Inhibition and Activation of Humoral Phenoloxidase and Their Effect on Divalent Cations in the Hemolymph of Freshwater Prawn Macrobrachium rosenbergii (De Man, 1879)
The aim of the present study is to examine phenoloxidase (PO), a copper-containing enzyme that plays a crucial role in the immune responses of arthropods, especially crustaceans and insects. However, PO activity is present in the plasma of the hemolymph without pathogenic infections. Here we report the PO activity in the plasma of M. rosenbergii was spectrophotometrically studied. The phenylthiourea (PTU) inhibition of PO activity was found to be the highest at a 10 mM concentration. The determination of PO activity was also highest activated by trypsin (2 mg.ml-1), detergents like sodium dodecyl sulphate (SDS; 8 mg.ml-1) and laminarin (4 mg.ml-1) enzyme expression. The effect of divalent cations on PO activity like sodium chloride (100 mM), calcium chloride, magnesium chloride, manganese chloride, and potassium chloride (10 mM) affected the activation of PO activity in trypsin, SDS, and laminarin. However, these divalent cations only slightly affected PTU inhibition of PO activity. In addition to calcium chloride, the PO activity with trypsin and laminarin is more or less the same as with control, but a great enhancement was found in SDS, which indicates that the combination of SDS with calcium chloride is required for increasing the PO activity in M. rosenbergii. The concentration of ethylenediaminetetraacetic acid (EDTA) of 1–10 mM has the maximum inhibition in PTU and activation of trypsin, SDS, and laminarin, which indicates that EDTA is a divalent cation chelator that affects the PO activity in M. rosenbergii. Hence, an attempt has been made to optimize the conditions for inhibitor, activators, divalent cations, EDTA sensitivity, and phenoloxidase activity in the plasma of the freshwater prawn M. rosenbergii.
Introduction
The giant freshwater prawn, M. rosenbergii (De Man, 1879) is an economically important crustacean, which is farmed on a large scale in many different countries. This species lives in tropical freshwater environments influenced by adjacent brackishwater areas. It is often found in extremely turbid conditions [1]. They are well known for their fine delicate flavour and tasty flesh. They also hold an outstanding market value in domestic as well as international markets and their economic value increases as they grow bigger in size [2]. Although low in production quantity, some of the smaller product groups, such as shrimp and marine fish have an inadequate economic significance because of their high unit value. Freshwater aquaculture contributes to over 95% of the total aquaculture production. Freshwater finfish, especially Chinese and Indian carp species, account for the largest share of total aquaculture production, followed by molluscs [1]. The M. rosenbergii culture was introduced and developed in India around 8 to10 years ago in compensation for the substantial losses of marine shrimp due to the epidemic white spot syndrome, assuming the resistance of the giant freshwater prawn to white spot disease [3].
The immune system is generally divided into two major types namely innate or natural and adaptive or acquired immunity. The innate immune system is further divided into humoral and cellular defense responses. In arthropods, the external cuticle is the first line of defense that provides both effective physical and chemical barriers against the attachment and entry of unwanted pathogens and parasites [4, 5]. The digestive tract is the main route of invasion and is partly lined with chitinous membranes, and the presence of acids and enzymes in the tract can inactivate and digest many viruses and bacteria [6]. In most cases, the cuticular defenses are enough to protect against virulent pathogens, disease occurs when the pathogens penetrate through the integument by physically damaging the cuticle [7]. Once pathogens gain entry into the haemocoel of the host, they encounter an intricate system of innate defense mechanisms that involve cellular and humoral responses [8]. The defense mechanisms of crustaceans depend completely on the innate immune system, due to the lack of immunoglobulin and memory [9]. The innate immune system is activated when pathogen-associated molecular patterns are recognized by soluble or by cell surface host proteins [10], such as lectins [11, 12], antimicrobial [13], agglutination [14], clotting [15], and pattern recognition proteins which in turn activate cellular or humoral effector mechanisms to destroy invading pathogens [16, 17, 18, 19, 20, 21].
Innate immune responses include nodulation [22], synthesis of antimicrobial peptides (AMPs) [23], encapsulation, phagocytosis [24] and melanization by the activation of proteolytic cascades [25], blood coagulation [26], opsonization which is assumed to be due to the release of stress-responsive proteins and molecules and iron sequestration [7]. Through the action of phenoloxidase, the formation of nodules and capsules are often observed to become melanized [27]. In invertebrates, physical injuries or the presence of foreign objects such as parasites and microorganisms result in melanin deposition around the damaged tissue or foreign objects like parasites [28]. The melanin will physically shield an intruder and therefore prevent or hinder its growth, and more importantly during melanin formation; highly reactive and toxic quinone intermediates are produced [29]. The active form of the enzymes phenoloxidase (PO) is responsible for these activities [9, 17, 20]. Phenoloxidase after activation will catalyze the initial steps in the pathway to melanin formation [5, 30]. Phenoloxidase does not only involve in melanin formation it is also involved in sclerotization and wound healing [31].
Phenoloxidase is synthesized as an inactive zymogen, prophenoloxidase (proPO) which can be activated by specific proteolysis [28]. When insects are infected by pathogens, activation of proPO is elicited by microbial cell surface components, such as lipopolysaccharide (LPS), peptidoglycans, β–1,3–glucose [32]. The proPO activation is undertaken by specialized PO activating enzymes, which themselves are activated by a cascade of serine proteases [33].
The key enzyme of melanization is PO or tyrosinase; monophenol, ortho–dihydroxyphenylalanine: oxygen oxidoreductase, which contains two copper atoms at its active site. This enzyme has three forms: “met” (with Cu(II)–Cu(II) in the active site), “deoxy”(Cu(I)– Cu(I)), and “oxy”(Cu(II)–O2 – Cu(II)) [34, 35]. The active enzyme is a bifunctional enzyme that will catalyze the o–hydroxylation of monophenols to catechols and the oxidation of o-diphenols into o-quinones [36]. The first step in the biosynthesis pathway from tyrosine to melanin involves the monophenolase activity in which conversion of L–tyrosine to L–DOPA (L-3,4–dihydroxyphenylalanine) takes place. This is then oxidized to L–DOPA quinone by phenoloxidase diphenolase activity [36, 37]. The quinones are then converted to melanin by both enzymatic and non– enzymatic reactions [15, 39] in one of these reactions; dopachrome is decarboxylated by dopachrome isomerase to form dihydroxy indole, which is then converted into melanin [39, 40, 41].
Since the 1940s, PTU was used as a PO inhibitor. It was observed that upon different phenols oxidation catalyzed by PO, the oxygen consumption decreased noticeably in the presence of PTU [42, 43]. After a series of thiourea derivatives were investigated, PTU was found to be the most effective melanization inhibitor [44]. The action of PTU is based on its interaction with copper ions at the active site of PO [45, 46]. However, some of the authors suppose that PTU is the noncompetitive inhibitor of PO [47].
Phenoloxidase activity can also be induced by exogenous reagents such as detergents SDS or cetylpyridinium chloride (CPC), isopropyl alcohol, or proteases [17, 48]. Whereas, there is a report that anticoagulants suppressed PO activity in certain arthropods. A cane sugar factor used as an anticoagulant suppressed PO activity in the hemolymph of the silkworm Bombyx mori [49]. Furthermore, there is evidence shown in cockroach Blaberus craniifer that the use of another anticoagulant, citrate-EDTA buffer, inactivated about 50% of the total PO activity in hemocyte lysate supernatant even after removal of anticoagulant by dialysis [50].
Trypsin is able to greatly enhance PO activity. The stimulation of PO activity by trypsin which is a serine protease, suggests that PO may be present in the acellular fraction of hemolymph in the form of a pro-enzyme and required a proteolytic cleavage for activation [17, 51]. A proteinaceous molecule has been purified with a specific affinity to β-1,3- glucan and the ability to make the prophenoloxidase activating system reactive to β-1,3-glucan [17, 52]. A protein known as β-glucan binding protein (BGBP) reacts with β-glucans. Then the glucan β-GBP complex induces degranulation and the activation of prophenoloxidase (proPO). This protein is highly conserved in all and is present in all the crustaceans studied so far. Shrimp β-GBP appears to be a constitutive plasma protein that after binding to β-glucans reacts with hemocyte surface and stimulates the release of hemocytic granules. The contents of the granules become activated in presence of plasma Ca²⁺ which leads to PO activity. Clearly, the activating reaction does not occur only in the presence of β-GBP, although its interaction with the hemocyte membrane could exist [53].
It was found that the PO activity was dependent on the ions calcium and magnesium. There is a specific mM concentration requirement of calcium and magnesium i.e. 5 mM and 10 mM respectively that could enhance the PO activity, but if this concentration is increased inhibition of enzyme activity occurs. Furthermore, the simultaneous addition of both cations (5 mM of CaCl2 and 10 mM MgCl2) induced a higher enzyme activity (60% approximately) than if each cation was added separately [54].
The EDTA inhibits PO activity; this inhibition of PO activity with EDTA indicates the involvement of divalent cations in the melanin-synthesis pathway [55, 56, 57]. In fact, the effects of EDTA may be due to the calcium dependency of the POs, as calcium is known to increase the activity of several invertebrate POs [58], which suggests that the binding of some calcium atoms is necessary for the activating center of PO [59]. Hence, an attempt has been made to optimize the conditions for inhibitor, activators, divalent cations and EDTA sensitivity phenoloxidase activity of plasma in freshwater prawn M. rosenbergii.
Materials and Methods
Collection and Acclimatization of Experimental Prawns
The freshwater prawn M. rosenbergii weighing around 25-30 ± 2.12 g was collected from Aqua Farm, Kalpakkam, Kanchipuram District, Tamil Nadu, India. In the laboratory, the prawns were maintained in plastic tanks containing continuously aerated freshwater, which was changed twice once. This ensures sufficient oxygen supply to the prawns and an environment devoid of accumulated metabolic wastes. The animals were fed with egg white ad libitum and were acclimatized to the laboratory conditions for one week prior to use. The uninjured, intermoult animals were used throughout this study.
Hemolymph Collection and Preparation of Plasma
The preparation of plasma follows Sivakumar, et al. [17] and is briefly explained. Hemolymph (100 µl) was collected by cardiac puncture using a 23G needle attached to a clean, sterile plastic syringe containing 1.9 ml of ice-cold iso-osmotic buffer (TBS I: Tris-50 mM, NaCl-210 mM, KCl-5 mM, MgCl2-2.5 mM; pH 7.5). Rapidly the syringe was gently rotated to enable thorough mixing of the hemolymph sample with iso-osmotic buffer and immediately transferred to a pre-chilled polypropylene tube, centrifuged (161 x g, 8 min at 4 °C) to obtain 1.5 ml of the supernatant as plasma. The exclusion of hemocytes was verified in the collected plasma by observation under a phase contrast microscope. About 50 prawns (N = 50) were required for the collection of 100 µl acellular plasma for each determination in this study.
Determination of Protein Concentration
The protein content in the experimental samples was determined according to Bradford [60] using bovine serum albumin as the standard. All chemicals used in the study were purchased from Sigma-Aldrich, St. Louis, MO, USA.
Phenoloxidase Activity by Plasma
The PO activity of plasma with the phenolic substrate was tested by incubating 100 µl of plasma with 1.9 of substrate solution (2.5 mM L-DOPA) for 15 min at 25 °C in a spectrophotometer for plasma. The colour developed was measured spectrophotometrically at 470 nm against a blank reagent of L-DOPA [20].
Effect of Inhibitor on PO Activity
In this experiment, 100 μl of plasma was separately mixed with 100 μl of 1-10 mM PTU. In control, these chemicals were substituted with Tris-HCl buffer (10 mM; pH 7.5) and pre-incubated for 15 min at 25 °C. An aliquot of 200 μl of the reaction mixture from control or experiments was incubated with made up to 2 ml of 2.5 mM L-DOPA for 15 min at 25 °C. The optical density of both control and experiments was measured spectrophotometrically at 470 nm against a blank reagent of L-DOPA.
Effect of Activators on PO Activity
In this experiment, 100 μl of plasma was separately mixed with 100 μl of 1-10 mg.ml-1 of trypsin, SDS and laminarin. In control, these chemicals were substituted with Tris-HCl buffer (10 mM; pH 7.5) and pre-incubated for 15 min at 25°C. An aliquot of 200 μl of the reaction mixture from control or experiments was incubated with made up to 2 ml of 2.5 mM L-DOPA for 15 min at 25°C. The optical density of both control and experiments was measured spectrophotometrically at 470 nm against a blank reagent of L-DOPA.
Effect of Sodium Chloride on PO Activity
In this experiment, 100 μl of plasma was separately mixed with 100 μl of PTU (7.5 mM), Trypsin (2 mg.ml-1), SDS (8 mg.ml-1) and laminarin (4 mg.ml-1). In control, these chemicals were substituted with TBS II (Tris-10 mM, NaCl-100 mM; pH 7.5), and pre-incubated for 15 min at 25°C. An aliquot of 200 μl of the reaction mixture from control or experimentals were pre-incubated with made up to 2 ml of 2.5 mM L-DOPA for 15 min at 25°C. The optical density of both control and experiments was measured spectrophotometrically at 470 nm against a blank reagent of L-DOPA.
Effect of Divalent Cations on PO Activity
In this experiment, 100 μl of plasma was separately mixed with 100 μl of PTU (7.5 mM), Trypsin (2 mg.ml-1), SDS (8 mg.ml-1) and laminarin (4 mg.ml-1). In control, these chemicals were substituted with TBS III, IV, V and VI (Tris-10 mM, NaCl-100 mM, CaCl2-10 mM or MgCl2-10 mM or MnCl2- 10 mM or KCl-10 mM; pH 7.5), and pre-incubated for 15 min at 25°C. An aliquot of 200 μl of the reaction mixture from control or experimentals were pre-incubated with made up to 2 ml of 2.5 mM L-DOPA for 15 min at 25°C. The optical density of both control and experiments was measured spectrophotometrically at 470 nm against a blank reagent of L-DOPA. Effect of EDTA Aensitivity on PO Activity In this experiment, 100 μl of plasma was separately mixed with 100 μl of PTU (7.5 mM), Trypsin (2 mg.ml-1), SDS (8 mg.ml-1) and laminarin (4 mg.ml-1). In control, these chemicals were substituted with TBS VII-X (Tris-10 mM, NaCl-100 mM, EDTA 1.0, 2.5, 5.0 and 10 mM; pH 7.5), and incubated for 15 min at 25°C. An aliquot of 200 μl of the reaction mixture from control or experimentals were pre- incubated with made up to 2 ml of 2.5 mM L-DOPA for 15 min at 25°C. The optical density of both control and experiments was measured spectrophotometrically at 470 nm against a blank reagent of L-DOPA.
Statistical Analysis
The data were expressed as mean ± SD of triplicate experiments from five determinations. The variation between experimental and control were evaluated by SPSS/18 software (version 20; SPSS, New York, USA) by one-way analysis of variance (ANOVA) at 0.05, 0.01, 0.001 probability (p < 0.05*, 0.01, 0.001*).
Results
PO Activity of Plasma and Effect of Inhibition on PTU
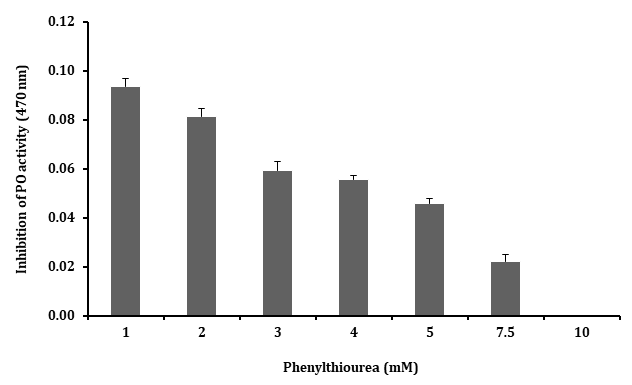
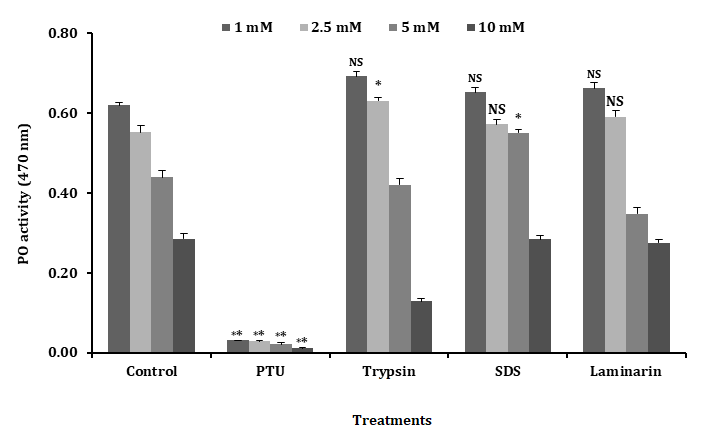
The plasma separated from the hemolymph of the freshwater prawn M. rosenbergii, oxidized the diphenolic substrate L-DOPA with an activity (Figure 1), at optimal conditions tested including 2.5 mM L-DOPA, 10 mM Tris- HCl, 25°C, pH 7.5 for 15 min at 470 nm (20). The plasma PO activity inhibition was tested with different mM concentrations of PTU (1 - 10 mM) and the PO activity was found to be completely inhibited with 10 mM concentration than that of 1 - 7.5 mM i.e. the lower concentrations as shown in Figure 1. These results clearly suggested that the optimum concentration for inhibiting the PO activity is 10 mM of PTU.
Effect of Trypsin Activation on PO Activity
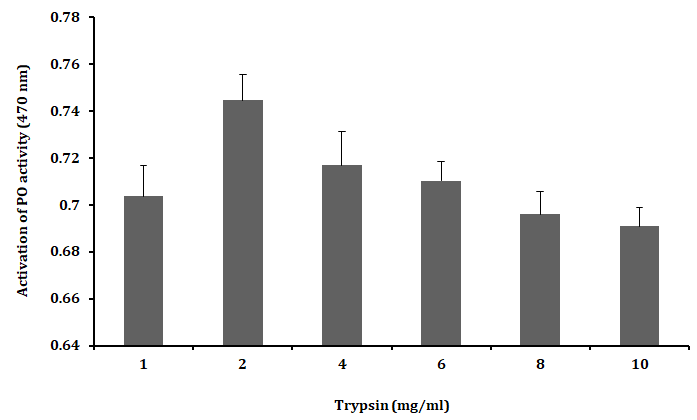
The plasma PO activity was tested with different concentrations of trypsin (1 - 10 mg.ml-1) and the PO activity was found to be higher with trypsin of 2 mg.ml-1 concentration than that of 1 mg.ml-1 concentration or higher concentrations (4, 6, 8 and 10 mg.ml-1) as shown in Figure 2. However, these results clearly suggested the optimum concentration for activating PO activity in 2 mg.ml-1 of trypsin.
Effect of SDS Activation on PO Activity
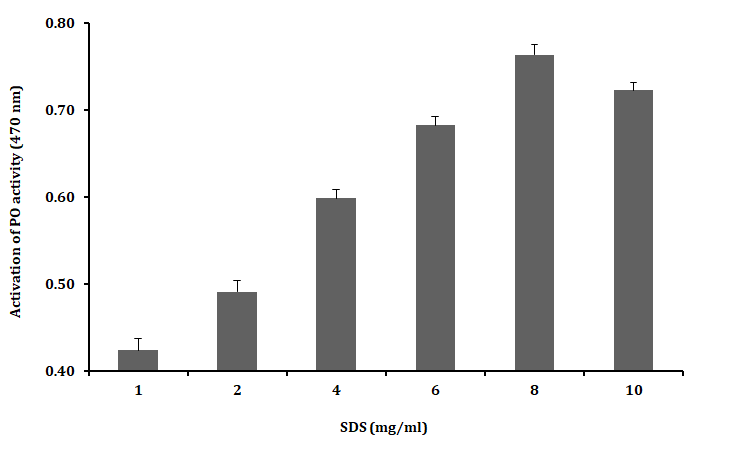
The plasma PO activity was tested with different concentrations of SDS (1 - 10 mg.ml-1) and the PO activity was found to be higher with SDS of 8 mg.ml-1 concentration than that of 1 - 6 mg.ml-1 concentration or higher concentrations (10 mg.ml-1) as shown in Figure 3. However, these results clearly suggested that the optimum concentration for activating PO activity in 8 mg.ml-1of SDS.
Effect of Laminarin Activation on PO Activity
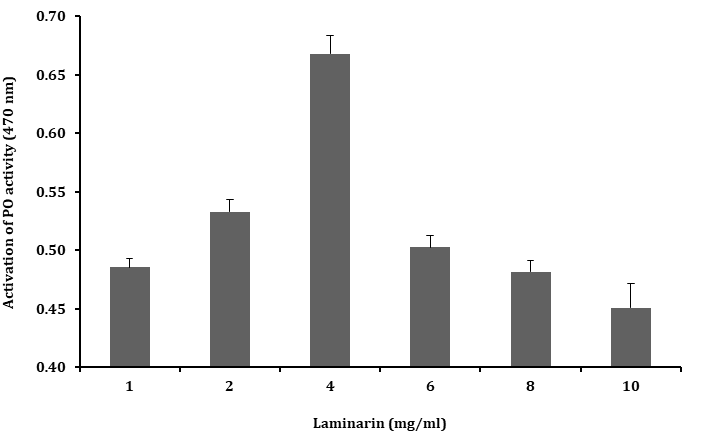
The plasma PO activity was tested with different concentrations of laminarin (1 - 10 mg.ml-1) and the PO activity was found to be higher with laminarin of 4 mg.ml-1 concentration than that of 1 - 2 mg.ml-1 concentration or higher concentrations (6, 8 and 10 mg.ml-1) as shown in Figure 4. However, these results clearly suggested that the optimum concentration for activating PO activity is 4 mg.ml-1 of laminarin.
Effect of Divalent Cations on Phenoloxidase Activity
• Effect of sodium chloride on PO activity The plasma PO activity was tested with TBS-II which is a divalent cation with PTU, the PO activity was found to be inhibited by PTU when compared to control, and the reduction was found to be about 97.76% (p < 0.001). Similarly, the pre-treatment of plasma with trypsin, SDS and laminarin showed PO activity, and there was no significant activation in the oxidation of L-DOPA as compared to control (Figure 5A).
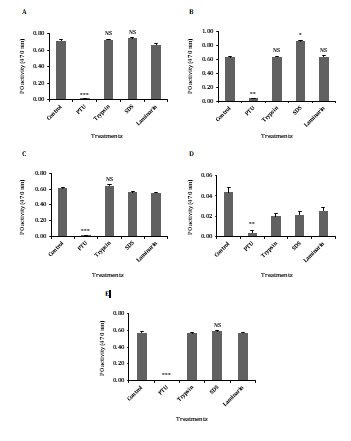
Figure 5: Effect of divalent cations using (A) sodium chloride, (B) calcium chloride, (C) magnesium chloride, (D) manganese chloride and (E) potassium chloride on phenoloxidase activity of plasma in the freshwater prawn M. rosenbergii Asterisk indicates significant variation determined from the value obtained for a treatment to untreated control by one-way ANOVA at *p < 0.05, p < 0.01, *p < 0.001.
- Effect of calcium chloride on PO activity The plasma PO activity was tested with TBS-III which is a divalent cation with PTU, the PO activity was found to be strongly inhibited (93.38%) by PTU (p < 0.001). Activation of PO activity was significantly (p < 0.05) increased in SDS when compared to control and the activation was found to be 26.25%. The PO activity was absent in trypsin and the laminarin has minimal activation but it is not statistically significant (Figure 5B).
- Effect of magnesium chloride on PO activity The plasma PO activity was tested with TBS-IV, which is a divalent cation with PTU, the PO activity was found to be strongly inhibited (98.21%) by PTU (p < 0.001). The activation of PO activity in trypsin was only slightly increased but was not statistically significant and no response was seen in the anionic detergent of SDS and polymer of laminarin compared to control. This result clearly suggests that the addition of magnesium chloride decreases the PO activity (Figure 5C).
- Effect of manganese chloride on PO activity The plasma PO activity was tested with TBS-V, which is a divalent cation with PTU, the PO activity was found to be strongly inhibited (90.90%) by PTU (p < 0.001) and the enormous decreased activity of PO is seen in trypsin, SDS and laminarin. These results clearly show that the addition of manganese chloride highly affected PO activity (Figure 5D).
- Effect of potassium chloride on PO activity The plasma PO activity was tested with TBS-VI, which is a divalent cation with PTU, the PO activity was found to be completely inhibited by PTU (p < 0.001). The activators and elicitor decreased the activity of PO, which suggested that the potassium chloride did not significantly enhanced PO activity when compared to the control (Figure 5E).
- Effect of EDTA sensitivity on PO activity The plasma PO activity was tested with EDTA of different concentrations with PTU, trypsin, SDS and laminarin, the PO activity was found to be above 95% inhibited by PTU of all the concentrations of EDTA. All the activators and elicitors decreased in the PO activity of 10 mM EDTA concentration.
No effects on activators were seen in 1, 2.5 and 5 mM concentrations. These results clearly suggested that the EDTA affected the plasma PO activity (Figure 6).
Discussion
The inhibition and activation of PO activity in plasma from the freshwater prawn M. rosenbergii have been described in the present study. However, the preponderance of the crustaceans PO is the terminal component of a cascade of proteolytic reactions that helps in the recognition of foreign materials in many crustaceans and insects [17, 20, 61]. The response of the entry of microbes and parasites is often visible as dark spots caused by melanin on the cuticle of arthropods, the enzyme responsible for the formation of melanin is phenoloxidase by catalyzing the oxidation of phenol to quinines that subsequently polymerize to melanin. Toxic metabolites with antimicrobial properties are formed during the formation of melanin [62].
It is well recorded that in arthropods, the zymogenic form, proPO is converted to PO through proteolytic cleavage by a serine protease such as trypsin [17, 28] and this activity can also be induced by exogenous reagents such as SDS [17, 48] and laminarin [17, 52]. The present studies optimize the condition for determining the activation of PO activity, its cation dependency and EDTA sensitivity of PO activity in plasma of M. rosenbergii. The substrate affinity and optimized condition (pH, ionic strength, temperature and time interval) of plasma PO were found to be a diphenol L-DOPA [20].
An inhibition study with the different concentrations of PTU was done in figure 1 the PO activity is almost entirely inhibited by PTU in the first six concentrations i.e. 1-7.5 mM and a complete inhibition in 10 mM was observed. The results clearly demonstrated 10 mM concentration of PTU is required for complete inhibition which may be due to the influence of PTU on this process which is caused by its interaction with active sites of PO rather than with intermediate products of DOPA oxidation preventing the melanin formation. The inhibition may be due to copper chelation by PTU since copper is essential for PO to function [51]. This clearly suggested that PO in the plasma of M. rosenbergii has copper in the active site and PTU, known as a chelating reagent of copper [17, 20, 63, 64].
The present study has also included the effect of different concentrations of trypsin, SDS and laminarin. Trypsin is a serine protease that can stimulate the activity of PO by enhancing the proteolytic cleavage of the pro-enzyme [51]. In the present study, the optimum concentration of trypsin for the activation of plasma PO activity in M. rosenbergii is 2 mg.ml-1. The different concentration shows different activation rate and the maximum activation was shown in 2 mg.ml-1. The same type of activation by trypsin was found in Erimacrus isenbeckii by the addition of 40 µM of trypsin [65]. The activation of PO by the addition of trypsin is the indication of the presence of PO as prophenoloxidase in M. rosenbergii and other invertebrate crustaceans [15, 66].
The incubation of SDS with plasma enhanced the activity of PO in M. rosenbergii. SDS activation does not occur at the same rate in different concentrations. A maximum PO activity was obtained with SDS concentration of 8 mg.ml-1 in plasma can be activated by SDS. This activation is due to a conformational change in the proenzyme in the form of distortion or slight unfolding of the proenzyme, this resulting conformational change should allow substrates to reach the active site [67]. The o-diphenolase activity of Cancer Pagurus hemocyanin on treatment with 4 mM SDS increases about twofold [68]. But in Limulus polyphemus no o-diphenolase activity was seen, but it exhibits a slight activity after incubation with SDS [69]. This activation by SDS is accompanied by changes in the secondary and tertiary structure [69]. The activation of PO activity is also found in Erimacrus isenbeckii with 5000 µM being its peaked concentration [65]. In contrast to this, no activity in hemocyanin of crayfish was detected when treated with SDS [70].
Laminarin or β-1,3 glucan can trigger the proPO activating system (Ochiai and Ashida, 1988). In the present study, the effect of different concentration of laminarin in the plasma of M. rosenbergii was found to be the highest in 4 mg.ml-1. Similar to what has been recorded in other crustaceans [30, 71, 72] proPO activation of M. rosenbergii was dose-dependent for laminarin. There is a β-glucan binding protein β-GBP which reacts with β-glucans, the glucan β-GBP complex thus formed induced the activation of proPO. This protein is present in all crustaceans studied so far and is highly conserved [53]. The β-GBP has been purified from three crustaceans, the freshwater crayfish Pacifastacus leniusculus [73], the yellow leg shrimp, Penaeus californiensis [74] and the white shrimp Penaeus vannamei [75]. The activation of PO activity by laminarin indicates the presence of β-GBP (β-glucan binding protein) in the plasma of M. rosenbergii.
In the present study, no regular trend was seen in all the five metal ions tested. The activities of PO were mostly increased by sodium chloride and calcium chloride, which is an indication that these metal ions played a positive role during the PO catalysis of M. rosenbergii. It was found that the addition of sodium chloride increased the activity of PO with trypsin and SDS and decreased laminarin and the highest activity was seen in SDS. Both activation by trypsin and SDS is minimal or negligible. The present study shows that calcium chloride and sodium chloride are essential to enhance PO activity in M. rosenbergii. In addition to calcium chloride, the activity with trypsin and laminarin is more or less the same as with control but a great enhancement was shown in SDS which indicates that the combination of SDS with calcium chloride is required for increasing the activity of PO in M. rosenbergii. PO activity in P. paulensis was considerably enhanced by the divalent ion calcium chloride (5 mM) and strongly inhibited by higher concentrations of this cation [54].
The magnesium chloride has both enhancements and decreased effects on PO activity, which indicates that it can play both positive and negative roles during the catalysis of PO. A significant increase in activity with trypsin was shown this clearly suggests that MgCl2 has a proper enhancement effect only with trypsin and decreased activity with SDS as well as laminarin. The same kind of decreased activity was recorded in Charybdis japonica, where PO activity was strongly inhibited by magnesium chloride [76]. On the other hand, the PO activity in P. paulensis was considerably enhanced by the divalent ion magnesium chloride (10 mM) and strongly inhibited by high concentrations of this cation [54].
Among all the divalent cations used, manganese chloride inhibits the PO activity and has drastically decreased the activity of PO with trypsin, SDS as well as laminarin. This clearly suggested that it played a negative role during the catalysis of PO in the plasma of M. rosenbergii. On the other hand, potassium chloride was found to have no effect on the PO activity in the plasma, even though a slight increase was shown only on SDS it can be considered as no effect. It can be concluded that potassium chloride has a negative effect on increasing the PO activity in M. rosenbergii. Some studies have suggested that the effect of metal ions is due to the modulation of PO by means of activating electrophile and nucleophile or changing the secondary structure of certain peptides of PO [77, 78].
The EDTA with a concentration of 10 mM has the maximum inhibition in PTU, and activation of trypsin, SDS and laminarin which indicates that EDTA is a divalent cation chelator that affects the activity of PO in M. rosenbergii. In the present study, smaller concentration does not have an effect on the PO activity which suggested that PO activity is dose-dependent. This kind of inhibition was also seen in L. vannamei suggested using EDTA-free anticoagulant in separating plasma for PO activity assay [79]. Likewise, in M. rosenbergii and Penaeus monodon the addition of 20 mM EDTA inhibits the PO activity [80].
Conclusion
In the present study, we can derive from the above observation is that, for the plasma phenoloxidase enzyme, the divalent cation studied, NaCl, CaCl2, MgCl2, MnCl2, KCl and EDTA sensitivity a little affected the inhibition and more affected activation of PO activity and the CaCl2 with SDS has enhanced the PO activity. However, the PO activity in plasma of M. rosenbergii the inhibition and activation determined are very distinct by showing specific concentrations. This finding proved plasma as a potent humoral component of the immune system with an ability to enzymatically function as PO in absence of cellular hemocytes and revealed catechol oxidase type PO. Conclusively, the study clearly indicates the significance of non-cellular immune components in boosting immune response. This finding provides evidence that the plasma of M. rosenbergii is a potent immune system with an ability to enzymatically function as phenoloxidase.
Conflict of Interest
The authors declare that they have no competing of interest.
References
-
FAO (2015) The State of World Fisheries and Aquaculture, Rome. Italy.
-
Venkatesh K, Rajesh P, Mukesh P, Arockiaraj J (2016) Impacts of environmental and biological stressors on immune system of Macrobrachium rosenbergii. Rev Aquac 9(3): 283-307.
-
Sahul Hameed AS, Xavier Charles M, Anilkumar M (2000) Tolerance of Macrobrachium rosenbergii to white spot syndrome virus. Aquaculture 183(3-4): 207-213.
-
Ashida M, Brey PT (1995) Role of the integument in insect defense: prophenoloxidase cascade in the cuticular matrix. Proceedings of Natural Academic Sciences, USA 92: 10698-10702.
-
Ashida M, Brey PT (1997) Recent advances on the research of the insect prophenoloxidase cascade, Molecular Mechanisms of Immune Responses in Insects. In: Brey P, Hultmark D (Eds.), London Chapman & Hall, UK, pp: 135-172.
-
Billingsley PF, Lehane MJ (1996) Structure and ultrastructure of the insect midgut. In: Lehane MJ, Billingsley PF, editors. Biology of the insect midgut. London: Chapman and Hall, UK, pp: 3-30.
-
Jiravanichpaisal P, Lee B, Söderhäll K (2006) Cell-mediated immunity in arthropods: Hematopoiesis, coagulation, melanization and opsonization. Immunobiology 211(4): 213-236.
-
Franssens V (2006) Study of two types of immune responses in insects: nodulation in the flesh fly, Neobellieria bullata, and prophenoloxidase activation in the desert locust, Schistocerca gregaria. Katholieke Universiteit Leuven, Thesis, pp: 1-117.
-
Lee SY, Söderhäll K (2002) Early events in crustacean innate immunity. Fish shellfish Immunol 12(5): 421-437.
-
García-Carreño FL, Cota K, Navarrete Del Toro MA (2008) Phenoloxidase activity of hemocyanin in whiteleg shrimp Penaeus vannamei, conversion, characterization of catalytic properties and role in postmortem melanosis. J Agric Food Chem 56(15): 6454-6459.
-
Marques MRF, Barracco MA (2000) Lectins, as non-self- recognition factors, in crustaceans. Aquaculture 191(1- 3): 23-44.
-
Denis M, Sivakumar MR, Tamilarasan K, Karthigayani T, Prabu M, Bhuvaneswari SD, et al. (2017) Activation of phenoloxidase activity by humoral lectin in hemocytes of freshwater crab Paratelphusa jacquemontii. Int J Biol Macromol 97: 258-263.
-
Mendoza HL, Faye I (1999) Physiological aspects of the immunoglobulin superfamily in invertebrates. Dev Comp Immunol 23(4-5): 359-374.
-
Sivakumar MR, Denis M, Shanthi S, Arumugam M (2020) Agglutination of plasma, hemocyanin, and separated hemocyanin from the hemolymph of the freshwater prawn Macrobrachium rosenbergii (De Man, 1879) (Decapoda: Caridea: Palaemonidae). J Crustac Biol 40(3): 309-315.
-
Sritunyalucksana K, Söderhäll K (2000) The proPO and clotting system in crustaceans. Aquaculture 191(1-3): 53-69.
-
Medzhitov R, Janeway CA (2002) Decoding the patterns of self and nonself by the innate immune system. Science 296(5566): 298-300.
-
Sivakumar MR, Denis M, Shanthi S, Arumugam M (2017) Phenoloxidase activity in humoral plasma, hemocyanin and hemocyanin separated proteins of the giant freshwater prawn Macrobrachium rosenbergii. Int J Biol Macromol 102: 977-985.
-
Shanthi S, Sivakumar MR, Rayvathy B (2021) Serum phenoloxidase activity in the hemolymph of the anomuran crab Albunea symmysta (Linnaeus, 1758) (Decapoda: Anomura: Albuneidae). J Crustac Biol 41(1): ruab003.
-
Shanthi R, Sivakumar MR (2022) Serum Phenoloxidase Activity in the Hemolymph of Giant Freshwater Prawn Macrobrachium rosenbergii (De Man, 1879). Int J Zoo Animal Biol 5(6): 000419.
-
Sivakumar MR, Shanthi R (2022) Plasma and hemocyanin phenoloxidase derived from the hemolymph of giant freshwater prawn Macrobrachium rosenbergii (De Man, 1879). Crustacea-Nano, Micro and Macro Studies pp: 1-20.
-
Sivakumar MR, Abisha Adline F, Shanthi R (2022) Hemocytes lysate supernatant derived phenoloxidase activity in the hemolymph of giant freshwater prawn Macrobrachium rosenbergii (De Man, 1879). Int J Zoo Animal Biol 5(5): 000403.
-
Baines D, DeSantis T, Downer RGH (1992) Octopamine and 5-hydroxyl tryptamine enhance the phagocytic and nodule formation activities of cockroach (Periplaneta americana) haemocytes. J Insect Physiol 38(11): 905- 914.
-
Lavine MD, Strand MR (2002) Insect hemocytes and their role in immunity. Insect Biochem Mol Biol 32(10): 1295-1309.
-
Söderhäll K (1999) Invertebrate immunity. Dev Comp Immunol 23(4-5): 263-266.
-
An C, Zhang M, Chu Y, Zhao Z (2013) Serine protease MP2 activates prophenoloxidase in the melanization immune response of Drosophila melanogaster. PLOS one 8(11): e79533.
-
Theopold U, Schmidt O, Söderhäll K, Dushay MS (2004) Coagulation in arthropods: defence, wound closure and healing. Trends Immunol 25(6): 289-294.
-
Iwanaga S, Lee BL (2005) Recent advances in the innate immunity of invertebrate animals. J Biochem Mol Biol 38(2): 128-150.
-
Cerenius L, Söderhäll K (2004) The prophenoloxidase- activating system in invertebrates. Immunol Rev 198: 116-126.
-
Masuda T, Otomo R, Kuyama H, Omoji K, Tonomoto M, et al. (2012) A novel type of prophenoloxidase from the kuruma prawn Marsupenaeus japonicus contributes to the melanization of plasma in crustaceans. Fish Shellfish Immunol 32(1): 61-68.
-
Söderhäll K, Cerenius L (1998) Role of the prophenoloxidase-activating system in invertebrate immunity. Curr Opin Immunol 10(1): 23-28.
-
Ratcliffe NA (1985) Invertebrate immunity a primer for the non-specialist. Immunol Lett 10(5): 253-270.
-
Mómen SA, Salem DAM, Barakat EMS, Salama MS (2012) Activation of prophenoloxidase during bacterial injection into the desert locust, Schistocerca gregaria. Int J Bioeng Life Sci 6(8): 291-300.
-
Marmaras VJ, Charalambidis ND, Zervas CG (1996) Immune response in insects: the role of phenoloxidase in defense reactions in relation to melanization and sclerotization. Arch Insect Biochem Physiol 31(2): 119- 133.
-
Jolley RL, Evans LH, Makino N, Mason HS (1974) Oxytyrosinase. J Biol Chem 249(2): 335-345.
-
Itoh S, Kumei H, Taki M, Nagatomo S, Kitagawa T, et al. (2001) Oxygenation of phenols to catechols by A (µ-η2: η2-Peroxo) dicopper (II) complex: mechanistic insight into the phenolase activity of tyrosinase. J Am Chem Soc 123(27): 6708-6709.
-
Yamamoto K, Yakiyama M, Fujii H, Kusakabe T, Koga K, et al. (2000) Expression of phenoloxidase mRNA during silkworm hemocyte development. Biosci Biotechnol Biochem 64(1): 1197-1202.
-
Bai G, Brown JF, Watson C, Yoshino TP (1997) Isolation and characterization of phenoloxidase from egg masses of the gastropod mollusc, Biomphalaria glabrata. Comp Biochem Physiol B 118(2): 463-469.
-
Fang J, Han Q, Johnson JK, Christensen BM, Li J (2002) Functional expression and characterization of Aedes aegypti dopachrome conversion enzyme. Biochem Biophys Res Commun 290(1): 287-293.
-
Sugumaran M (1996) Roles of the insect cuticle in immunity. In: Söderhäll K, Iwanaga S, et al. (Eds.), New Directions in Invertebrate Immunology. SOS Publications, Fair Haven, pp: 355-374.
-
Shelby KS, Adeyeye OA, Okot-Kotber BM, Webb BA (2000) Parasitism-linked block of host plasma melanization. J Invertebr Pathol 75(3): 218-225.
-
Olivares C, Jimenez-Cervantes C, Lozano JA, Solano JC, Borron G (2001) The 5, 6-dihydroxyindole-2-carboxylic acid (DHICA) oxidase activity of human tyrosinase. Biochem J Rev 354(1): 131-139.
-
Bernheim F, Bernheim MLC (1942) Тhe action of phenylthiocarbamide on tyrosinase. J Biol Chem 145(1): 213-217.
-
Ryazanova AD, Alekseev AA, Slepneva IA (2012) The phenylthiourea is a competitive inhibitor of the enzymatic oxidation of DOPA by phenoloxidase. J Enzyme Inhib Med Chem 27(1): 78-83.
-
Dubois KP, Erway WF (1946) Studies on the mechanism of action of thiourea and related compounds. II. Inhibition of oxidative enzymes and oxidations catalyzed by copper. J Biol Chem 165(2): 711-721.
-
Benjakul S, Visessanguan W, Tanaka M (2005) Properties of phenoloxidase isolated from cephalothorax of kuruma prawn (Penaeus japonicus). J Food Biochem 29(5): 470- 485.
-
Garcia-Gil DMF, Lanz-Mendoza H, Hernandez-Herhandez FC (2007) Free rаdical generation during the activation of hemolymph prepared from the homopteran Dactylopius coccus. Archives of Insect Biochemistry Physiology 65: 20-28.
-
Rescigno A, Sollai F, Pisu B, Rinaldi A, Sanjust E (2002) Tyrosinase inhibition: general and applied aspects. J Enzyme Medicine Chem 17(4): 207-218.
-
Kanost MR, Gorman MJ (2008) Phenoloxidase in insect immunity. In: Beckage NE (Ed.), Insect immunity, New York: Academic, pp: 69-96.
-
Ashida M, Ishizaki Y, Iwahana H (1983) Activation of prophenoloxidase by bacterial cell walls or beta- 1,3-glucans in plasma of the silkworm, Bombyx mori. Biochem Biophys Res Commun 113(2): 562-568.
-
Leonard C, Söderhäll K, Ratcliffe, NA (1985) Studies on prophenoloxidase and protease activity of Blaberus craniifer hemocytes. Insect Biochem 15(6): 803-810.
-
Hellio C, Bado-Nilles A, Gagnaire B, Renault T, Thomas- Guyon H (2007) Demonstration of a true phenoloxidase activity and activation of a proPO cascade in Pacific oyster, Crassostrea gigas (Thunberg) in vitro. Fish Shellfish immunol 22(4): 433-440.
-
Ochiai M, Ashida M (1988) Purification of a β-1,3-Glucan recognition protein in the prophenoloxidase activating system from hemolymph of the silkworm, Bombyx mori. J Biol Chem 263(24): 12056-12062.
-
Vargas-Albores F, Plascencia GY (2000) Beta glucan binding protein and its role in shrimp immune response. Aquaculture 191(1-3): 13-21.
-
Perazzolo LM, Barracco MA (1997) The prophenoloxidase activating system of the shrimp Penaeus paulensis and associated factors. Dev Comp Immunol 21(5): 385-395.
-
Aspán A, Söderhäll K (1991) Purification of prophenoloxidase from crayfish cells, and its activation by an endogenous serine proteinase. Insect Biochem 21(4): 363-373.
-
Aspán A, Huang TS, Cerenius L, Söderhäll K (1995) cDNA cloning of prophenoloxidase from the freshwater crayfish Pacifastacus leniusculus and its activation. Proc Natl Acad Sci USA 92(4): 939-943.
-
Nellaiappan K, Sugumaran M (1996) On the presence of prophenoloxidase in the hemolymph of the horseshoe crab, Limulus. Comp Biochem Physiol B 113(1): 163-168.
-
Perdomo-Morales R, Montero-Alejo V, Perera E, Pardo- Ruiz Z, Alonso-Jiménez E (2007) Phenoloxidase activity in the hemolymph of the spiny lobster Panulirus argus. Fish Shellfish Immunol 23(6): 1187-1195.
-
Mahmoud DM, Salem DAM, Mómen SA, Barakat EMS, Salama MS (2015) Purification and characterization of phenoloxidase from immunized haemolymph of Schistocerca gregaria. African J Biotechnol 14(44): 3027-3036.
-
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal Biochem 72: 248-254.
-
Parrinello N, Arizza V, Chinnici C, Parrinello D, Cammarrata M (2003) Phenoloxidases in ascidian hemocytes: characterization of the prophenoloxidase activating system. Comp Biochem Physiol B 135(4): 583- 591.
-
Söderhäll K, Cerenius L (1992) Crustacean immunity. Annu Rev Fish Dis 2: 3-23.
-
Laskin JD, Piccinini LA (1986) Tyrosinase isozyme heterogeneity in differentiating B16/C3 melanoma. J Biol Chem 261(35): 16626-16635.
-
Wright J, McCaskill-Clark W, Cain JA, Patterson A, Coates CJ, et al. (2012) Effects of known phenoloxidase inhibitors on hemocyanin-derived phenoloxidase from Limulus polyphemus. Comp Biochem Physiol B 163(3-4): 303-308.
-
Kim SG, Jung BW, Kim H (2011) Hemocyanin derived phenoloxidase activity with broad temperature stability extending into the cold environment in hemocytes of the hair crab Erimacrus isenbeckii. Comp Biochem Physiol B 159(2): 103-108.
-
Decker H, Jaenicke E (2004) Recent findings on phenoloxidase activity and antimicrobial activity of hemocyanins. Dev Comp immunol 28(7-8): 673-687.
-
Decker H, Tuczek E (2000) Tyrosinase/catecholase activity of hemocyanins: structural basis and molecular mechanism. Trends Biochem Sci 25(8): 392-397.
-
Jaenicke E, Decker H (2004) Conversion of crustacean hemocyanin to catecholoxidase. Micron 35(1-2): 89-90.
-
Baird S, Kelly SM, Price NC, Jaenicke E, Meesters C, et al. (2007) Hemocyanin conformational changes associated with SDS-induced phenol oxidase activation. Biochim Biophys Acta 1774(11): 1380-1394.
-
Lee SY, Lee BL, Söderhäll K (2004) Processing of crayfish hemocyanin subunits into phenoloxidase. Biochem Biophys Res Commun 322(2): 490-496.
-
Smith V J, Söderhäll K (1983) β-1,3-glucan activation of crustacean hemocytes in vitro and in vivo. Biol Bull 164: 299-314.
-
Söderhäll K (1992) Biochemical and molecular aspects of cellular communication in arthropods. Boll Zool 59(2): 141-151.
-
Duvic B, Söderhäll K (1990) Purification and characterization of a β-1,3-glucan binding protein from the plasma of the crayfish Pacifastacus leniusculus. J Biol Chem 265(16): 9327-9332.
-
Vargas-Albores F, Vega FJ, Söderhäll K (1996) A plasma protein isolated from brown shrimp (Penaeus californiensis) which enhances the activation of prophenoloxidase system by β-1,3-glucan. Dev Comp Immunol 20(5): 299-306.
-
Vargas-Albores F, Jiménez-Vega F, Yepiz-Plascencia G (1997) Purification and comparison of β-1,3-glucan binding protein from white shrimp Penaeus vannamei. Comp Biochem Physiol B 116(4): 453-458.
-
Liu G, Yang L, Fan T, Cong R, Tang Z, et al. (2006) Purification and characterization of phenoloxidase from crab Charybdis japonica. Fish Shellfish Immunol 20(1): 47-57.
-
Feng CJ, Song QS, Lü WJ, Lu JF (2008) Purification and characterization of hemolymph prophenoloxidase from Ostrinia furnacalis (Lepidoptera: Pyralidae) larvae. Comp Biochem Physiol B 151(2): 139-146.
-
Zibaee A, Bandani AR, Malagoli D (2011) Purification and characterization of phenoloxidase from the hemocytes of Eurygaster integriceps (Hemiptera: Scutelleridae). Comp Biochem Physiol B 158(1): 117-123.
-
Huang J, Yang Y, Wang A (2010) Reconsideration of phenoloxidase activity determination in white shrimp Litopenaeus vannamei. Fish Shellfish Immunol 28(1): 240-244.
-
Sung HH, Chang HJ, Her CH, Chang JC, Song YL (1998) Phenoloxidase activity of hemocytes derived from Penaeus monodon and Macrobrachium rosenbergii. J Invertebr Pathol 71(1): 26-33.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind