Nutraceutical Properties of Potentially Pollen Sources in Riyadh, Saudi Arabia and their Impact on the Activities of Honey Bee Colonies
Pollen plants is the sole source of strength for honey bees’ colonies, estimating pollen nutritional value is a crucial aspect to comprehend its influence on the growth of honey bee colonies in any region. In this study the potentially discovered pollen source in Riyadh region, central Saudi Arabia, Asian mustard (Brassica tournefortii Gouan.), willow wattle (Acacia salicina Lindl.), mesquite (Prosopis juliflora (Sw.) DC.) and cat’s head (Tribulus terresetris L. var. terrestris) were evaluated and their role in improving honey bee colonies were analyzed. Important criteria for determining the nutraceutical properties of pollen included fresh and dry weight, protein content, and the presence of essential and non-essential amino acids were analyzed. The study revealed that the highest content of protein in the dominant pollen types was recorded for B. tournefortii pollen (20.53%), while P. juliflora pollen was the highest in essential amino acids content. In conclusion, in an effort to achieve highquality development, careful plant breeding and rehabilitation of these potentially discovered pollen plant species would be taken into consideration. In conclusion, wise use and rehabilitation of these potentially identified bee plant species shall be considered in attempting beekeeping development.
Introduction
By foraging on flowering plants, honeybees can obtain all of their nutritional requirements via collecting nectar and pollen grains (i.e. protein, carbohydrates, amino acids, vitamins and minerals) [1, 2]. Pollen grains play a vital key role in honeybee nutrition and honeybees collect pollen grains from plants to obtain protein which is necessary for their survival and colony strength [3, 4, 5], as well as, pollination [6], and their productivity [7]. The botanical composition of natural vegetation varies depending on the topography, climate and soil type. However, the potential for different colony products and success in beekeeping development is dependent mainly on the type and quantity of the available flora [8]. Generally, the diversity of pollen depends on plant habitat, environmental conditions, distributions, and flowering season [9]. However, not all flowering plants offer the same amounts or blends of nutrients [10]. Depending on plant species, studies conducted by Roulston, et al. [11] and Begum, et al. [12] suggested that the protein content of pollen varies significantly, from around
2.5% dry weight in Solanum sp. (Solanaceae) to 62% in Dodecatheon clevelandii (Primulaceae). Thus, the variation and diversity of the available foraging flora to honeybee is dependent not only on the local landscape composition, but also on the nutritional content provided by the pollen and nectar of these plants [13].
Many researchers focused on palynological studies to help beekeepers in the abundance, distribution and flower blooming calendars of the main pollen sources in a selected study area [8, 14, 15]. Beekeepers must have information about the flowering periods of both major and minor nectar and pollen producing plants in the vicinity of their apiaries for successful honey production to enable them to determine the suitable time to introduce various management practices to their colonies [16]. Bees should be given a range of diverse floral resources from which they can self-select their diet to meet their component nutrient requirements, which will sustain healthy populations that can endure diseases and stresses. Among the feeding resources, polyfloral pollen diets can provide a balance of essential amino acids and fatty acids, whose concentrations differ between plant species [17].
In the central region of Saudi Arabia, long dry periods and recurrent droughts are very common. These conditions have led to a decline in the number of flowering plants, resulting in a shortage of pollen available and nectar sources. Thus, a high percentage of imported honeybees and a significant number of local honeybee colonies are dying annually [18]. Hence, this study aims to investigate the nutraceutical properties of pollen plant species that may have been discovered in Riyadh, Middle Arabia, and assess their effect on the activities of honeybee colonies, as well as, the adequacy of each plant species as a pollen source for honeybee nutritional needs.
Material and Methods
The experiments were conducted in the laboratories and apiary of Bee Research Unit, College of Food and Agriculture Sciences, King Saud University, Riyadh region, Saudi Arabia. Ten colonies, equal in strength of the native honeybee race (Apis mellifera jeminitica) and imported Carniolan hybrid (Apis mellifera carnica) headed by sister queens, were located in Derab farm (24°24’1.34”N 56°39’36.21”E) in Riyadh during 2016 from January to December.
Determination of Seasonal Fluctuation of Bee Bread Areas and Some Activities of Honeybee Colonies
The present work was carried out through the period from January to December in 2016 to find the time of pollen shortage. Two methods were monitored to realize: the first was the measurement of bee bread, sealed brood area and bee population of honeybee colonies. The second was the determination of protein percentage variations of stored pollen in honeybee colonies.
Measurement of Bee Bread, Sealed Brood Areas and Bee Populations
Areas of bee bread and sealed brood were determined around the year at 12 days intervals by using graduated grid divided into square inches after the honeybee had been shaken from the combs [19]. Ten honeybee colonies were used, five colonies for Yemeni race and another for Carniolan hybrid bees. Number of combs covered with bees from both sides was recorded in all colonies under investigation. Colony bee population was estimated in the rate of 2000 adult bees for every comb covered with bees from both sides [16].
Determination of Bee Bread Quality
Samples Collection: To determine bee bread quality, mixed samples were collected monthly from the combs near brood nest during the experimental period from five colonies (5 gram/colony) of Yemeni and Carniolan honeybee colonies. The samples were kept at -10 °C until the time of chemical analysis.
Total Protein Determination: To measure the total protein content 0.7 g of the dry samples from bee-pollen and/or bee bread were digested in a macro Kjeldahl flask containing 4 g of the catalytic mixture (1:3 CuSO4 and K2SO4) and 20 ml of concentrated H2SO4. After digestion, 80 ml of 40% NaOH was added to release ammonia, which was collected in an H2SO4 solution, and then, titrated with a standardized NaOH, 0.1 M solution. Calculate the total protein, the nitrogen values were multiplied by conversion factor 5.60 using a Vapodest 45 (Gerhardt) according to Rabie, et al. [20].
Determination of Total Amino Acids (Hcl-Hydrolyzed): Amino acids content for pollen grains was determined as described by Spackman, et al. [21]. The analysis was performed in Central Service Unit, National Research Center, Egypt, A qualitative and quantitative analysis of the amino acids was performed by ion exchange chromatography with the use of an Automatic Amino Acid Analyser LC3000 (Eppendorf-Biotronik, Germany). The technique was based on the separation of the amino acids using strong cation exchange chromatography followed by the ninhydrine colour reaction and photometric detection at 570 nm. 0.1g dry matter of every pollen type was hdyrolysed with 10 ml 6N HCl in a sealed tube at 110ºC in teflon capped vials for 24 hr. [22]. The pollen samples were treated with performic acid in order to avoid the decomposition of sulphur-containing amino acids. The hydrolysis tube was then closed by melting the glass with a suitable gas burner and hydrolyzed in an oven with a uniform temperature distribution of 110°C for 24 hr (in order to create well reproducible hydrolysis conditions. After hydrolysis, the solution was evaporated in a vacuum evaporator (EYELA Rotary Vacuum Evaporator) under reduced pressure at 40ºC and then dissolved with approximately 1 ml distilled water and evaporated once again in order to remove traces of acid. The dry residue was transferred to a 50 ml measuring flask. The obtained solution was filtered through a 0.45 μm membrane filter and 100 μl of filtrate was injected into the apparatus. After vacuum removal of HCl, the residues were dissolved in a lithium citrate buffer, pH 2.2. Twenty µl of the solution were loaded onto the cation exchange column (pre-equilibrated with the same buffer), then four lithium citrate buffers with pH values of 2.2, 2.8, 3.3 and 3.7, respectively, were successively applied to the column at flow rate 0.2 ml/min. The ninhydrine flow rate was 0.2 ml/min and pressure of reagent 0-150 bar. The pressure of buffer was from 0 to 50 bar and reaction temperature was 130ºC. Amino acids in pollen were identified by comparing the retention time of particular amino acids in the reference solution and in an examined solution (qualitative analysis); the quantitative analysis were performed by comparing the peak area which corresponded to those amino acids. The standard solution containing a mixture of amino acids, with each one present at a concentration of 2.5 μM/ml as a reference solution. The data were converted into dry matter. Essential amino acid content was also expressed as a percentage of the total concentration of all amino acids in the tested samples.
Statistical Analysis: The experiment was set up according to a complete randomized design. For the aforementioned parameters, the obtained data were statistically analyzed using analysis of variance (ANOVA). The means were compared using Duncan’s multiple range test at a significance level of 5% with the SAS 9.1.3 programme [23].
Results
Seasonal Fluctuations of Bee Bread Areas and Some Activities in Honeybee Colonies Under Riyadh Conditions
Bee Bread Areas and its Protein Content: Monthly means are graphed in (Figure 1). The results showed that the area of bee bread fluctuated during active season. Bee bread stored in January was small and started to increase in February. Then, it decreased during March. In April bee bread areas increased and the second peak was recorded. The third and largest peak of stored pollen area was recorded in August, the period of some main plants flowering. The area of bee bread started to decrease during September.
In a comparison between the local race and imported hybrid, ability for storing bee bread, the local honeybee race ranked the first. Clear variations between the two honeybee races were recorded. The highest variance was recorded in July (79.40) while the lowest variance was recorded in October (-1.25). Results indicated that local race stored 89.90, while Carniolan one stored 70.21 (sq.inch/colony) with highly significant differences as a general monthly mean for the study period. The results revealed shortage of pollen area stored during March followed by another one in May-June period under study area conditions. This drop of records indicated that the pollen collection stoped and more consumption from bee bread was used in feeding the reared brood and newly emerged bees.
Our data showed that crude protein percentages samples of bee bread contained more than 19% for stored pollen in February, April and July. However, in the autumn period (September – October) the crude protein percentage decreased and ranged between 14.03-16.37%. The highest protein percentage was recorded in August while the lowest one was recorded in November. It is clear that the protein content of bee bread greatly depending on plant species.
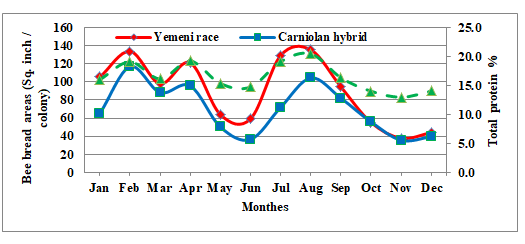
Brood Rearing Activity: The brood areas measured and means of monthly results are presented in Table 1. The obtained results for all tested bees under environmental conditions of Riyadh indicated a similar trend for seasonal fluctuation of sealed brood area. It started with small records in January and started to increase in February and reached to the first peak in March, then decreased relatively during April, the first period of pollen scarcity in the area of the study. Brood rearing activity started to increase again at May associated with second peak of pollen collection. The highest third peak of brood rearing was recorded during August and then the area of sealed brood decreased gradually in September and reached to the minimal level on December. Furthermore, sealed brood areas in Yemeni colonies showed high areas during the experimental period than those in Carniolan hybrid colonies. The area of sealed brood in Yemeni race averaged 177.52 sq.inch/colony is equal 1.20 fold of that (147.64 sq.inch/colony) constructed by the Carniolan hybrid. Colony Population: Population of honeybee workers In respect to bee populations and sealed brood areas proportions, clear variations were detected between Yemeni and Carniolan races all over the study period. Concerning the bee populations the highest variable was recoded during July (31.16%), while the lowest variable was recorded during May (0.02%). Concerning the sealed brood areas the highest variable was recorded during November (36.92%), while the lowest variable was recorded during October (3.43%).
measured during season of 2016. Data were listed in Table 1. The results indicated that there was a similar trend for seasonal fluctuations of bees number for both race and hybrid honeybees. It was closely related with the variation of sealed brood area. Data show that bees number started to increase from the beginning of February and reached to the first peak on March, then decreased until May. The bees number started to increase again to reach a second peak of population in June. The maximum numbers of honeybee workers were recorded during September – October period. The honeybee colonies started to decrease in November.
| Months | Means (±S.D. ) of Bee Population** | Means (±S.D.) of Sealed Brood (Sq.Inch/Colony) | ||||
|---|---|---|---|---|---|---|
| Carniolan Hybrid | Yemeni Race | Variation % | Carniolan Hybrid | Yemeni Race | Variation % | |
| Jan. | 6833.5 ± 297.76 n* | 7515.0 ± 577.74 kl | 9.97 | 111.00 ± 10.06 i | 133.50 ± 14.26 h | 20.27 |
| Feb. | 7229.3 ± 287.53 lmn | 9268.7 ± 386.15 hi | 28.21 | 152.13 ± 12.30 fgh | 177.13 ± 17.55 cde | 16.43 |
| Mar. | 8843.0 ± 345.25 ij | 10450.6 ± 421.8 le | 18.18 | 182.85 ± 26.76 bcd | 203.55 ± 22.12 b | 11.32 |
| Apr. | 8576.1 ± 337.07 j | 9861.7 ± 338.82 fg | 14.99 | 156.76 ± 16.77 efg | 164.63 ± 18.29 defg | 5.02 |
| May | 7816.0 ± 440.67 k | 7817.5 ± 446.68 k | 0.02 | 171.35 ± 32.86 def | 197.55 ± 35.55 bc | 15.29 |
| Jun. | 8545.0 ± 313.58 j | 9607.0 ± 405.79 gh | 12.43 | 142.60 ± 19.89 gh | 164.80 ± 16.97 defg | 15.57 |
| Jul. | 6900.0 ± 277.11 mn | 9050.0 ± 335.02 i | 31.16 | 150.20 ± 13.06 fgh | 202.85 ± 41.60 b | 35.05 |
| Aug. | 9200.0 ± 361.79 hi | 11166.7 ± 274.3 ld | 21.38 | 175.56 ± 16.62 de | 240.36 ± 19.40 a | 36.91 |
| Sept. | 12544.1 ± 319.96 b | 12543.1 ± 419.96 b | -0.01 | 182.78 ± 24.37 bcd | 225.63 ± 28.00 a | 23.44 |
| Oct. | 12644.8 ± 243.51 b | 13509.5 ± 424.60 a | 6.84 | 160.55 ± 25.37 defg | 166.05 ± 32.21 defg | 3.43 |
| Nov. | 10072.5 ± 355.04 ef | 11937.5 ± 362.86 c | 18.52 | 104.70 ± 10.75 hg | 143.36 ± 9.58 i | 36.92 |
| Dec. | 7341.8 ± 361.95 lm | 9001.0 ± 568.90 ij | 22.6 | 81.25 ± 4.05 i | 110.85 ± 10.14 j | 36.43 |
| General mean | 8878.75 ± 1966.43 B | 10232.51 ± 1972.41 A | 15.25 | 147.64 ± 29.75 B | 177.52 ± 50.93 A | 20.24 |
Table 1: Fluctuation of colony population and sealed brood areas (sq.inch/colony) in Yemeni races and Carniolan hybrid honeybe
Source: Means followed by the same letter are not significantly different at 0.05 level of probability, by Duncan’s multiple range test. S.D.= Standard deviation. Table 1: Fluctuation of colony population and sealed brood areas (sq.inch/colony) in Yemeni races and Carniolan hybrid honeybee colonies under Riyadh conditions, during 2016.
Nutraceutical Properties of Pollen Sources
Crude Protein, Fresh and Dry Weight of Pollen Pellets: Data presented in Table 2 showed the crude protein and weight of fresh pollen pellets of the main pollen sources collected by the two types of honeybee in Riyadh. It is important to note that, the fresh weight of pollen pellets collected by Carniolan hybrid honeybee recorded more than 900 mg/100 pellets during their foraging on the aforementioned plant species. The highest fresh weight in the collection was obtained by the Carniolan hybrid honeybee during its foraging on salad rocket (E. sativa). An average of 999.46 mg/100 pellets was recorded. However, the lowest fresh weight was recorded when Yemeni honeybee broad on cat’s head (T. terrestris) with an average of 619.02 mg/100 pellets. Significant differences were recorded between the pellets collected by Carniolan hybrid and Yemeni honeybee race (P< 0.05). The obtained data revealed that clear variations between total protein content values in the four dominant pollen sources. The data showed that, the highest concentration of the total protein content was found in asian mustard (B. turneforti) pollen (20.53 mg/gm), while, the lowest concentration was recorded in the cat’s head (T. terrestris) pollen (17.69 mg/ gm), with highly significant difference (P<0.01). In another view, data clarified that, the total protein contents in asian mustard pollen is equal 1.16, 1.09 and 1.04 fold of those recorded in cat’s head, mesquite (P. juliflora) and willow (A. salicina) pollen, respectively.
| Common Name | Family Name | Scientific Name | Fresh weight (MG/100 Pellets) ± S.D. | Means (±S.D.**) of Crude Protein (%Dray Matter) | |
|---|---|---|---|---|---|
| Yemeni Race | Carniolan Hybrid | ||||
| Date Palm | Arecaceae | Phoenix dactylifera L. | 651.51 ± 0.01 B* | 967.51 ± 0.01 C | 22.31 ± 0.04 A |
| Salad Rocket | Brassicaceae | Eruca sativa miller) Thell. | 649.15 ± 0.01 C | 999.46 ± 0.03 A | 16.05 ± 0.03 DE |
| Asian Mustard | Brassicaceae | Brassica tournefortii gouan. | 673.46 ± 0.01 A | 986.41 ± 0.05 B | 20.53 ± 0.05 AB |
| Willow Wattle | Fabaceae | Acacia salicina lindl. | 651.98 ± 0.02 B | 996.55 ± 0.01 A | 19.71 ± 0.07 B |
| White Lead Tree | Fabaceae | Leuceana leucocephala (Lam.) de wit | 620.88 ± 0.01 D | 982.72 ± 0.01 Ab | 16.00 ± 0.07 DE |
| Parkinsonia | Fabaceae | Parkinsonia leculeate L. | 650.52 ± 0.01 Bc | 946.02 ± 0.03 D | 15.45 ± 0.04 E |
| Mesquite | Fabaceae | Prosopis juliflora (Sw.) Dc. | 626.14 ± 0.02 C | 908.06 ± 0.01 E | 18.67 ± 0.07 C |
| Camphor | Myrtaceae | Eucalyptus camaldulensis dehnh. | 637.15 ± 0.01 Cd | 965.46 ± 0.01 C | 15.24 ± 0.02 E |
| Cat’s Head | Zygophyllaceae | Tribulus terrestris L. var. terrestris | 619.02 ± 0.01 E | 980.37 ± 0.01 Ab | 17.69 ± 0.02 D |
Table 2: Weight of fresh pollen pellets of the main pollen sources collected by Yemeni race and Carniolan hybrid honeybee colo
Source: Means followed by the same letter in the same column are not significantly different at 0.05 level of probability, by Duncan’s multiple range test. S.D.= Standard deviation. Table 2: Weight of fresh pollen pellets of the main pollen sources collected by Yemeni race and Carniolan hybrid honeybee colonies in Riyadh during 2016. Essential and Non-Essential Amino Acids: Eighteen amino acids were identified in the main four dominant pollen source. It is clear that all essential amino acids i.e. arginine, histidine, isoleucine, leucine, lysine, methionine, phenylalanine, threonine, tryptophan and valine, were found in the samples collected from the examined pollen sources. However, 8 non-essential amino acids contained alanine, aspartic acid, cysteine, glutamic acid, glycine, proline, serine and ammonia, were identified.
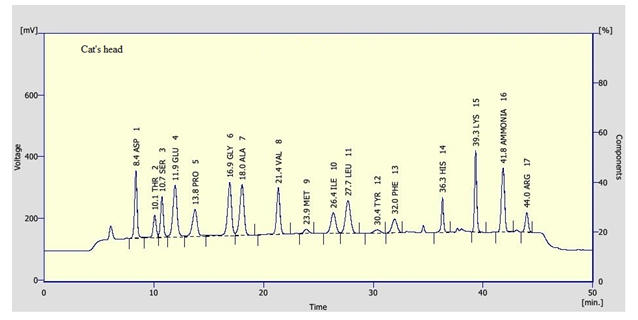
Clear variations between the concentrations of the essential amino acids were recorded between the tested pollen types. Leucine and Lysine were found in relatively high concentration in the tested samples. Leucine was recorded as the highest constituent between the tested essential amino acids in willow wattle, mesquite and cat’s head pollen pellets with an average of 11.57, 17.51 and 8.36 mg/g, respectively. However, Lysine was recorded as the highest concentration in mesquite (Figure 3) pollen pellets with an average of 16.80 mg/g. Leucine and Lysine together were accounted for about 14.42 and 34.31 mg/g DM, respectively of total amino acid content in tested pollen sample. In general, data revealed that cat’s head (Figure 5) pollen pellets contained the highest essential amino acid concentrations rather than the remaining pollen sources. However, willow wattle (Figure 2) pollen pellets contained the least essential amino acid concentrations. The concentration of methionine was relatively low. Its content ranged from 1.07 to 2.14 mg/g DM. The highest concentration of methionine was found in mesquite pollen and the lowest one was recorded in cat’s head pollen.
In respect to the non-essential amino acids, values were recorded in the tested pollen types. Prolin and Cysteine were recorded as the highest and the lowest concentration among the non-essential amino acids, respectively. Prolin values were 21.32, 26.45 and 28.15 mg/g, in willow, mesquite and cat’s head pollen pellets, respectively. However, Cysteine value was the lowest concentration in willow pollen pellets (0.83 mg/g) and mesquite pollen (1.27 mg/g). Also, it completely absent from asian mustard (Figure 4) and cat’s head pollen.
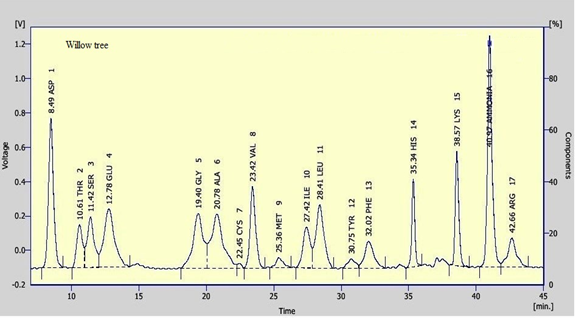
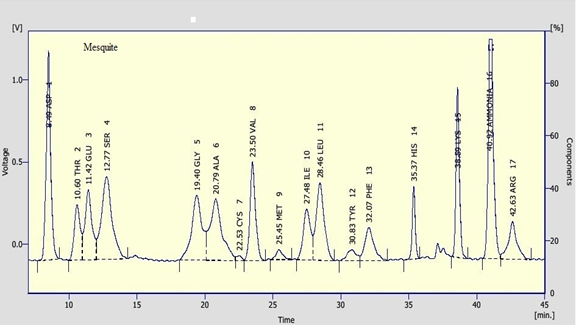
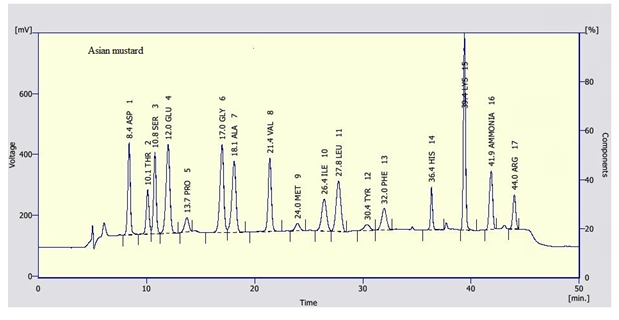
Seasonal Fluctuations of Bee Bread and Some Activities of Honeybee Colonies Under Riyadh Environmental Conditions
Bee Bread Areas and its Protein Content: The largest stored bee bread areas initiated by Yemeni race and Carniolan hybrid honeybee were recorded during summer months when honeybees foraged on white lead tree (L. leucocephala), mesquite (P. juliflora) and cat’s head (T. terrestris), followed by spring season during the flowering periods of date palm (Phoenix dactylifera L.), asian mustard (B. tournefortii) and willow wattle (A. salicina). The current results showed that rape, date palm and the Acacia trees are the main pollen sources. That were in agreement with the findings of Taha, et al. [24] whose stated that the main pollen sources of Alhasa region in Saudi Arabia were rape, date palm and the Acacia trees.
The amount of bee bread in the colonies depends on the amount of collected pollen, as well as, the rate of pollen consumption by nurse bees [25]. In the present study, Yemeni honeybees stored pollen 1.3 fold more than Carniolan hybrid honeybees. This increase in pollen storage may be due to the high numbers of pollen foragers resulted from the high bee population in Yemeni bees as compared to Carniolan hybrid bees. These results are supported by the findings of Al-Qarni [26]; Al-Ghamdi [18]; Ali [27], Taha, et al. [24] and Taha [28]. They reported that pollen collected by Yemeni bee colonies was higher than that collected by the Carniolan bee colonies. Meanwhile, Yemeni bees appeared vigor in brood rearing activity rather than Carniolan bees. This finding, may be considered as the main reason for superiority of Yemeni honeybee colonies in pollen storage.
It is important to note that the highest stored bee bread areas were coincided with the highest protein percentage The present results exhibited clear difference between pollen and bee bread protein content. In this approach, Haydak [30] and Amro, et al. [31] attributed differences between pollen and bee bread protein content to the addition of other matters such as honey and honeybee secretions, and the activity of microorganisms in the process of transformation of pollen into bee bread. Also, this might be a result of adding nectar sugars during formation of pellets, which caused increase of the content of carbohydrates, while the protein, lipid and ash contents decreased in bee bread as described by Nicolson, et al. [32]
Bee Population and Sealed Brood Areas: The fluctuation of bee population in local honeybee colonies were higher than those in Carniolan hybrid one. These results were in agreement with Al-Qarni [33] and Al-Qarni, et al. [34] that highly significant difference between Yemeni and Carniolan colonies population. On the other hand, the highest numbers of bees on combs were recorded during autumn (mesquites pollen flow) as a result of the high rate of stored pollen and worker sealed brood during summer. Another peak of bee population has been shown in March in the same year but smaller than the first one. This result reflects the highly nutritional effect of asian mustard pollen (B. tournefortii) for honeybee to build up their colonies. Also, the lowest bee population density was recorded during winter in both tested races. These results agree with the results of Taha, et al. [24]
who found that the major peak of brood activity and higher rate of stored pollen were recorded in May, consequently the maximum number of house bees was recorded during June and July. The decrease in population size of colony resulted from low rate of brood rearing due to the sharp decrease in pollen collection as a result of dearth of most pollen flora or the available pollen source in the late autumn. Moreover, Parkinsonia and salad rocket pollen which naturally available in this period, don’t met the nutritional requirement of honeybees to build up their colonies.
All over the study period, the area of worker sealed brood in Yemeni honeybee colonies was larger than that of Carniolan hybrid. Finding of Ali [27] and Al-Qarni, et al. [34] confirmed the present results. They reported that the highest monthly sealed brood areas recorded with native bees were 1 fold higher than Carniolan one. The highest brood rearing activities peak presented in August which synchronized with presence of mesquite pollen in the study area and the second peak was synchronized with presence of asian mustard pollen in February. However, the third sealed brood peak was coincided with presence of willow wattle pollen flow in early May. Hence, resulting in three peaks of population size, highest one was in October, another in March and third in June for both tested races. These results are considered as evidence for the goodness of mesquite, asian mustard and willow wattle pollen and reflect their quality for brood rearing activity and colony population size. Also, high ability and good adaptation to the harsh environmental conditions by indigenous race was recorded by Ali [27]. Meanwhile the Carniolan race is not acclimatized by the hot summer. Differences between variable races were discovered by Camazine [35] and Pankiw, et al. [36]. They found that the genotype strongly affected the foraging behavior.
Significant (P<0.01) positive correlations were found between worker sealed brood area and each of stored pollen area and colony population (r = 0.618, 0.228, respectively). These results agreed with the findings of Schneider and McNally, et al. [37] and Mladenovic, et al. [38] who are, reported that there is a positive correlation between the increase pollen gathering activity and increase of brood rearing. In addition, Jevtic, et al. [39] proved that there is a positive correlation between the colony population, brood area and honey yield.
Chemical Characteristics of Pollen Grains
The obtained results revealed that the fresh weight of pollen grains collected by Carniolan hybrid honeybee was higher than those collected by Yemeni honeybee race. Unfortunately, few information is available about the fresh weight of pollen pellets collected by diverse honeybee races from different plant sources. However, Radev [40] concluded that the water content in the analyzed samples from the selected botanical origins ranged between 9.7% (Centaurea calcitrapa) to 23.1% (Crepis sp.) with the average of 17.4%. The water content in the mixed pollen for different periods ranged between 12.3% to 26.6% and the average was 18.8%. The water content in the pollen is affected mostly by the humidity of the environment. The determination of water in the pollen samples was necessary to report the protein content. Protein Contents: Data published about Saudi Arabian monofloral pollen sources are few. The present study compared four main pollen sources in Riyadh (willow wattle, mesquite, asian mustard and cat’s head) pollen in chemical composition. Protein and amino acids contents recorded at selected locations and relation to some determined components. Among thirteen pollen types utilized by honeybees in the study area, only four sources were considered monoflora. These samples were exclusively composed of B. tournefortii (Brassicaceae), A. salicina and P juliflora (Fabaceae) and T. terrestris (Zygophyllaceae) pollens. Highly significant differences between pollen protein content percentages for different plant sources were recorded. Superiority of crude protein percentage was recorded for asian mustard (20.53%) pollen followed by willow wattle (19.71%), mesquite pollen (18.67%) and cat’s head pollen (17.69%).
Studies conducted by Roulston, et al. [11] and Weiner, et al. [17] reported that bees taking the majority of their protein from pollen, but protein concentration varies considerably between plant species and ranged from 2 to 60%. The average levels of protein found in the current study exhibited low range and in agreement with Szczêsna [41] results. However, the average values were lower than the values found in Polish pollen as reported by Rogala, et al. [42]. In respect to the collected pollen from Polish, South Korean and Chinese bees, Szczêsna [43] found that protein values ranged from 15.80 to 24.14 g/100g, 17.63 to 24.51 g/100g and 17.83 to 26.13 g/100g, respectively. The great variability of protein contents found in bee pollen can be partly explained by the natural compositional variation. Such variation is influenced by floral origin, biological, ecological, and geographic factors during production as well as handling and storage conditions [44].
In the same line, finding of Radev [40] confirmed that during spring, pollen with protein content above 21%, and especially above 27% allows colonies to maintain a high level of reproduction and development. Moreover, honeybees collected pollen widely different in protein content; but a protein percentage 20% or more is enough to satisfy the nutritional requirements of honeybees [45]. Also, preference for high protein pollen has not been clearly demonstrated for honeybees [46].
The current results concluded that, bee pollen collected from B. tourneforii (Brassicaceae) and A. salicina have the highest protein content, followed by P. juliflora (Fabaceae). However, the lowest protein content was detected in bee pollen of T. terrestris. These results were confirmed with findings of Kosonocka [47] in USA, Pernal, et al. [48] in Canada, Tasei, et al. [49] in France and Taha [29] in Saudi Arabia. They also found high protein content in Brassica spp. pollen collected by bees and reached to 21.7%, especially on mustard. Moreover, Dastagir, et al. [50] indicated that T. terrestris pollen had the maximum protein and gross energy among another tested plants in their study. The results obtained by Almaraz Abarca, et al. [51] suggested that pollen of P. juliflora is an important source of protein. On the other hand, the earlier Agarwal, et al. [52] reported that the total protein content of willow wattle pollen recorded 19.71%.
Differences between protein contents recorded in this work in Saudi Arabia and the values found in other countries might be correlated to the plant growth conditions, such as: soil, irrigation water, and geographic origin. For example; the electrical conductivity of groundwater used for irrigation in semi-arid region ranged between 1.23 and 5.05 dS m-1 with Na as the most abundant cation. Likewise, the groundwater of study region is mainly Na and Ca water rather than Ca and Mg dominant water [53]. Amino Acid Contents: Clear differences of amino acid content between different pollen type were recorded. In the same line concluded that the concentration of amino acids content was dependent on the floral origin of pollen [41, 54]. In present results tryptophan and phenylalanine were recorded as the lowest essential amino acid contents in all tested pollens in addition to full absence of cysteine as non- essential amino acid in cat’s head plant pollen. This finding is congruence with Lundén [55] results who recorded that tryptophan and phenylalanine were absent in most pollen types. Moreover, mustard is a superior pollen source of bee nutrition because of amino acid rich contents in asian mustard pollen (Brassicaceae) [56].
Concerning to the aspartic acid, glutamic acid and glycine as non-essential amino acids in willow wattle, the current results were disagreed with Agarwal, et al. [52]. They reported that protein bound amino acids were found to be rich in aspartic acid, glutamic acid and glycine. This difference may be due to the lack of amino acid in other tested plants, or affecting of different geographical regions.
Low concentrations of the essential amino acids in bee pollen of cat’s head plant were currently recorded, especially histidine. Also, the tested honeybees showed the same trend in the period of cat’s head pollen flow. So, polyfloral pollen diets can provide a balance of essential amino acids and fatty acids, whose concentrations differ between species [17].
Authors Contributions
All authors of this manuscript contributed equally to the design and/or execution of the experiments described in the manuscript. AMA prepared and edited the final version of this manuscript. All authors approved the final manuscript.
Conflicts of Interest
The authors declare that there are no conflicts of interest associated with this publication.
References
-
Morgano MA, Martins MCT, Rabonato LC, Milani RF, Yotsuyanagi K, et al. (2012) A comprehensive investigation of the mineral composition of Brazilian bee pollen: geographic and seasonal variations and contribution to human diet. J Braz Chem Soc 23(4): 727- 736.
-
Donkersley P, Rhodes G, Pickup RW, Jones KC, Wilson K (2014) Honeybee nutrition is linked to landscape composition. Ecol Evol 4(21): 4195-4206.
-
Barth OM, Munhoz MC, Luz CFP (2009) Botanical origin of Apis pollen loads using colour, weight and pollen morphology data. Acta Alimentaria 38(1): 133-139.
-
Belay A, Solomon WK, Bultossa G, Adgaba N, Melaku S (2015) Botanical origin, colour, granulation, and sensory properties of the Harenna forest honey, Bale, Ethiopia. Food chem 167: 213-219.
-
Amro A, Omar M, AL Ghamdi A (2016) Influence of different proteinaceous diets on consumption, brood rearing, and honey bee quality parameters under isolation conditions. Turkish Journal of Veterinary and Animal Sciences 40(4): 468-475.
-
Amro AM (2021) Pollinators and pollination effects on three canola (_Brassica napus_ L.) cultivars: A case study in Upper Egypt. Journal of King Saud University-Science 33(1): 101240.
-
Ghosh S, Jung C (2017) Nutritional value of bee-collected pollens of hardy kiwi, Actinidia arguta (Actinidiaceae) and oak, Quercus sp. (Fagaceae). Journal of Asia-Pacific Entomology 20(1): 245-251.
-
Amro A, Omar M, AL Ghamdi A (2023) Surveying, identification and characterization for the potential honeybee (_Apis mellifera_ L.) pollen sources in the arid region of Riyadh-Saudi Arabia. Bee Studies 15(1): 1-11.
-
Amro A, Omar M, AL Ghamdi A (2020) Impact of using pollen substitutes on performance of honey bee (_Apis_ _mellifera_ l.) colonies under harsh environmental conditions. Science and Animal Health 8(3): 236-256.
-
Amede T (2003) Natural resources degradation and environmental concerns in the Amhara national regional state: impact on food security. In: Proceedings of the National Resource Management Conference, Bahir Dar, Ethiopia, pp: 24-26.
-
Roulston TH, Cane JH (2000) Pollen nutritional content and digestibility for animals. Plant systematics and Evolution 222: 187-209.
-
Begum HA, Iqbal J, Aziz A (2021) Characterization of pollen profile of Apis mellifera L. in arid region of Pakistan. Saudi journal of biological sciences 28(5): 2964-2974.
-
Albarrak A, Gray A (2023) An overview of beekeeping in the Kingdom of Saudi Arabia. Journal of Apicultural Research 62(1): 11-22.
-
Wubie AJ, Bezabeh A, Kebede K (2014) Floral phenology and pollen potential of honeybee plants in North-East dry land areas of Amhara region, Ethiopia. J Agri Vet Sci 7(5): 36-49.
-
El Sofany A, Naggar YA, Naiem E, Giesy JP, Seif A (2020) Authentication of the botanical and geographic origin of Egyptian honey using pollen analysis methods. Journal of Apicultural Research 59(5): 946-955.
-
Omar EM, Amro AM (2023) Improving pollen substitutes to maintain development and hemolymph parameters of honey bees (_Apis mellifera_ L.) during pollen dearth periods. Journal of Apicultural Research 62(4): 777-786.
-
Weiner C, Hilpert A, Werner M, Linsenmair KE, Blüthgen N (2010) Pollen amino acids and flower specialisation in solitary bees. Apidologie 41: 476-487.
-
Al Ghamdi A (2009) Comprehensive study for the current beekeeping industry of imported and native honeybee in Kingdom of Saudi Arabia. King Abdulaziz City for Science and Technology. Final report of project, pp: 21-39.
-
Jeffree EP (1958) A shaped wire grid for estimating quantities of brood and pollen in combs. Bee World 39(5): 115-118.
-
Rabie AL, Wells JD, Dent LK (1983) The nitrogen content of pollen protein. J Apic Res 22(2): 119-123.
-
Spackman DH, Stein WH, Moore S (1958) Automatic recording apparatus for use in chromatography of amino acid. Annal Chem 30(7): 1190-1206.
-
Bailey JL (1967) Techniques in Protein Chemistry. Elsevier Publishing Co London, UK, pp: 340-351.
-
SAS Institute (2004) The SAS System Version 9.1.3. SAS Institute, Cary, NC.
-
Taha EKA, Al Kahtani SN (2013) Relationship between population size and productivity of honeybee colonies. J Entomol 10(3): 163-169.
-
De Grandi Hoffman G, Eckholm B, Huang M (2013) A comparison of bee bread made by Africanized and European honeybees (_Apis mellifera_) and its effects on hemolymph protein titers. Apidologie 44: 52-63.
-
Al Qarni AS (2006) Influence of some protein diets on the longevity and some physiological conditions of honeybee Apis mellifera L. workers. J Biol Sci 6(4): 734- 737.
-
Ali MAM (2011) Comparative study for evaluating two honeybee races, Apis mellifera jementica (indigenous race) and _Apis mellifera_ carnica (carniolan race) in brood production, population development and foraging activity under the environmental conditions of the central region of the Kingdom of Saudi Arabia. Ann Agric Sci 56(2): 127-134.
-
Taha EKA (2015) A study on nectar and pollen sources for honeybee, _Apis mellifera_ L. in Al Ahsa Saudi Arabia. J Entomol Zool Studies 3(3): 272-277.
-
Taha EKA (2015) Chemical composition and amounts of mineral elements in honeybee-collected pollen in relation to botanical origin. J Apic Sci 59(1): 75-81.
-
Haydak MH (1958) Pollen-pollen substitutes-beebread. Am Bee J 98: 145-146.
-
Amro A, Younis M, Ghania A (2020) Physiological effects of some pollen substitutes diets on caged honey bee workers (_Apis mellifera_ L.). International Journal of Environment 9(1): 87-99.
-
Nicolson SW Human H (2013) Chemical composition of the ‘low quality ‘pollen of sunflower (_Helianthus annuus_, Asteraceae). Apidologie 44: 144-152.
-
Al Qarni AS (1995) Morphometrical and biological studies of the native honeybee race _Apis mellifera_ L.; the carniolan AM carnica Pollmann and their F1 hybrid. M.Sc. Thesis, Fac Agric King Saud University, Riyadh, Saudi Arabia.
-
Al Qarni AS, Balhareth HM, Owayss AA (2014) Performance evaluation of indigenous and exotic honeybee (_Apis mellifera_ L.) races in Assir region, Southwestern Saudi Arabia. Saudi J Biol Sci 21(3): 256- 264.
-
Camazine S (1993) The regulation of pollen foraging by honeybees: how foragers assess the colony’s need for pollen. Behav Ecol Sociobiol 32: 265-272.
-
Pankiw T, Tarpy DR, Page RE (2002) Genotype and rearing environment affect honeybee perception and foraging behaviour. Anim Behav 64(4): 663-672.
-
Schneider SS, McNally LC (1993) Spatial foraging patterns and colony energy status in the African honey bee, _Apis mellifera_ scutellata. J Insect Behav 6: 195-210.
-
Mladenovic M, Miadan V, Dugalic Vrndic N (1999) Effects of a vitamin mineral preparation on development and productivity of bee colonies. Acta Vet 49(2/3): 177-184.
-
Jevtic G, Mladenovic M, Andelkovic B, Nedic N, Sokolovic B, et al. (2009) The correlation between colony strength, food supply and honey yield in honeybee colonies. Biotechnol Anim Husb 25: 1141-1147.
-
Radev ZR (2015) The impact of the protein content of pollen on the development, reproduction and productivity of honeybee (_Apis mellifera_ L.). Ph.D. Thesis, Fac Agric Forest Nat Env Aristotle University of Thessaloniki, Greece.
-
Szczêsna T (2006) Protein content and amino acid composition of bee-collected pollen from selected botanical origins. J Apic Sci 50(2): 81-90.
-
Rogala R, Szymas B (2004) Nutritional value for bees of pollen substitute enriched with synthetic amino acids. Part I. Chemical methods. J Apic Sci 48(1): 19-27.
-
Szczêsna T (2006) Protein content and amino acids composition of bee-collected pollen originating from Poland, South Korea and China. J Apic Sci 50(2): 91-99.
-
Villanueva MO, Marquina AD, Serrano RB, Abellán GB (2002) The importance of bee-collected pollen in the diet: a study of its composition. Int J Food Sci Nutr 53(3): 217-224.
-
Shaw DE (1999) Bees and fungi, with special reference to certain plant pathogens. Australas Plant Pathol 28(4): 269-282.
-
Pernal SF, Currie RW (2001) The influence of pollen quality on foraging behavior in honeybees (Apis mellifera L.). Behav Ecol Sociobiol 51: 53-68.
-
Kosonocka L (1990) Pollen: miracle food or farce. Am Bee J 130: 653-655.
-
Pernal SF, Currie RW (2000) Pollen quality of fresh and 1-year-old single pollen diets for worker honeybees (_Apis mellifera_ L.). Apidologie 31: 387-409.
-
Tasei JN, Aupinel P (2008) Nutritive value of 15 single pollens and pollen mixes tested on larvae produced by bumblebee workers (_Bombus terrestris_, Hymenoptera: Apidae). Apidologie 39: 397-409.
-
Dastagir G, Hussain F, Khattak KF (2014) Nutritional evaluation of plants of family Zygophyllaceae and Euphorbiaceae. Pak J Bot 46(5): 1703-1707.
-
Almaraz Abarca N, Da Graça Campos M, Avila Reyes JA, Naranjo Jimenez N, Corral JH, et al. (2007) Antioxidant activity of polyphenolic extract of monofloral honeybee- collected pollen from mesquite (_Prosopis juliflora_, Leguminosae). J Food Comp Anal 20 (2): 119-124.
-
Agarwal A, Nair PKK (1989) Free and protein-bound amino acids of pollen of Acacia auriculaeformis (Mimosaceae). Grana 28(2): 155-157.
-
Al Zarah AI (2008) Chemistry of groundwater of AI-Alisa Oasis Eastern Region Saudi Arabia and its predictive effects on soil properties. Pak J Biol Sci 11(3): 332-341.
-
Höcherl N, Siede R, Illies I, Gätschenberger H, Tautz J (2012) Evaluation of the nutritive value of maize for honeybees. J Insect Physiol 58(2): 278-285.
-
Lundén R (1956) Literature on pollen chemistry. Grana Palynologica 1: 3-19.
-
Singh RP, Singh PN (1996) Amino acid and lipid spectra of larvae of honeybee (_Apis cerana_ Fabr) feeding on mustard pollen. Apidologie 27(1): 21-28.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind