The Pathophysiology of Trypanosomiosis Associated Anaemia: A Multifactoral Phenomenon
The pathobiology of anaemia in trypanosomiosis remains one of the aspects that still lacks thorough understanding. Extra vascular haemolysis is often identified as a major contributor to anaemia, primarily due to the mononuclear phagocytic activity of the host. This sets off a series of reactions, ultimately resulting in a significant alterations in haemato-biochemical indices. The magnitude of these alterations becomes more pronounced during successive waves of fluctuating parasitemia. However, it's important to note that the basic cause of anaemia is intricate and typically multifaceted, mainly affecting the cellular integrity of red blood cells, terminating into hemolytic anemia and/or increased erythrophagocytosis. Injuries inflicted by the flagella and microtubules of the parasite onto the membranes of erythrocytes further facilitate erythrophagocytosis of damaged RBCs. Additionally, erythrocytes, reticulocytes, and platelets often bind to trypanosomes via. sialic acid receptors, causing injuries to the cells promoting their rupture. A number of factors attributing to haemolytic anaemia include trypanosome are discussed in the present review. This review elaborates on the details and interactions of all these contributing factors.
Introduction
Many aspects of trypanosomiosis pathogenicity require further elucidation. While immunosuppression and hypoglycemia are well-documented in the literature, anaemia remains a less understood aspect of trypanosomiosis pathogenesis. Despite being a pivotal immunopathological feature of animal trypanosomiosis, anaemia remains a topic of ongoing debate. The complexity of anaemia in trypanosomiosis is exacerbated by its multifactorial origin [1], adding to its intricacy. This review aims to consolidate information concerning anaemia in trypanosomiosis, including its stages, a glimpse into its multifactorial etiology, and a multidimensional exploration to enhance our comprehension of the disease.
The Complexity of Trypanosomiosis Inflicted Anaemia
The pathophysiology of anaemia associated with trypanosomiosis is exceedingly intricate and surpasses with those caused by other infectious agents due to its multifaceted origins [1]. In several other parasitic diseases, anaemia directly results from blood loss caused by the parasite, as seen in hookworms [2], or the blood-sucking habits of the parasite, as in haemonchosis [2]. Alternatively, it may arise from haematophagia and subsequent hemorrhages during the migratory phase, or injurious nature of parasite involved, as observed in immature liver flukes [3]. Additionally, anaemia can be induced by the discharge of antihaematinic metabolites into the host circulation by the parasite, as seen in fasciolosis [4]. However, in trypanosomiosis, the etiology of anaemia is notably intricate. It is assumed that in trypanosomiosis cascade of events culminate into cardiovascular collapse owing to haemolytical anaemia [5].
To facilitate comprehension, the anaemia in trypanosomiosis has been categorized into three distinct phases (Figure 1).
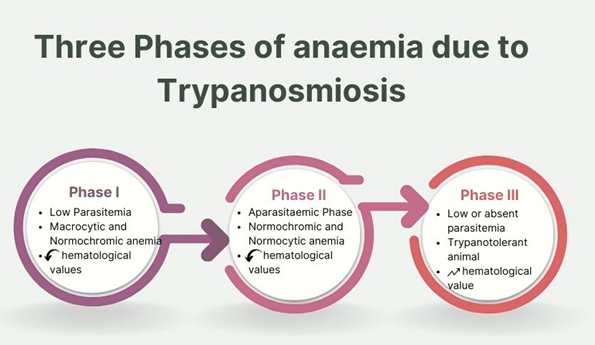
Phase I: The Acute Crisis Initiation Phase: This phase marks the initial onset of anaemia, and coincide with initial symptoms and establishment of trypanosomes inside the host’s blood stream. Following the establishment of the infection, parasitemia periodically surges with characteristic fluctuations in its levels [6, 7]. During this phase, the anaemia is macrocytic, normochromic [8]. Additionally, a decline in hematological and erythrocytic values becomes evident. Sub- acute phases are seen in experimental infection of rodents with Trypanosoma congolense [9] and T. brucei [7].
Phase II: The Progression or the Chronic Phase: Following the acute crisis phase, this stage depicts lower levels of fluctuating parasitemia, spanning up to months. In the case of tissue-dwelling trypanosomes like the T. brucei group, this phase is also referred to as the aparasitaemic phase, wherein the parasites tends to establish themselves outside the bloodstream and are either found in reduced numbers [10] entirely absent in blood smear [11]. Throughout this phase, the anaemia transitions to a normochromic and normocytic type [12]. Hematological and erythrocytic values are generally depressed during this period.
Phase III: The Stabilizing or the Recovery Phase: This phase is distinguished by either the low, infrequent, or complete absence of parasitemia. During this stage, the previously depressed erythrocytic values start to normalize, accompanied by the resolution of other pathological alterations [5]. This phase leads to self-recovery and is commonly referred to as the trypanotolerant stage [11]. Wild animals often demonstrate a moderate level of trypanotolerance, reflecting their relative ability to control the excessive proliferation of trypanosomes within their bodies and limit their pathogenic effects [11].
Multifactorial Etiology of Trypanosomiosis Associated Anaemia
The primary subtype of anaemia prevalent in trypanosomiosis is hemolytic anaemia. It is end result of complex interactions between various host and parasite- associated factors, which can act independently or synergistically, ultimately leading to a fatal form of hemolytic anaemia. Common attributes (Figure 2).
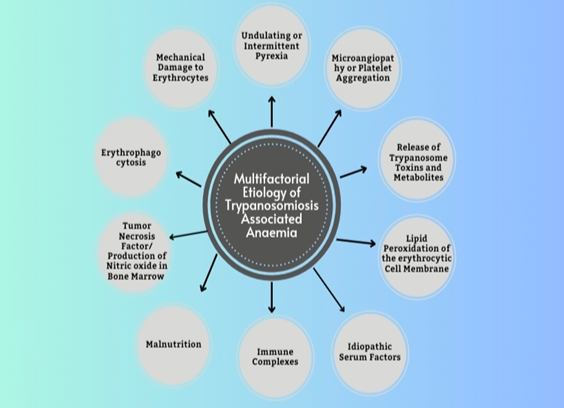
Anaemia Arising Due to Mechanical Damage to Erythrocytes: In acute infection of trypanosomiosis, the forceful movement of flagella and microtubule-reinforced bodies in millions of parasites causes significant mechanical damage to erythrocytic cell membrane [13]. This erythrocytic cell membrane damage is further exacerbated by adhesion of various blood cells like erythrocyte, platelets, and reticulocytes to the trypanosome surfaces through sialic acid mediated receptors, resulting in additional cell membrane damage [14]. As a result, multiple points of disruption emerge on the surface of erythrocyte membranes at the points of attachment with the body of trypanosomes. Furthermore, there have been documented instances of mechanical harm to the vascular endothelium by the invasion of trypanosomes penetrating the tissues [15]. Together, these elements contribute to the mechanical damage of erythrocytes, ultimately resulting in extensive rupture of erythrocytes and subsequent anemia.
Anaemia Caused due to Undulating or Intermittent Pyrexia: The Variant Surface Glycoproteins (VSGs) of trypanosomes are recognized to exert a notable influence on the degrees of recurring fevers and changing levels of parasitic presence [11]. Remarkably, there have been substantial elevations in measures of osmotic fragility and the permeability of erythrocytes, particularly at temperatures exceeding the host’s typical body temperature [16]. Furthermore, elevated body temperature reduces the flexibility and lifespan of erythrocytes in vivo [17]. As a result, heightened temperatures have been linked to an augmented occurrence of immunochemical reactions, culminating in the lipid peroxidation of erythrocytes [18]. Hence, in phases of recurring fevers when temperatures reach their peak, the stability of erythrocytes is diminished, making them more prone to rupture [16].
Anaemia Due to Microangiopathy or Aggregation of Platelets: Microangiopathy or platelet aggregation is commonly linked with both intact as well as fragmented trypanosomes [19]. The harm inflicted on trypanosomes is more frequent during trypanocidal drug treatment. This harm causes the burst of auto-platelet antibodies, subsequently initiating procoagulant release leading to fibrin deposition. This ultimately terminates into formation of microthrombi further attributing to dispersed intravascular coagulopathies [18]. Weaking of erythrocytes, owing to reasons discussed above, tends to their breakdown into fragments as they move through the fibrin deposits of microthrombi [15], eventually leading to dispersed sites of intravascular coagulation [11].
Anaemia Caused by Release of Toxins and/or Metabolites of Trypanosomes: It has been well understood for a significant duration that both viable and deceased trypanosomes secrete a range of bioactive chemical compounds like neuraminidase, pyruvates, free fatty acids, proteases, phospholipase, etc. capable of harming erythrocytes [1]. Neuraminidases are produced in laboratory settings by various trypanosome species. They are recognized for elevating the susceptibility of erythrocytes [20] to get phagocytosed by the mononuclear phagocytic system (MPS). Neuraminidases enhance the vulnerability of erythrocytes by splitting the sialic acid component found on the cell surface [21]. They are also known to hinder erythropoiesis by causing harm to the erythropoietin hormone [22]. Trypanosomes have the ability to release proteases from their reservoirs and also from the bodies of compromised or deceased parasite [18]. This feature is very common post therapy when a large number of trypanosomes are available inside the host’s body. Proteases cause harm to erythrocytes and vascular endothelium by cleaving sialic acid segments from the cell membrane in the glycopeptide form [23]. Reports indicate that toxic by-products, generated through the metabolism of aromatic amino acids by trypanosomes, directly affect the erythrocyte cell membrane, leading to imbalances associated with osmotic fragility and eventual lysis of erythrocytes [18]. Phenylalanine undergoes catabolism to phenylpyruvate that hampers mitochondrial gluconeogenesis [18]. Moreover, in the context of trypanosomiosis, tryptophan is metabolized into indole-ethanol that harms erythrocyte cell membranes [18], rendering them more vulnerable to rupture.
Anaemia due to Lipo-Peroxidation of Erythrocytic Cell Membrane: Irrespective of the factors involved, the process of anemia in trypanosomiasis is closely connected to the production of free radicals and superoxides stemming from lipid peroxidation. These oxidative substances are known to impact the erythrocytes [18, 24] by targeting polyunsaturated fatty acids and proteins found in the membrane of erythrocytes [25, 26]. On occasion, red blood cells experience direct influence, resulting in oxidative hemolysis [22, 24]. During trypanosomiasis, the formation of antigen-antibody complexes containing sialic acids plays a role in erythrophagocytosis [27] and the ensuing anemia [26]. Animals infected with trypanosomiasis have been noted to exhibit a significant deficit in their systemic antioxidant capacity [28]. The vulnerability of erythrocytes to harm from free radicals after exposure to hydrogen peroxide is extensively documented in several trypanosome species [22]. The peroxidation of erythrocytes leads to the generation of substantial quantities of lipid peroxidation byproducts, which diminish antioxidant capacity and worsen the erythrocytes’ diminished ability to resist oxidative stress [18]. Trypanosomes generate neuraminidase, which removes sialic acid from the surface of erythrocytes, rendering red blood cells more susceptible to phagocytosis, elevating serum sialic acid levels and frequently coinciding with the development of anemia [29]. Erythropoietin levels are known to undergo regression owing to sialic acid fragmentation, thereby aggravating anaemia [22]. As parasitemia increases in trypanosomiasis, glycolysis leads to the buildup of pyruvate in the host’s body [30], This buildup leads to acidosis and a decreased affinity of hemoglobin for oxygen [31], thereby increasing the strain on erythrocyte membranes. Acidosis is once more linked to the suppression of the tricarboxylic acid cycle (TCA) within the mitochondria [32], further exacerbating anemia. After the autolysis of dead trypanosomes, a substantial quantity of phospholipase A1 and lysophospholipase A1 [33] is discharged into the bloodstream, leading to erythrocyte deterioration, impairment of vascular endothelial cells, and hemolysis [34]. These phospholipases are predominantly characterized by their palmitic, stearic, and linoleic acid composition [35], and they subsequently liberate free fatty acids (FFA) from phosphatidylcholine. inoleic acid exhibits detergent- like properties and is responsible for causing pronounced hemolysis and cytotoxic effects [33]. The peroxidation of erythrocyte membranes by lipids is frequently connected with reduced membrane flexibility and a decrease in the activities of membrane-bound enzymes [18], thereby worsening anemia. Lipid peroxidation and oxidative harm to erythrocyte membranes are indicators of oxidative stress, which is associated with T. evansi infection in murine models [36].
Anaemia Caused due to Idiopathic Serum Factors: In trypanosomiosis, there have been observations of two unknown (idiopathic) serum factors, which are essentially heat-stable proteins not originating from trypanosomes. These proteins inhibits erythropoiesis activity [37], indirectly affecting erythroid colony functions [22], thereby aggravating anemia.
Anaemia Caused by Immune Complexes: Within trypanosomiosis, immunological processes result in the surface removal of autologous IgM and IgG antibodies and factors related with C3 compliment from erythrocytes [1]. The emergence of autoantibodies usually occurs after the initial surge in parasitemia, concurrently with lower Packed Cell Volume (PCV). Consequently, erythrocyte surfaces become capable of binding auto- or polyreactive antibodies, and they may also become sensitized through the absorption of immune complexes [1]. Additionally, erythrocytes absorb trypanosome molecules passively, anti-trypanosome antibodies attachment occurs leading to their removal from host [1]. The remarkable increase in tissue macrophages during trypanosomiosis has been well-documented [18] and is facilitated through the interplay of various lymphokines, antigen-antibody complexes, and C3b complement fragments [38], thereby playing a role in cytokine-mediated erythrocyte depletion in the context of trypanosomiasis [1]. Cytokines such as TNF-α, IFN-γ, etc. [39] are often implicated as contributing factors to anemia. Additionally, hypoferremia is sometimes associated with IL-6 and hepcidin response [40].
Anaemia Arising Due to Malnutrition: Trypanosomiosis triggers inappetence and anorexia, leading to lower energy and depleted body score and tissue reserves. This depletion is attributed to the glycogenesis, deficiencies in essential amino acids, vitamin C, and B vitamins [18]. Poor ATP supply to erythrocytes causes alterations to the erythrocytic membrane surface, weakening it, increasing its osmotic fragility, and promoting hemolysis [41], consequently accelerating erythrocytic destruction.
Anaemia Caused by Tumor Necrosis Factor/ Production of Nitric Oxide in Bone Marrow: The production of Tumor Necrosis Factor (TNF-α) by monocytes is a significant contributor of anemia in trypanosomiosis [42]. In this condition, there’s an observed increase in proliferation of bone marrow tissue, leading to elevated levels of nitric oxide (NO) within the bone marrow. This coincides with the inhibition of T-cell proliferation when stimulated with the mitogen concanavalin, both in experimental conditions and within the living organism. NO also inhibits the proliferation of hematopoietic precursors, exacerbating anemia in subsequent phase trypanosomiosis [43]. T. evansi causes the generation of NO in camels as well [44]. When NO is created, it reacts with superoxide to make peroxynitrate anion or hydroxyl radicals. Both of these produced compounds have a necrotic impact [45]. NO enters the RBC, where it causes the synthesis of methemoglobin and changes the permeability of the membrane, increasing cellular fragility [46].
Anaemia Caused by Erythrophagocytosis: Erythrophagocytosis by monocytes and macrophages, significantly contributes to anemia in trypanosomiosis through an immunological mechanism. This process is initiated by antibody-antigen (Ab-Ag) complexes generation on RBC membrane proceeded by trypanosome destruction. These complexes transform the erythrocytes into foreign proteins, making them susceptible to engulfment by macrophages [47]. During trypanosomiosis, there’s a modification in the antigenicity of the host animal’s erythrocytes, triggering immunoglobulin-mediated erythrophagocytosis facilitated by macrophage fragment crystallizable receptors for IgG [48]. The host animal produces IgG and IgM, which coat the modified erythrocytes, often involving IgM and IgG complexes and complement C-3 [49], thereby promoting their erythrophagocytosis by macrophages.
Besides the details of anaemia in trypanosomiosis are very much explained in detail in classical work of Mbaya, et al. [50].
Conclusion
In conclusion, trypanosomiasis-associated anemia is undoubtedly a multifactorial condition, with multiple factors working synergistically to trigger a series of reactions leading to significant anemia in affected hosts. The pathobiology of anemia in trypanosomiasis is complex, but deeper understanding of the condition aids in assessing the state of infected hosts. While extensive research has postulated many theories, addressing the challenges associated with anemia in animals remains a complex issue. Effective management of anemia in trypanosomiasis-affected hosts is a detrimental factor for their survival and reducing the overall impact of the disease.
Conflict of Interest
None of the authors is having any conflict of interest.
References
-
Jaiswal AK, Sudan V, Neha, Verma AK (2015) Insight into trypanosomiasis in animals: various approaches for its diagnosis, treatment and control: a review. Asian Journal of Animal Sciences 9: 172-186.
-
Soulsby EJL (1982) Helminthes, Arthropods and Protozoa parasites of domesticated Animals. In: 7th (Edn.), Bailere Tindal, London, UK.
-
Sharma RL, Godara R, Thilager MB (2011) Epizootiology, pathogenesis and immunoprophylactic trends to control tropical bubaline fasciolosis: an overview. Journal of Parasitic Diseases 35(1): 1-9.
-
Ganga G, Varshney JP, Sharma RL (2004) Effect of Fasciola gigantica excretory secretory antigen on rat hematological indices. Journal of Veterinary Sciences 7(2): 123-125.
-
Anosa VO (1988) Haematological and biochemical changes in human and animal trypanosomosis, Parts I and II. Revue d’ Elevage et de Med Ve’te’rinaire des pays Tropicaux 41(2): 65-78.
-
Mbaya AW, Nwosu CO, Kumshe HA (2011) Genital lesions in male red fronted gazelles (Gazella rufifrons) experimentally infected with Trypanosoma brucei and the effect of melarsamine hydrochloride (Cymelarsan®) and diminazene aceturate (Berenil®) in its treatment. Theriogenology 76(4): 721-728.
-
Mbaya AW, Ibrahim UI (2011) In-vivo and in-vitro activities of medicinal plants on haemic and humoral trypanosomes: A review. International Journal of Pharmacology 7(1): 1-11.
-
Anosa VO, Isoun TT (1980) Haematological studies on Trypanosoma vivax infection of goats and splenectomized sheep. Journal of Comparative Pathology 90(1): 153-168.
-
Isoun TT, Esuruoso GO (1972) Pathology of natural infection of Trypanosoma vivax in cattle. Nigerian Veterinary Journal 1(2): 42-45.
-
Rabo JS (1995) Toxicity studies and trypanosuppressive effects of stem bark extract of Butyrospermum paradoxum in laboratory animals. PhD Thesis, University of Maiduguri, Maiduguri, Nigeria.
-
Sudan V, Verma AK, Jaiswal AK (2017) Trypanosomosis of wild animals with emphasis on Indian scenario. Veterinary Parasitology Regional Studies and Reports 10: 25-28.
-
Maxie MG, Losos GJ (1979) Release of Trypanosoma vivax from the microcirculation of cattle by Berenil®. Veterinary Parasitology 3: 277-281.
-
Vickerman K, Tetley L (1978) Biology and ultra- structure of trypanosomes in relation to pathogenesis. In: Pathogenecity of trypanosomes, Proceedings of a workshop Nairobi, Kenya, November 20-23.
-
Shehu SA, Ibrahim NDG, Esievo KAN, Mohammed G (2006) Role of erythrocyte surface sialic acid inducing anaemia in Savannah Brown bucks experimentally infected with Trypanosoma evansi. Veterenarski Arhiv 76 (6): 521-530.
-
Anosa VO, Kaneko JJ (1983) Pathogenesis of Trypanosoma brucei infection in deer mice (Peromyscus maniculatus): Light and electron microscopic studies on erythrocyte pathologic changes and phagocytosis. American Journal of Veterinary Research 44(4): 645-651.
-
Karle H (1974) The pathogenesis of the anaemia of chronic disorders and the role of fever in erythrogenesis. Scandinavian Journal of Haematology 13(1):81-86.
-
Woodruff AW, Topley E, Knight R, Downie CGB (1972) The anaemias of Kalaazar. British Journal of Haematology 22: 319-329.
-
Igbokwe IO (1994) Mechanisms of cellular injury in African trypanosomiosis. Veterinary Bulletin 64(7): 611-620.
-
Davies CE, Robins RS, Weller RD, Broude AI (1974) Thrombocytopenia in experimental trypanosomiosis. Journal of Clinical Investigation 53(1): 1359-1367.
-
Esievo KAN (1983) Trypanosoma vivax, stock V 953: inhibitory effect of type A influenza virus anti HAV8 serum on in-vitro neuraminidase (sialidase) activity. Journal of Parasitology 69(3): 491-495.
-
Adamu S, Maashin Useh N, Ibrahim DN, Nok AJ, Esievo KAN (2009) Erythrocyte surface sialic acid depletion as predisposing factor to erythrocyte destruction in sheep experimental model of African trypanosomosis: A preliminary Report. Slovenian Veterinary Research 46(1): 19-28.
-
Igbokwe IO (1989) Dyserythropoesis in animal trypanosomiosis. Revue d’Elavage et de Med veterinaire des pays Tropicaux 42(3): 423-429.
-
Cook GMW, Heard DH, Seaman GVF (1966) A sialomuscopeptide liberated by trypsin from the human erythrocyte. Nature 188(1): 1011-1012.
-
Umar IA, Ogenyi E, Okodaso D, Kimeng E, Stanecheva GI, et al. (2007) Amelioration of anaemia and organ damage by combined intraperitoneal administration of vitamin A and C to Trypanosoma brucei brucei infected rats. African Journal of Biotechnology 6(18): 2083-2086.
-
Slater TF (1984) Free radical mechanism in tissue injury. Biochemistry Journal 222(1): 1-15.
-
Audu PA, Esievo KAN, Mohammed G, Ajanusi OJ (1999) Studies of infectivity and pathogenicity of an isolate of Trypanosoma evansi in Yankasa sheep. Veterinary Parasitology 86(3):185-190.
-
Holmes PH, Jennings FW (1976) Pathogenicity of parasitic infections. Academic Press New York, ISBN 0372-5480, New York, USA.
-
Esievo KAN, Saror DI, Ilemobade AA, Hallaway MH (1982) Variation in erythrocyte surface and free serum sialic acid concentrations during experimental Trypanosoma vivax infection in cattle. Research in Veterinary Science 32(1):1-5.
-
Nok AJ, Balogun EO (2003) A blood stream Trypanosoma congolense sialidase could be involved in anaemia during experimental trypanosomiosis. Journal of Biochemistry 133(6): 725-730.
-
Coleman RM, Brand T, Von (1957) Blood pyruvate levels of rats during haematophagus infections. Journal of Parasitology 43(1):263-270.
-
Newton BA (1978) The metabolism of African trypanosomiosis in relation to pathogenic mechanisms. In: Pathogenicity of trypanosomes. Losos G, Chouinard A, (Eds.), Proceedings of a workshop, Nairobi Kenya, pp: 17-22.
-
Seed JR, Hall JE (1985) Pathophysiology of African trypanosomiosis. In: Immunology and pathogenesis of trypanosomiosis. Tizard IJ (Ed.), CRC Press, Boca Raton, Florida, USA.
-
Tizard IR, Sheppard J, Nielsen K (1978) The characterization of a second class of haemolysin from Trypanosoma brucei. Transactions Royal Society of Tropical Medicine and Hygiene 72(2): 198-200.
-
Colley CM, Zwaal RFA, Roeofsen B, Decssen LLM (1973) Lytic and non-lytic degradation of phospholipids in mammalian erythrocytes by pure phospholipase. Biochimica et Biophsica Acta 307(1): 74-82.
-
Tizard IR, Holmes WL (1977) The release of suitable vasoactive material from Trypanosoma congolense intraperitoneal diffusion chambers. Transactions Royal Society of Tropical Medicine and Hygiene 71(1): 52-55.
-
Pandey V, Nigam R, Jaiswal AK, Sudan V, Singh RK, et al. (2015) Haemato-biochemical and oxidative status of buffaloes naturally infected with Trypanosoma evansi. Veterinary Parasitology 212(3-4): 118-122.
-
Kaaya GP, Tizard IR, Maxie MG, Vall VO (1980) Inhibition of leucopoiesis by sera from Trypanosoma congolense infected calves, partial characterization of the inhibitory factor. Tropical Medical Parasitology 31(2): 232-238.
-
Allison AC (1978) Mechanisms by which activated macrophages inhibit lymphocyte responses. Immunological Reviews 40(1): 23-27.
-
Jelkmann W (1998) Proinflamatory cytokines lowering erythropoietin production. Journal of Interferon and Cytokine Research 18(8): 555-559.
-
Nemeth E, Rivera S, Gabayan V, Keller C, Taudorf S, et al. (2004) IL-6 mediates hypoferremia of inflammation by inducing the synthesis of the iron regulatory hormone hepcidin. Journal of Clinical Investigation 113(9): 1271- 1276.
-
Jennings FW (1976) The anaemia of parasitic infections, Proceedings of the 7th International Conference of the World Association for the Advancement of Veterinary Parasitology. Thessalonica, Greece, pp: 41-67.
-
Sileghem M, Flynn JN, Logan-Henfrey L, Ellis J (1994) Tumour necrosis factor production by monocytes from cattle infected with Trypanosoma (Dutonella) vivax and T. (nannomonas) congolense, Possible association with severity of anaemia associated with the disease. Parasite Immunoassay 16(1): 51-54.
-
MacLean L, Odiit M, Sternberg JM (2001) Nitric oxide and cytokine synthesis in human African trypanosomiosis. Journal of Infectious Diseases 184(8): 1086-1090.
-
Saleh MA, Al-Salhy BM, Sanousi SA (2009) Oxidative stress in blood of camels naturally infected with Trypanosoma evansi. Veterinary Parasitology 162(3-4): 192-199.
-
Herold S, Rock G (2003) Reactions of deoxy-, oxy-, and methemoglobin with nitrogen monoxide. Mechanistic studies of the S-nitrosothiol formation under different mixing conditions. Journal of Biological Chemistry 278(9): 6623–6634.
-
Halliwell B, Chirico S (1993) Lipid peroxidation: its mechanism, measurement and significance. American Journal of Clinical Nutrition 57(supp 5): 715-724.
-
Kimeto BA (1989) Erythrophagocytosis in cattle experimentally infected with T. vivax. Bulletin of Animal Health Production Africa 143–146.
-
Weiss DJ, Klausner JS (1988) Neutrophil-induced erythrocyte injury: a potential cause of erythrocyte destruction in the anaemia associated with inflammatory disease. Journal of Veterinary Pathology 25(6): 450-455.
-
Facer CA, Crosskey JM, Clarkson MJ, Jenkins GC (1982) Immune haemolytic anaemia in bovine trypanosomiosis. Journal of Comparative Pathology 92(3): 393-401.
-
Mbaya A, Kumshe H, Nwosu C (2012) The mechanisms of anaemia in trypanosomosis: a review. In: Silverberg D (Ed.). Anaemia. In Tech, Shanghai, China, pp: 269-82.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind