaxial as well as abaxial surface of tomato leaf in PSB supplemented plants (Table 1). Glandular trichomes were significantly increased on both the surfaces, with PSB supplementation especially upon insect damage (Univariate ANOVA; Adaxial F(3,39)=72.631, P<0.001; Abaxial F(3,39)=36.855, P<0.001 ). Irrespective of insect damaged, non-glandular trichome number was higher in PSB supplemented plants in comparison to control plants (Univariate ANOVA; Adaxial F(3,39)=9.427, P<0.001; Abaxial F(3,39)=7.528, P<0.001 ). On the contrary, unsupplemented control plants showed no significant difference in trichome number upon insect damage, except for non-gladular trichomes on the adaxial surface (P=0.021). Insect damage positively influenced trichome number in both control and PSB supplemented plants (P=0.001).
| Glandular adaxial | Glandular abaxial | Non glandular adaxial | Non glandular abaxial | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Control Undamaged | 17.0 ± 2.46c | 12.5 ± 1.38d | 10.4 ± 1.55b | 28.4 ± 3.93b | |||||||
| Control damaged | 12.2 ± 0.85c | 25.4 ± 2.36c | 18.7 ± 3.331a | 28.0 ± 3.34b | |||||||
| PSB Undamaged | 51.9 ± 5.92b | 39.7 ± 1.52b | 10.9 ± 5.93b | 51.3 ± 5.67a | |||||||
| PSB damaged | 80.8 ± 3.22a | 74.0 ± 4.64a | 28.7 ± 3.61a | 46.1 ± 4.57a |
Table 1: ANOVA table of trichomes on adaxial and abaxial surface of newly emerged leaf of tomato plant after 7 days of damage (me
Mean followed by similar letter(s) in each column, are not significantly different at 0.05% level of probability according to LSD Test. Table 1: ANOVA table of trichomes on adaxial and abaxial surface of newly emerged leaf of tomato plant after 7 days of damage (mean ± SE).
Mean fresh weight of the four treatments did not change significantly. However, dry weight increased significantly in PSB supplemented plants irrespective of insect damage (Univariate ANOVA; F(1,3)=6.357, P<0.001) (Table 2). Further, shoot dry weight also increased significantly with insect damaged (P=0.029). This difference was prominently observed in damaged PSB supplemented compared to damaged control plants (P<0.001).
| Treatments | Shoot | Root | Total | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control Undamaged | 0.72 ± 0.014 c | 0.16 ± 0.004 | 3.34 ± 0.068 | |||||||
| Control Damaged | 1.20 ± 0.027 b | 0.22 ± 0.004 | 3.30 ± 0.159 | |||||||
| PSB Undamaged | 1.20 ± 0.026 b | 0.20 ± 0.005 | 3.70 ± 0.091 | |||||||
| PSB Damaged | 1.40 ± 0.028 a | 0.23 ± 0.04 | 3.44 ± 0.109 |
Table 2: Mean (+ SE, N=22) ANOVA table for shoot and root dry weight of control and PSB supplemented plants.
Mean fallowed by similar letter(s) in each column, are not significantly different at 0.05% level of probability according to LSD Test. PSB means Phosphate Solubilizing Bacteria, Damaged means insect damaged and Undamaged means not damaged. Table 2: Mean (+ SE, N=22) ANOVA table for shoot and root dry weight of control and PSB supplemented plants.
SOD levels in undamaged control plants were significantly high compared to the other three treatments at 3h (Univariate ANOVA; F(3,36)=20.729, P<0.001) (Figure 1a). On the contrary, at 9h SOD increased in damaged control and undamaged PSB plants (Univariate ANOVA; F(3,36)=38.857, P<0.001). Moreover, at 9h, damaged PSB supplemented and control undamaged plants showed equivalent levels of SOD which were significantly low than damaged control (P<0.001) and undamaged PSB plants (P<0.001).There was no significant change in the GR activity in the four treatments at 3h (Univariate ANOVA; F(1,26)=2.009, P=0.057) (Figure 1b). Moreover, GR significant increased from 3h
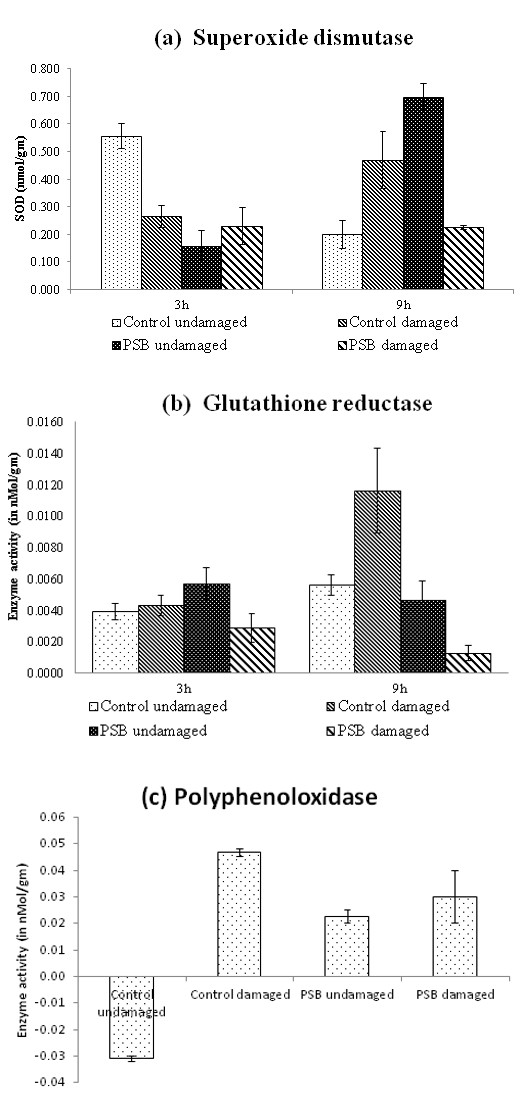
to 9h in insect damaged unsupplemented control plants only (Univariate ANOVA; F(1,66)=3.288, P<0.005). The enzyme activity in this set showed a drastic increase in at 9h (Univariate ANOVA; F(3,26)=6.323, P<0.002). Further, at 9h GR activity tends to increase in insect damaged unsupplemented plants compared with undamaged control plant (P=0.073). Also, in PSB supplemented plants as well GR significantly increased with insect damage at 9h (P=0.001). At 48h, the presence of PPO in undamaged control plants was negligible. However, PPO increased with insect damaged in both control and PSB plants (Figure 1c).
Among the two types of chlorophylls only chlorophyll a levels were significantly high in undamaged control plants compared the other treatments (Univariate ANOVA;
F(3,27)=3.905, P<0.05) (Table 3). No significant changes in Chlorophyll a & b levels were found with the other three treatments (Univariate ANOVA; F(3,27)=0.168).
| Treatments | Chl a | Chl b | ||||||
|---|---|---|---|---|---|---|---|---|
| Control Undamaged | 38.5 ± 1.46a | 35.8 ± 1.23 | ||||||
| Control Damaged | 35.3 ± 0.31b | 35.1 ± 1.57 | ||||||
| PSB Undamaged | 35.3 ± 0.20b | 34.1 ± 1.06 | ||||||
| PSB Damaged | 34.5 ± 1.00b | 34.7 ± 2.81 |
Table 3: Percentage (+SE, n=10) of differently treated tomato leaf consumed by third instar _S. litura_ during leaf preference as
In choice based larval preference assay, feeding area was least on PSB supplemented plant that was previously damaged. However, there was no significant difference between all treatments in amount of leaf area consumed (Table 4).
Treatments N Leaf consumed χ2 P Control undamaged 10 39.0 ± 8.27 3.143 0.958 Control damaged 10 39.0 ± 6.22 0.833 1 Bacteria undamaged 10 40.5 ± 5.33 1.333 0.998 Bacteria damaged 10 21.7 ± 4.88 2.667 0.914 χ2 Friedman test: P<0.05. Table 4: Percentage (+SE, n=10) of differently treated tomato leaf consumed by third instar S. litura during leaf preference assay.
Larval consumption was highest on previously damaged control plant leaf whereas the least consumption was observed on damaged PSB (Table 5). Larvae weight gain was highest on PSB undamaged plants followed by control undamaged, control damaged and damaged PSB bacteria damaged after 24h of insect feeding. The result from efficiency of conversion of digested food to body shows that larvae converted high amount of consumed food from PSB supplemented undamaged and damaged leaf (173.1 and 136.5% respectively). Larva utilized high percentage of ingested food from undamaged plant compared to previously damaged plants.
| Control | Control | PSB undamaged | PSB damaged | |||||
|---|---|---|---|---|---|---|---|---|
| undamaged | damaged | |||||||
| Larval weight gain (g) | 0.04a ± 0.002 | 0.03b ± 0.01 | 0.04a ± 0.02 | 0.01c ± 0.003 | ||||
| Consumption index (g g-1 day -1) | 0.06c ± 0.01 | 0.17a ± 0.01 | 0.08c ± 0.02 | 0.12b ± 0.03 | ||||
| Approximate digestibility (%) | 57.60c ± 6.00 | 173.40a ± 5.50 | 70.60 b ± 9.10 | 52.3c ± 8.10 | ||||
| Efficiency of conversion of digested food (%) | 69.90b ± 34.10 | 40.80b ± 5.60 | 173.10a ± 46.70 | 136.5ab ± 49.20 | ||||
| Efficiency of conversion of ingested food (%) | 81.70a ± 9.33 | 25.10c ± 0.86 | 68.20a ± 11.34 | 42.7b ± 8.99 |
Table 4: Mean ± SE (n = 10) nutritional indices of Spodoptera litura in bacteria supplemented and control plant leaves.
Discussion
When plants are stressed by biotic or abiotic factors, major physiological and morphological changes are induced and plant resistance mechanisms are generally activated. In spite of major advances in our understanding of the plant responses, little information is available on how PSB supplementation affects induced responses. Through present study we found a positive simulation in number of trichomes present in PSB supplemented plants with almost 2 times increase in granular trichomes. These trichomes further increased with insect damaged on PSB supplemented plants to > 4 times on both adaxial as well as abaxial surface. Glandular trichomes have heads containing various sticky and/or toxic exudates that may be secreted onto the plant surface or may rupture on contact with herbivores, causing irritation, entrapment, or death Simmons and Gurr, et al. [38]. Plant uses these structures as resistance mechanism against biotic as well as abiotic stresses. Furthermore, our results also indicated a significant increase in non-glandular trichomes was also observed in insect damaged PSB supplemented plants. Unlike glandular trichomes, non- glandular trichomes do not have such heads, but they affect herbivores by mechanically obstructing their movement across the plant surface [38].
Our results also showed that PSB supplementation substantially increased shoot growth and development of tomato plant in terms of increased shoot biomass. Increased plant biomass due to microbial fertilizer administration in various crops is well documented [39, 40, 41]. Our study indicated increased shoot biomass in insect damaged plants of both control as well as PSB supplemented. This may be due to increased allocation of dry mass towards shoot as a mean of compensatory mechanism to maintain functional shoot at the expense of root [40]. Water retention in shoots of PSB supplemented plants was higher than unsupplemented undamaged control plants. However, interestingly water retention in root and shoot was higher in insect damaged control plants compared to the other three sets of treatments (supplementary data).
The amount of chlorophyll a was found to be elevated compared to chlorophyll b. Chlorophyll a is recognized as the main pigments which convert light energy into chemical energy. Chlorophyll b as accessory pigments acts indirectly in photosynthesis by transferring the light it absorbs to chlorophyll a [43]. These help tomato plant to capture and convert more light energy to chemical energy. Further, unsupplemented plants without insect damaged show a significantly higher Chlorophyll a, on the contrary rest of the three treatments has similar levels. When comparing these results to induction of trichomes (Table 1), the pattern is reversed which may be attributed to nutritional allocation from plant physiological processes to defence.
During insect preference assays larvae consumed less percentage of leaf area from PSB supplemented plants with insect damage than all three treatment plants however, there was no significant difference between all treatments. Less percentage consumption from PSB supplemented plant leaf is due to induced toxic secondary metabolites from previous insect damage. Previous studies have indicated that after a prior attack by herbivores, plants can be induced to increase their polyphenol oxidase activity, which decreases the nutrient value of the foliage and reduces the feeding preference of the insects [44, 45, 46].
Our results indicate no significant change in antioxidants (SOD and GR) post 3h of insect damage. However, in case of control damaged plants antioxidants increased after 9h. This indicates that even though oxidative stress initiates within minutes of insect feeding [47], our results suggest a systemic antioxidants response in tomato is generated only after 9h. Being sensitive to metabolic fluctuations, SOD is the part of first line of defense in case of biotic and abiotic stress, where its levels had been reported to increase [48]. Contrary to unsupplemented treatment, plants with PSB supplementation did not indicate elevated SOD or GR levels at 9h, indicating susceptibility to insect induced oxidative damaged at early hours. Moreover, increased GR helps the plant in up regulating cells glutathione pool to improve stress tolerance [49]. On the contrary, PPO activity significantly increased in PSB damaged plants at 48h which indicates activation of resistance mechanism against insect induced oxidative stress at this time point. PPO catalysis oxidation of polyphenols to quinones and activates defense against insects [50]. This is also evident in damaged control plants at this time. Although after comparing the overall antioxidant profile, PSB supplementation does not indicate positive effect on insect induced oxidative stress.
Larval weight gain on undamaged plant leaf on both sets of control as well PSB supplemented plants compared to damaged ones. Specifically, PSB supplemented damaged plants showed least larval weight gain although consumption index in these plats was next to highest among the four sets. Subsequently, percentage approximate digestibility of PSB supplemented plants with insect damage was least. Possible reason for this feeding behaviour is the presence of increased level of glandular trichomes observed in these treatments which increases insect resistance. Moreover, the presence of defence related metabolites (such as α tomatin and rutin) in tomato plant, which reduces the quality of food [51]. Also, after having been damaged by insect feeding, plants increase their polyphenol oxidase activity and reduce their foliar carbohydrate concentrations. This increase in oxidase activity and decrease in carbohydrate content might decrease the nutritive value and thus reduce performance of subsequent feeding insects [45, 46, 52, 53, 54]. On the other hand control undamaged plants showed least consumption index but highest larval weight gain.
Although, PSB supplemented undamaged plants also showed high larval consumption and hence the percentage approximate digestibility, which shows improved nutritional quality in these plants resulting in increased palatability. Moreover, larvae consumed higher amount of food from damaged leaves than undamaged one. This is may be because of damage by insect releases some volatile chemicals which initiate subsequent insect to choose first.
The result from efficiency of conversion of digested food shows that higher percentage of digested food was converted in to larvae body from PSB supplemented undamaged plants. This is related with higher weight gain of larvae from PSB supplemented undamaged plants. Conversion of ingested food to larvae body was higher from undamaged plants. This is because in damaged plants defensive toxic chemicals are higher and larvae eliminate these toxic substances through excretion rather than converting in to the body.
Compared to insect damaged control plant lacking PSB supplementation, tomato plants with PSB administration in tomato shows a positive effect on its insect induced responses. At early hours of insect exposure, both these plants showed equal susceptibility to oxidative stress. Although contrary to unsupplemented plants, PSB plants after 48h of insect damage, showed an increased mechanical resistance towards insect in terms of elevated trichome number. However, biochemical responses in these plants at 48h showed no significant change, which is indicated by both chlorophyll and PPO profiles. On the other hand, when growth period of tomato in PSB is prolonged, a positive effect on shoot biomass is observed after 48h of insect damage. Insect feeding behaviour towards these plants indicated a decline in larval weight gain on previously damaged plants. However, an increased consumption index does indicate increased palatability of the plant, possibly due to high phosphorus availability. Corresponding to the increased consumption index reduced efficiency of ingested food conversion was observed.
Therefore, results indicate that PSB administration improves growth as well as induced resistance in the tomato, which further aids in increasing growth period with PSB. However, in order to understand intricate mechanisms underlying the microbial fertilizer-plant- insect interactions field based studies in this direction are required. Also, an effective implementation of these microbial fertilizers in sustainable practices can be formulated based on their interactions with the rhizospheric microbiota.
Acknowledgement
The authors would like to acknowledge NAM S&T centre for granting RTF-DCS fellowship 2015 to Mr. Melesse Angulo Torro for this work. The authors would also like to thank the post graduate students involved with the experimentation and observation of various experiments.
References
-
Fifth leaf of each of the four treated plant sets (after 48h of insect damage) was removed and their leaf area was measured as per O'Neal, et al. [32]. One leaf per treatment set was placed at equal distance in a 20cm diameter Petri dish. Four S. litura of third instar were released at the centre of the plate for 24h. The insects were satiated with water for 12h before the experiment. Leaf area was measured again after feeding to calculate the amount of leaf consumed from each set.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind