An Army of Ants in the Defense of the Sacred: the Myrmecofauna (Hymenoptera, Formicidae) of the Sacred Trees of Ipeiros (North West Greece)
In mountainous Greece, cultural landscapes exhibit distinctive features, such as scattered trees marking village boundaries and sacred forests serving as locally adapted conservation systems. These sacred landscapes play a crucial role in protecting villages from natural hazards and providing essential resources for the community. The unique status of certain trees as "sacred" is maintained through supernatural fears and taboos associated with logging. While the significance of sacred forests for biodiversity is acknowledged, there is a lack of evaluation regarding the importance of individual aged trees for ant species. This study aims to fill this gap by documenting the dominant ant species associated with aged trees in sacred landscapes. Ant specimens were collected from sacred forests and individual old growth trees near churches in the Ipeiros region, providing insights into the association between tree species and ant communities. The research contributes to understanding the ecological dynamics of these culturally significant landscapes. Identification to species level provided species that are collected for the first time from the administrative area of Ipeiros, namely Camponotus gestroi Emery, 1878, Camponotus jaliensis Dalla Torre, 1893, and Camponotus samius Forel, 1889.
Introduction
In mountainous Greece, characteristic features of cultural landscapes are monumental, scattered trees or protected groves in outlying churchyards or sacred forests in close proximity to villages. The first, often mark village boundaries, tame liminal places, indicate abandoned settlements, serve aesthetics, used for communities’ important celebrations and often encircle magically present settlements acting as supernatural guardians of village entrances [1, 2, 3, 4]. On the contrary, sacred forests function as locally adapted conservation systems that serve community needs, acting as protective wood belts above villages, protecting them from natural hazards [5] and safeguarding the use of important natural resources for all community members, providing foliage fodder, dead wood or even timber in times of exceptional needs [6].
Supernatural fears and taboos associated with tree logging have as a result impressive ‘veterans’ to survive as “sacred” trees. What differentiates these trees from adjacent secular private trees are their huge dimensions, old age and maiden shape that indicate their protective status. Concerning species, there is a general typology that relates different species of oaks and maples and more rarely other species, e.g. mahaleb cherries, with outlying churches, while sacred forests species composition depends on altitude, geology, slope and management; can vary from prickly oaks to mix broadleaves or homogenous beech or black pine forests [7]. Moreover, their structure and composition can vary from dense forests to wood pastures or infilled savanna, depending on management regimes and their conservation histories [8].
While sacred forests and trees have been widely recognized for their significant contributions to overall biodiversity [9, 10] a notable research gap exists regarding the specific evaluation of the importance of individual aged and old growth sacred trees for various ant species. Ants, belonging to the Hymenoptera family Formicidae, emerge as prominent inhabitants of aged trees, exploiting the natural hollows formed in the trunk due to the accumulation of dead and rotten wood over time. Despite the ecological importance of these ant-tree interactions, there has been a dearth of studies comprehensively examining the myrmecofauna associated with individual sacred trees. This study aims to address this knowledge gap by systematically documenting and analyzing the dominant ant species inhabiting each tree, providing valuable insights into the intricate and symbiotic relationship between tree species and ant communities within sacred landscapes. Through this exploration, a nuanced understanding of the ecological dynamics within these culturally significant ecosystems is anticipated.
Methods
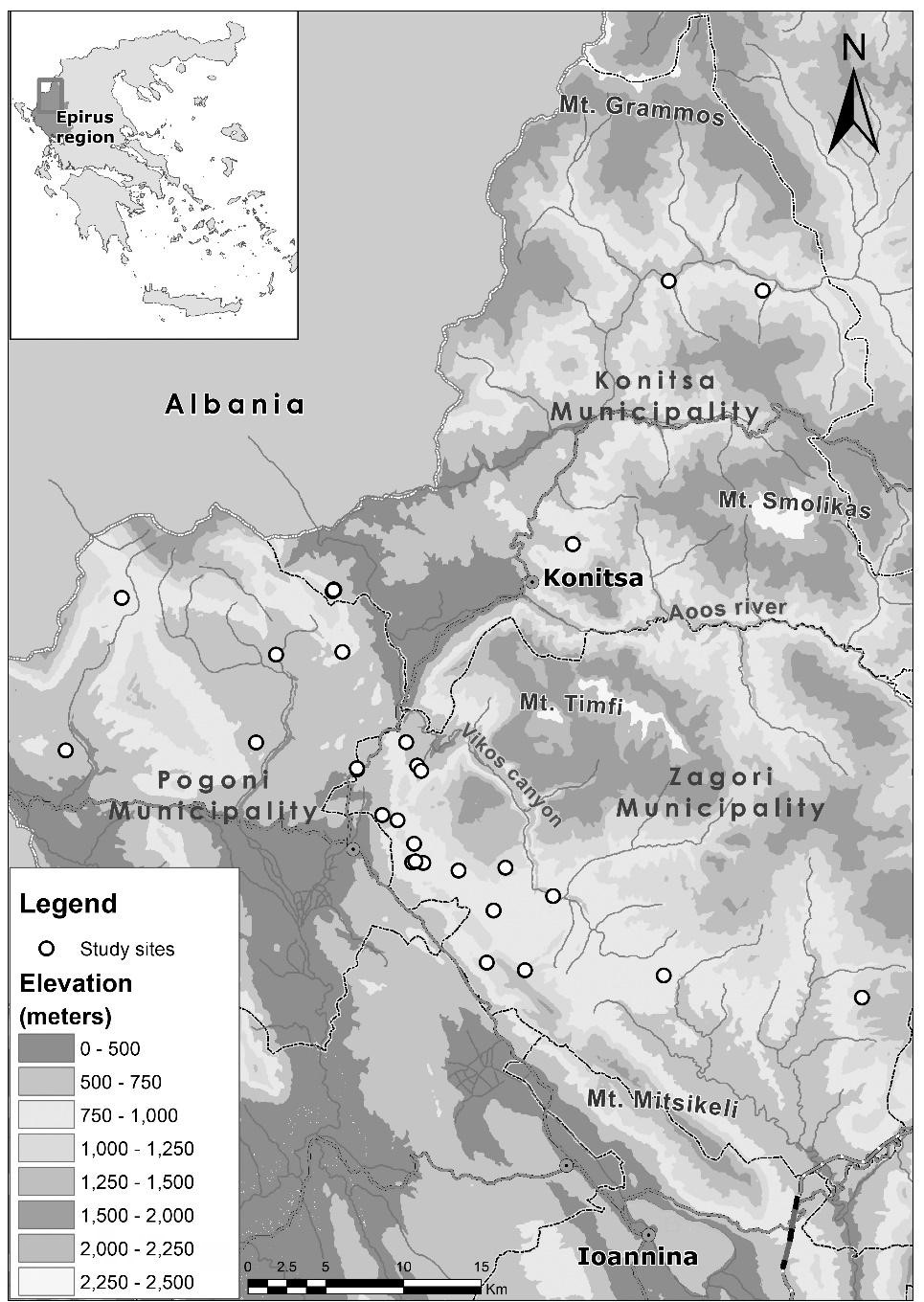
We collected ants both from sacred forests and individual old growth, aged trees associated with churches and icon stands, mainly outside settlements. Our sampling area was within Ipeiros region, near the Albanian borders and research sites belong to three different municipalities (Konitsa, Zagori and Pogoni) covering approximately an area of 2500 km². All research sites are located between 500 to 1100 m altitudes within the lower evergreen or mixed broadleaves forest zone in the west part of the area dominated by several oak species, except one site that is a beech forest in the southeast (1220 m altitude) (Figure 1). All trees sampled are deciduous broadleaves except the stinking or Grecian juniper (Juniperus foetidissima Willd. 1806) and the evergreen prickly oak (Quercus coccifera L. 1753). Research sites were selected according to two criteria: the occurrence of aged trees of monumental size known from previous research (Stara, et al. [4], Stara and Tsiakiris, Thalis-SAGE unpublished data) and the existence of features that are known to be used by ants (e.g. cavities, lows, broken branches etc.). The trees selected for sampling were the biggest in diameter in every site. Research protocol for each site recorded: the name of the site and of the nearest village, municipality, type of Sacred Natural Site, e.g, sacred forest, grove, solitary tree in a churchyard, tree species, perimeter and characteristics. Just before sampling we were knocking on the tree in order to stimulate and stress the ants so as for them to come out. Tree species and the location of each tree were recorded using a GPS unit. Research period was from 30 Apr. 2015 to 24 Aug. 2015.
From each tree, ant specimens were collected through direct sampling (hand collecting) as they were climbing up and down the tree trunk. They were stored in plastic vials containing 99% alc. The specimens are deposited in the myrmecological collection of the Zoological Museum of the University of Athens (ZMUA) and the Forest Research Institute (Hellenic Agricultural Organization Demeter). Details for the localities are given in Table 1 and Figure 1.
For the localities, the Hellenic Standards Organization system 743 (tantamount to ISO 843) was used for the transliteration of the names in Greek. The reason for this is that it standardizes the transcription of localities (in the past you would have names as Zagori, Zhagori, Zagóri etc. and some more minor alter spellings) and future researchers can follow easier the records. In addition, automated data mining engines for biodiversity (e.g. Plazi, TaxPub, EU BON and others), can easily assimilate the information provided and link data from similar publications.
All specimens were digitized using a Sony NEX 5N camera body and the Canon MP-E 65mm Macro f/2.8 lens. All pictures were curated using the Wacom Intuos Pro tablet integrated with the Adobe Photoshop CC 15.5 software. The stacking software used was Zerene Stacker (Zerene Systems LLC, http://zerenesystems.com/cms/).
Results
List of Ant Species Identified from the Sacred Trees
For each species recorded we provide its distribution for Greece according to literature and personal (CG and AL) records. The parenthesis provides a more detailed catalogue of the localities within Greece (in alphabetical order) that each species has been collected according to 1st level Administrative Regions and islands. We also give a list of other countries where the species has also been recorded from. Voucher codes are provided for the specimens that were deposited to the ZMUA. When voucher codes are missing, the specimens were used for DNA analyses after their identification.
New Records Dolichoderinae Liometopum microcephalum (Emery, 1878) (Figure 2) = Formica microcephala [11] Materials Examined: GREECE
- 1 ☿; Zagori, Vitsa, Agios Nikolaos – Livadakia; 39.8535°N, 20.7326°E; 30 Apr. 2015; KS, ΑΚ, ThV leg.; ZMUA MYRM 2017–000111
- 1 ☿; Zagori, Mesovouni, Agios Charalampos; 39.9323°N, 20.6262°E; 01 May 2015; KS, RT, ΑΤ, DT leg.
- 13 ☿; Pogoni, Vissiani, Abel monastery; 39.9457°N, 20.5494°E; 31 Jul. 2015; RT, KS leg.
- 11 ☿; Zagori, Fragades, Agios Christoforos church; 39.8194°N, 20.8619°E; 02 Aug. 2015; RT, KS leg.
- 15 ☿; Konitsa, Peklari, Agios Athanasios church; 40.0673°N, 20.7821°E; 07 Aug. 2015; RT, KS leg.
- 1 ☿; Zagori, Monodentri, Metamorfosi Sotira church; 39.8786°N, 20.7403°E; 09 Aug. 2015; KS leg.
- 15 ☿; Pogoni, Oraiokastro, Kouri; 39.9970°N, 20.5620°E; 22 Jul. 2015; RT, YM, AP leg.

Distribution: A species recorded from Greece (Central Greece, Central Makedonia, Eastern Makedonia and Thraki, Ikaria, Ipeiros, Kefalonia, Kriti, Lefkada, Mytilini, Thessalia, Peloponnisos, Western Makedonia), Albania, Austria, Bosnia & Herzegovina, Bulgaria, Croatia, Czech Republic, Georgia, Hungary, Iran, Israel, Italy, Lebanon, Northern Macedonia, Moldova, Montenegro, New Zealand†, Romania, Serbia, Slovakia, Slovenia, Turkey, Ukraine. Its presence in Balearic Islands, Portugal, Spain, and Switzerland is dubious. † antmaps.org notes this species as Indoor Introduced.
Formicinae Camponotini Camponotus gestroi Emery, 1878 Materials examined: GREECE • 1 ☿; Zagori, Agios Minas, Ikonisma Agiou Athanasiou; 39.9490°N, 20.6622°E; 26 Jul. 2015; RT, KS leg. Distribution: A species recorded from Greece (Attiki, Central Greece, Central Makedonia, Chalki, Irakleia, Karpathos, Kastelorizo, Kefalonia, Kerkyra, Kos, Kriti, Naxos, Nera, Rodos, Symi, Peloponnisos, Zakynthos), Algeria, Armenia, Bulgaria, Cyprus, France, Iran, Iraq, Israel, Italy, Lebanon, Montenegro, Northern Macedonia, Morocco, Portugal, Serbia, Spain and Turkey.
Camponotus jaliensis Dalla Torre 1893 = Camponotus oertzeni var. jaliensis Dalla Torre 1893 Materials examined: GREECE • 1 ☿; Zagori, Aristi, Agios Athanasios church; 39.9328°N, 20.6744°E; 27 Jul. 2015; RT, KS leg. Distribution: A species recorded from Greece (Central Makedonia, Chios, Gyali, Karpathos, Kefalonia, Kriti), Cyprus, Israel and Turkey.
Camponotus samius Forel 1893 = Camponotus rubripes r. samius Forel [13] Materials examined: GREECE • 1 ☿; Zagori, Aristi, Agios Athanasios church; 39.9328°N, 20.6744°E; 27 Jul. 2015; RT, KS leg. Distribution: A species recorded from Greece (Attiki, Central Greece, Eastern Makedonia and Thraki, Nisyros, Rodos, Samos, Santorini, Western Greece), Afghanistan, Cyprus†, Bulgaria, Romania, Serbia†, Sicily† and Turkey. † antmaps.org notes this species as Needs Verification.
Camponotus vagus (Scopoli, 1763) = Formica vaga Scopoli [14] Materials Examined: GREECE
- 2 ☿; Pogoni, Vissiani, Abel monastery; 39.9457°N, 20.5494°E; 31 Jul. 2015; RT, KS leg.
- 1 ☿; Zagori, Fragades, Agios Christoforos church; 39.8194°N, 20.8619°E; 02 Aug. 2015; RT, KS leg.
- 1 ☿; Pogoni, Vasiliko, Agia Varvara; 39.9999°N, 20.6117°E; 22 Jul. 2015; RT, YM leg. Distribution: A species recorded from Greece (Attiki, Central Greece, Central Makedonia, Ipeiros, Kefalonia, Kerkyra, Kriti, Peloponnisos, Thessalia, West Makedonia), Albania, Algeria, Andorra, Austria, Balearic Islands, Belarus, Belgium, Bosnia & Herzegovina, Bulgaria, China (Xinhiang district), Croatia, Czech Republic, Denmark, Finland, France, Georgia, Germany, Hungary, India (Andhra Pradesh district), Israel†, Italy, Kazakhstan, Latvia, Liechtenstein, Lithuania, Luxembourg, Northern Macedonia, Montenegro, Morocco, Netherlands†, Norway, Poland, Portugal, Moldova, Romania, Russia (not Russia East), Serbia, Slovakia, Slovenia, Spain, Sweden, Switzerland, Tunisia, Turkey and Ukraine. † antmaps.org notes this species as Indoor Introduced.
Lasiini Lasius alienus (Foerster, 1850) (Figure 3) = Formica aliena [15] Materials Examined: GREECE • 1 ☿; Konitsa, Aidonochori, Aidonolaloussa; 40.0356°N, 20.6039°E; 03 May 2015; KS, RT, ΑΤ, DT leg.; ZMUA MYRM 2017–000110

Distribution: A species recorded from Greece (Attiki, Andros, Central Greece, Central Makedonia, Ipeiros, Kalymnos, Karpathos, Kos, Kriti, Mytilini, Peloponnisos, Rodos, Santorini, Thessalia, Western Makedonia), Albania, Algeria, Andorra, Armenia, Austria, Azerbaijan, Bahamas, Belarus, Belgium, Bosnia & Herzegovina, Bulgaria, Canada, Channel Islands, China, Croatia, Czech Republic, Denmark, Estonia, Finland, France, Georgia, Germany, Hungary, India (Jammu & Kashmir, Himachal Pradesh and Uttar Pradesh districts), Iran, Iraq, Ireland, Italy, Japan, Kazakhstan, Kyrgyzstan, Latvia, Lebanon, Lithuania, Luxembourg, Northern Macedonia†, Malta, Mexico, Moldova, Mongolia, Montenegro, Morocco, Netherlands, Norway, Pakistan, Poland, Portugal, Romania, Russia (Central, South and East) Serbia, Slovakia, Slovenia, South Korea, Spain, Sweden, Switzerland, Tajikistan, Tunisia, Turkmenistan, Turkey, Ukraine, United Kingdom, USA, Uzbekistan. Its presence in Israel and North Korea is dubious. † antmaps.org notes this species as Needs Verification. Lasius lasioides (Emery, 1869) (Figure 4) = Prenolepis fusculus [16]
Materials Examined: GREECE • 2 ☿; Konitsa, Aidonochori, Aidonolaloussa; 40.0350°N, 20.6026°E; 03 May 2015; KS, RT, ΑΤ, DT leg.; ZMUA MYRM 2017–000109

Distribution: A species recorded from Greece (Central Greece, Central Makedonia, Eastern Makedonia and Thraki, Ipeiros, Kefalonia, Thessalia, Western Greece), Albania, Algeria, Azerbaijan, Balearic Islands, Croatia, Cyprus, Egypt, France, Georgia, Iran, Israel, Italy, Malta, Montenegro, Morocco, Northern Macedonia, Portugal, Serbia, Slovenia, Spain, Syria, Tunisia, Turkey.
Lasius illyricus (Zimmermann, 1935) = Lasius alienus subsp. Illyricus [17] Materials examined: GREECE • 2♀, 7 ☿; Zagori, Elati, Agia Triada church; 39.8195°Ν, 20.7577°Ε; 02 Aug. 2015, RT, KS leg. Distribution: A species recorded from Greece (Central Makedonia, Eastern Makedonia and Thraki, Ipeiros, Kerkyra, Kriti, Peloponnisos), Croatia, Northern Macedonia, Ukraine. Notes: A species very similar to L. emarginatus but with distinct morphological characters (predominately the chaetotaxy of the scapes and hind tibia) separating the two species [18].
Prenolepis nitens (Mayr, 1853) (Figure 5) = Tapinoma nitens [19] Materials examined: GREECE • 1 ☿; Zagori, Elafotopos, Kri Panagias; 39.9037°N, 20.6579°E; 30 Apr. 2015; KS, VMG, ΑΚ, ThV leg.; ZMUA MYRM 2017–000114

Distribution: A species recorded from Greece (Attiki, Central Greece, Central Makedonia, Eastern Makedonia and Thraki, Ipeiros, Kerkyra, Mytilini, Peloponnisos, Thessalia, Western Greece, Western Makedonia), Albania, Austria, Bosnia and Herzegovina, Bulgaria, Croatia, Georgia, Hungary, Italy, Lebanon, Northern Macedonia, Montenegro, Romania, Russia (Central), Serbia, Slovakia, Slovenia, Turkey, Ukraine, United Kingdom†. Its presence in the USA is dubious. † antmaps.org notes this species as Needs Verification.
Myrmicinae Attini Pheidole pallidula (Nylander, 1849) sensu lato = Myrmica pallidula [20] Materials examined: GREECE • 5 ☿; Zagori, Aristi, Agios Athanasios church; 39.9328°N, 20.6744°E; 27 Jul. 2015; RT, KS leg. Distribution: A species recorded from Greece (Attiki, Andros, Central Greece, Central Makedonia, Chios, Ipeiros, Kalymnos, Karpathos, Kasos, Kefalonia, Kos, Kriti, Lefkada, Limnos, Milos, Mytilini, Naxos, Nisyros, Peloponnisos, Rodos, Samos, Santorini, Skiathos, Skyros, Syros, Telendos, Thessalia, Western Greece, Zakynthos), Albania, Algeria, Andorra, Afghanistan, Armenia, Austria, Azerbaijan, Balearic Islands, Bosnia & Herzegovina, Bulgaria, Croatia, Cyprus, Denmark‡, Egypt, France, Georgia, Gibraltar, Germany, Iran, Iraq, Israel, Italy, Kazakhstan, Kyrgyzstan, Lebanon, Libya, Macaronesia†, Northern Macedonia, Malta, Monaco, Montenegro, Morocco, Portugal, Romania, Saudi Arabia, Serbia, Slovenia, Socotra Archipelago, Spain, Syria, Switzerland, Tunisia, Turkey, Turkmenistan, Uzbekistan, Zimbabwe. Its presence in Brazil and the USA is dubious. † antmaps.org notes this species as Exotic for the archipelagos of the Canary Islands. ‡ antmaps.org notes this species as Indoor Introduced. Notes: A very well-established species (or species complex) in Greece and adjacent areas. It has been suggested by various authors Seifert, et al. [21, 22] that this taxon includes at least four species; three of which (P. pallidula, P. balcanica and P. koshewnikovi) being sympatric in the Balkans.
Crematogastrini Crematogaster schmidti (Mayr, 1853) (Figure 6) = Acrocoelia schmidti [19] Materials examined: GREECE
- 3 ☿; Zagori, Messovouni, Agios Charalampos; 39.9330°N, 20.6260°E; 01 May 2015; KS, RT, ΑΤ, DT leg.
- 2 ☿; Zagori, Kato Pedina, Agia Varvara; 39.8905°N, 20.6710°E; 01 May 2015; KS, RT, ΑΤ, DT leg.
- 1 ☿;
Zagori, Kato Pedina, Agioi Taxiarches; 39.8796°N, 20.6785°E; 01 May 2015; KS, RT, ΑΤ, DT leg.
- 1 ☿; Konitsa, Aidonochori, Aidonolaloussa; 40.0359°N, 20.6034°E; 03 May 2015; KS, RT, ΑΤ, DT leg.
- 1 ☿; Konitsa, Aidonochori, Aidonolaloussa; 40.0355°N, 20.6031°E; 03 May 2015; KS, RT, ΑΤ, DT leg.
- 2 ☿; Zagori, Kato Pedina, Megala Pournaria; 39.8796°N, 20.6701°E; 13 May 2015; KS, ΑΚ, ThV leg.
- 2 ☿; Zagori, Kato Pedina, Megala Pournaria, 39.8803°N, 20.6725°E; 13 May 2015; KS, ΑΚ, ThV leg.
- 6 ☿; Zagori, Agios Minas, Ikonisma Agiou Athanasiou; 39.9490°N, 20.6622°E; 26 Jul. 2015; RT, KS leg.
- 13 ☿; Zagori, Aristi, Agioi Taxiarches church; 39.9358°N, 20.6711°E; 26 Jul. 2015; RT, KS leg.
- 14 ☿; Konitsa, Drosopigi, Panagia church; 40.2181°N, 20.9184°E; 08 Aug. 2015; RT, KS leg.; ZMUA MYRM 2017–000112
- 8 ☿; Pogoni, Teriachi, Agios Prodromos; 39.9367°N, 20.4066°E; 12 Aug. 2015; RT, YM, AP, GM leg.
- 15 ☿; Pogoni, 1km SE of Stavroskiadi; 40.0263°N, 20.4442°E; 13 Aug. 2015; RT, YM, AP, GM leg.

Distribution: A species recorded from Greece (Attiki, Central Greece, Central Makedonia, Kalymnos, Kefalonia, Kerkyra, Kriti, Lefkada, Mytilini, Peloponnisos, Rodos, Samos, Santorini, Thessalia, Zakynthos), Algeria†, Afghanistan, Armenia, Austria, Azerbaijan, Bosnia & Herzegovina, Bulgaria, Croatia, Cyprus, Georgia, Hungary, Iran, Italy, Libya, Northern Macedonia, Montenegro, Morocco†, Romania, Serbia, Slovenia, Sudan, Syria, Tunisia†, Turkey, Turkmenistan, Ukraine. Its presence in France is dubious.
† antmaps.org notes this species as Needs Verification.
Stenammini Aphaenogaster subterranea (Latreille, 1798) (Figure 7) = Formica subterranea Materials examined: GREECE • 2 ☿; Zagori, Asprageloi, Asprageloi church; 39.8232°N, 20.7288°E; 02 Aug. 2015; RT, KS leg.; ZMUA MYRM 2017–000113

Distribution: A species recorded from Greece (Central Greece, Central Makedonia, Kerkyra, Peloponnisos, Thessaly, Thraki, Western Greece and Western Makedonia), Albania, Algeria, Andorra, Armenia†, Austria, Azerbaijan, Balearic Islands, Belgium, Bosnia & Herzegovina, Bulgaria, Croatia, Cyprus†, Czech Republic, France, Georgia, Germany, Hungary, Iran, Italy, Luxemburg, Malta†, Moldova, Montenegro, North Macedonia, Poland, Portugal†, Romania, Slovakia, Slovenia, South Russia, Spain, Switzerland, Tunisia, Turkey and Ukraine. Its presence in the United Kingdom is dubious. † antmaps.org notes this species as Needs Verification.
Identification
All ant specimens were identified in the lab with the use of dichotomous keys [23, 24] as well as comparing with records from the literature Borowiec, et al. [25, 26]. When available, we compared our samples with high quality images of the type specimens on the AntWeb website (http://www. antweb.org). Species ranges are provided by comparing data from the Antmaps website (http://antmaps.org/?), the AntWeb website (http://www.antweb.org) and data indexing of previously published works (CG and AL personal database).
The relationship of ant species with particular trees species is documented vividly in Figure 8. The results indicate that the mostly utilized tree species by ants was Quercus pubescens onto which 8 species were collected. Extrapolating the findings to the genus level, trees belonging to the genus Quercus were associated with 10 ant species and only 2 ant species (namely Lasius flavus and Aphaenogater subterranea) were not collected from their barks. This coincides to what was expected since Quercus species comprise 67,9% of all tree species censused. Between the two most abundant species (Liometopum microcephalum and Crematogaster schmidti), the former one was entirely present on Quercus species showing an almost obligatory relationship to that tree genus, while the later one demonstrated a wider association with a variety of tree species (6 genera in total). What is interesting to note is that although L. microcephalum is regarded as a territorial, behaviorally dominant species in one occasion it was found coinhabiting a Quercus cerris tree with Camponotus vagus with no apparent conflict between the two species [27, 28]. Crematogaster schmidti, although mostly uniquely occupying trees, in two instances it was present alongside another ant species (Camponotus gestroi and C. vagus). Finally, all Camponotus species collected in the present study were seen on trees alongside other ant species.
![Figure 8: The results indicate that the mostly utilized tree species by ants was Quercus pubescens onto which 8 species were collected. Extrapolating the findings to the genus level, trees belonging to the genus Quercus were associated with 10 ant species and only 2 ant species (namely Lasius flavus and Aphaenogater subterranea) were not collected from their barks. This coincides to what was expected since Quercus species comprise 67,9% of all tree species censused. Between the two most abundant species (Liometopum microcephalum and Crematogaster schmidti), the former one was entirely present on Quercus species showing an almost obligatory relationship to that tree genus, while the later one demonstrated a wider association with a variety of tree species (6 genera in total). What is interesting to note is that although L. microcephalum is regarded as a territorial, behaviorally dominant species in one occasion it was found coinhabiting a Quercus cerris tree with Camponotus vagus with no apparent conflict between the two species [27,28]. Crematogaster schmidti, although mostly uniquely occupying trees, in two instances it was present alongside another ant species (Camponotus gestroi and C. vagus). Finally, all Camponotus species collected in the present study were seen on trees alongside other ant species.](/fulltextimages/11611/fig_8.png)
Discussion
Although the ant fauna presented in this study has been previously collected from several regions around Greece, there are species that are collected for the first time from the administrative area of Ipeiros, namely Camponotus gestroi, C. jaliensis, and C. samius. Most of the species collected from the trees (e.g. Camponotus vagus, Crematogaster schmidti, Liometopum microcephalum) have been known to create strong dependencies especially with broad-leaved tree species, something that is verified also from our investigation. Nevertheless, species such as Aphaenogaster subterranea and Pheidole pallidula are not usually associated with arboreal activity. Although most sites where ants have been collected were isolated in the past, all are now surrounded by newly established dense forests or scrublands that are flourishing due to land abandonment that caused natural regeneration, as the research area is now one of the most unpopulated areas in Greece.
We also need to note that the present work did not utilize a full array of collection techniques that would probably yield a larger number of species associated with the sacred trees surveyed. We acknowledge that most likely genera within the Ponerinae subfamily, as well as species of the Temnothorax, Tetramorium or Plagiolepis genera would probably be recorded if collection techniques such as sifting and beating were used, but the techniques utilized in this survey were to record the most dominant species associated with the trees in question.
Though preliminary, our findings highlight the often- neglected value of monumental, scattered trees in terms of biodiversity and species conservation [29, 30, 31]. In a future step, disentangling the phylogenetic relationships within and among species would greatly contribute towards understanding the currently observed distribution patterns. Moreover, questions regarding community ecology, synecology, coinhabiting preferences of certain species and distribution according to specific old growth characteristics of tree species and surrounding vegetation history could also be examined to explain patterns and preferences of myrmecofauna found in Ipeiros (Northwest Greece).
Acknowledgements
We would like to thank our colleagues Valentino Marini Govigli, Aris Kyparissis, Theofilos Vanikiotis (University of Ioannina), Jennifer L. G. Wong (Wild Resources Limited UK), Yannis Mantelis, Giorgos Margiolas, Alexis Pailas (Forestry Service of Ioannina), Artemis Tsiakiri and Dafni Tsiakiri for their help during specimen collection. CG extends his gratitude towards Anastasios Vaiou in helping curate and do the initial identifications of the specimens. We would like to sincerely thank Haris Prinianakis (https://www.flickr.com/ photos/harisprin/) for the production of the ant photos. We would like to finally give thanks to an anonymous reviewer for useful comments for the final version of this manuscript.
Funding
Part of the specimens were collected during field work in the frame of the project University of Ioannina-THALIS/ SAGE: - “Conservation through religion. The sacred groves of Epirus” co-financed by the European Union (European Social Fund – ESF) and Greek national funds through the “Operational Program of Education and Lifelong Learning of the National Strategic Reference Framework (NSRF) - Research Funding Program: THALIS”. Investing in knowledge society through the European Social Fund.
Authors’ Contributions
DNA, RT and KS developed the research idea that resulted in the article. RT and KS participated in the collection of specimens. CG curated and made the identifications of the specimens. AL provided data from his personal collection. CG, DNA and KS wrote the text. AL reviewed and provided valuable comments on the manuscript. All authors read, made suggestions and accepted the last version of the manuscript. The authors have declared that no competing interests exist.
References
-
Lagopoulos AF (2002) O Ouranos Pano sti Gi. Teletourgies Kathagiasis tou Ellinikou Paradosiakou Oikismou kai Proelefsi tous (The Sky Over the Earth. Sanctity Ceremonies in Greek Traditional Settlements and their Origin). Odysseas Editions, Athens [in Greek], pp: 303.
-
Nixon L (2006) Making a Landscape Sacred. Outlying Churches and Icon Stands in Sphakia, Southwestern Crete. Oxbow Books, Oxford, pp: 196.
-
Stara K, Tsiakiris R, Wong JLG (2012) Sacred trees and groves in Zagori, Northern Pindos National Park, Greece. In: Pungetti G, Oviedo G, et al. (Eds.), Sacred Species and Sites. Advances in Biocultural Conservation. Cambridge University Press, Cambridge, pp: 392-396.
-
Stara K, Tsiakiris R, Wong JLG (2015) The trees of the Sacred Natural Sites of Zagori, NW Greece. Landscape Research 40 (7): 884-904.
-
Tsiakiris R, Gribilakou L, Betsis A, Rubas E, Stara K, et al. (2017) Can inhabitants’ testimonies about Zagori and Konitsa sacred forests protective function be confirmed by spatial predictive models? Proceedings of the 18th Hellenic Forestry Congress/International workshop: Information Technology, Sustainable Development, Scientific Networks & Nature Protection. Hellenic Forestry Association, Thessaloniki, pp: 1080-1087.
-
Stara K, Tsiakiris R, Nitsiakos V, Halley JM (2016) Religion and the management of the commons. The sacred forests of Epirus. In: Agnoletti M, Emanueli F (Eds.), Biocultural Diversity in Europe. Springer International Publishing pp: 283-302.
-
Korakis G, Kapsalis E, Tsiakiris R, Stara K, Halley JM, et al. (2015) Identification and composition of habitat types in eight sacred forests in the area of Konitsa and Zagori, Greece. Proceedings of the 17th Panhellenic Forestry Congress. Argostoli, pp: 790-796.
-
Rackham O (2014) Sacred Natural Sites: Sacred Forests in Epirus. A Preliminary Report by Oliver Rackham 14. Report to Thalis-SAGE project, University of Ioannina, Greece, pp: 19.
-
Byers BA, Cunliffe R, Hudak AT (2001) Linking the conservation of culture and nature: a case study of sacred forests in Zimbabwe. Human Ecology 29(2): 187-218.
-
Avtzis DN, Stara K, Sgardeli V, Betsis A, Diamandis S, et al. (2018) Quantifying the conservation value of Sacred Natural Sites, Biological Conservation 222: 95-103.
-
Panzer GWF (1798) Fauna Insectorum Germanicae Initia, Oder Deutschlands Insecten. Heft 54. Felssecker, Nürnberg, pp: 24.
-
Dalla Torre KW Von (1893) Catalogus Hymenopterorum Hucusque Descriptorum Systematicus. Formicidae (Heterogyna). 7: 289.
-
Forel A (1889) Ameisen aus den Sporaden, den Cykladen und Griechenland, gesammelt 1887 von Herrn von Oertzen. Berliner Entomologische Zeitschrift 32: 255- 265.
-
Scopoli JA (1763) Entomologia Carniolica Exhibens Insecta Carnioliae Indigena et Distributa in Ordines, Genera, Species, Varietates. Methodo Linnaeana. J Trattner Vindobonae 36: 420.
-
Foerster A (1850) Hymenopterologische Studien. 1. Formicariae. Ernst Ter Meer, Aachen, pp: 74.
-
Emery C (1878) Catalogo delle formiche esistenti nelle collezioni del Museo Civico di Genova. Parte seconda. Formiche dell’Europa e delle regioni limitrofe in Africa e in Asia. Annali del Museo Civico di Storia Naturale 12: 43-48.
-
Zimmermann S (1935) Beitrag zur Kenntnis der Ameisenfauna Süddalmatiens. Verhandlungen der Zoologisch-Botanischen Gesellschaft in Wien 84: 1-65.
-
Borowiec L, Salata S (2013) Ants of Greece – additions and corrections (Hymenoptera: Formicidae). Genus 24: 335-401.
-
Mayr G (1853) Einige neue Ameisen. Verhandlungen der Zoologisch-Botanischen Vereins in Wien 2: 143-150.
-
Nylander W (1849) Additamentum alterum adnotationum in monographiam formicarum borealium. Acta Societatis Scientiarum Fennicae 3: 25-48.
-
Seifert B (2016) Inconvenient hyperdiversity – the traditional concept of “Pheidole pallidula” includes four cryptic species. Soil Organisms 88 (1): 1-17.
-
Bračko G, Kiran K, Karaman C, Salata S, Borowiec L (2016) Survey of the ants (Hymenoptera: Formicidae) of the Greek Thrace. Biodiversity Data Journal (4): e7945.
-
Bračko G, Lapeva-Gjonova A, Salata S, Borowiec L, Polak S (2019) Aphaenogaster illyrica, a new species from the mountains of the Balkan Peninsula (Hymenoptera, Formicidae), ZooKeys 862: 89-107.
-
Seifert B (1992) A taxonomic revision of the Palaearctic members of the ant subgenus Lasius s. str. (Hymenoptera, Formicidae). Abhandlungen und Berichte des Naturkundemuseums Görlitz 66: 1-67.
-
Borowiec L, Salata S (2018) New records of ants (Hymenoptera: Formicidae) from Epirus, Greece. Acta Entomologica Silesiana 26(001): 1-22.
-
Salata S, Borowiec L (2018) Taxonomic and faunistic notes on Greek ants (Hymenoptera: Formicidae). Annals of the Upper Silesian Museum in Bytom, Entomology 27(008): 1-51.
-
Petráková, L, Schlaghamerský J (2011) Interactions between Liometopum microcephalum (Formicidae) and other dominant ant species of sympatric occurrence. Community Ecology 12(1): 9-17.
-
Tartally A (2006) Long term expansion of a supercolony of the invasive garden ant Lasius neglectus (Hymenoptera: Formicidae). Myrmecologische Nachrichten 9: 21-25.
-
Fabricius JC (1782) Species Insectorum Exhibentes Eorum Differentias Specificas, Synonyma, Auctorum Loca Natalia, Metamorphosin Adiectis Observationibus, Descriptionibus. Tome I. C. E. Bohn, Hamburgi et Kilonii pp: 552.
-
Forel A (1913) Fourmis de la faune méditerranéenne récoltées par MM. Sahlberg. Revue Suisse de Zoologie 21: 427-438.
-
Seifert B (2007) Die Ameisen Mittel- und Nordeuropas. Lutra Verlags- und Vertriebsgesellschaft, Görlitz/Tauer, pp: 368.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind