Molecular Identification of Seven Myxobolus Species (Myxosporea: Myxobolidae) in Cyprinids from India
This communication aims to detect the myxozoan infection in some cyprinid fishes that were mostly cultured and commonly used for food in the Meerut region, Uttar Pradesh, India. Myxozoan were identified morphologically and for molecular analysis we have used previously established PCR assays for genetic marker ssrDNA then phylogenetic analysis was performed. Total seven species of Myxobolus were identified i.e., Myxobolus bhadrensis, Myxobolus kalavatiae, Myxobolus haldari from Labeo rohita, Myxobolus calbasui, Myxobolus catlae, from Cirrihinus reba and Cirrihinus mrigala, Myxobolus hosadurgensis from Cirrihinus mrigala and Myxobolus saranae from Labeo bata. An integrated comparative analysis of the ssrDNA gene supported the identification of the collected species and represents the phylogenetic position of all species. This work addresses the problems in the taxonomy of myxozoans in India, where molecular studies are less focused. My species parasitized similar hosts for which genetic data is the only way to make a clear distinction between species and their validity.
Introduction
Myxozoa Grassé, 1970 are widespread cnidarian parasites and about 2596 species of these have been reported that constitute nearly one-fifth of the known Cnidarian species [1, 2]. Among them, more than 850 Myxobolus Bütschli, 1882 species are identified [3, 4] and from India contribution of about 130 Myxobolus species has been reported [5]. The number of myxozoan species is increasing in India [6, 7, 8, 9, 10]. Genus Myxobolus is characterized by ellipsoidal, ovoid or rounded spores in valvular view and biconvex in sutural view with two polar capsules, equal or unequal in size. Shell valves are generally smooth. Sporoplasm is binucleate, often with an iodinophilous vacuole [11].
The identification of myxozoan species is a big challenge in the case where multiple species of the same genus infected the same host, a situation that creates a taxonomic dilemma. Hence, molecular methods are very popular and boomed the myxozoan systematics, providing a more comprehensive resolution to their morphology and identification [6, 8, 12, 13]. Although in India a large number of myxozoan species have been described from carp hosts, which are mostly based on the spore morphology using traditional taxonomy methods [14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35]. Many of these described species have insufficient data that need to be revalidated or revised morphologically and should be supplemented with molecular characteristics.
Therefore, the aim of this study is to describe species based on morphological data and to evaluate the phylogenetic affinities of Myxobolus species infecting Indian carp fishes based on ssrDNA gene in order to resolve the uncertainties in their identification.
Materials and Methods
Parasite Collection and Morphology
Carp fish were sampled from the Parikshitgarh and Kheri Manihar of district Meerut and Bairaj of district Bijnour of Uttar Pradesh, India in January and February 2017 (n = 58; total length 11-16 cm) and June to August 2018 (n =74; total length 12-15 cm) respectively. Information regarding the collections of specimens of myxozoans is presented in Table 1. Fish were brought to the laboratory at the Department of Zoology, Chaudhary Charan Singh University, Meerut, UP, India and kept in an aerated aquarium. To check myxozoan infections they were euthanized with clove oil and dissected to check various organs like gill filaments, kidney, liver, intestine, gall bladder and muscles for myxozoan plasmodia/ cysts under a Motic stereomicroscope (SMZ-168 series). Plasmodia were carefully removed from the gills, liver, kidney and intestine, opened with a fine needle on a slide and observed under an Olympus microscope (CH30) for further morphological analysis. Measurements based on 25-30 fresh myxospores were taken according to the guidelines of Lom J [36]. All measurements are shown in micrometres (μm) unless stated otherwise also followed by the range (mean ± SD) shown in Table 2, with a comparison to the same species described previously. Some spores were fixed in 95% alcohol and stored at -20°C for subsequent molecular study.
| Sample | Species | Host | Locality | Prevalence and Intensity of Infection | Gills | Liver | Kidney | Intestine | GenBank Accession Nos. |
|---|---|---|---|---|---|---|---|---|---|
| LR1 | Myxobolus bhadrensis (Mbh) | Labeo rohita | Parikshitgarh | 52 % (22 infected out of 42), Moderate | Mbh | MN994379, MN994417 | |||
| LR2 | Myxobolus kalavatiae (Mka) | L. rohita | Parikshitgarh | 88 % (37 infected out of 42), High | Mka | MN994420, MN994422 | |||
| LR3 | Myxobolus haldari (Mha) | L. rohita | Parikshitgarh | 78.6 % (33 infected out of 42), High | Mha | MT002391, MT002392 | |||
| LB1 | Myxobolus saranae (Msa) | Labeo bata | Bijnour | 76.6 % (23 infected out of 30), High | Msa | - | MT002911, MT002914 | ||
| CM1 | Myxobolus hosadurgensis (Mho) | Cirrhinus mrigal | Parikshitgarh | 48.8 % (21 infected out of 43), Low | Mho | MT002915, MT002924 | |||
| CM2 | Myxobolus catlae (Mcat) | C. mrigal | Parikshitgarh | 69.7 % (30 infected out of 43), Moderate | Mcat | MT003664, MT002743 | |||
| CM4 | Myxobolus calbasui (Mcal) | Cirrhinus reba | Kheri Manihar, Mawana | 64.7 % (11 infected out of 17), Moderate | Mcal | MT009485, MT012423 | |||
| CM5 | Myxobolus catlae (Mcat) | C. reba | Kheri Manihar, Mawana | 52.9 % (09 infected out of 17), Moderate | Mcat | MT002747, MT002748 | |||
| CM6 | Myxobolus calbasui (Mcal) | C. mrigal | Parikshitgarh | 60 % (26 infected out of 43), Moderate | Mcal | MT012462, MT012463 |
Table 1: Myxobolus infection present or absent in the organs of carps examined from India.
| Species | Host | Infection site | Size of PC | LOS | WOS | TOS | LLPC | LSPC | WLPC | WSPC | NFC | PFL | Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M. bhadrensis | L. rohita | Kidney | Unequal | 11.02-12.48 (11.87±0.44) | 7.28- 8.58 (7.95±0.38) | 6.45-7.02 (6.74±0.21) | 4.16-5.72 (4.86±0.46 ) | 2.86-4.16 (3.54±0.37) | 1.82-2.86 (2.39±0.29) | 1.56-2.35 (1.94±0.25) | 4(L) 3 (S) | na | Present study |
| M. bhadrensis | L. rohita | Muscles | Unequal | 9.2-10.4 (10.0±0.41) | 6.0-7.2 (6.6±0.37) | 4.0-5.3 (4.5±0.56) | 4.8-6.0 (5.5±0.37) | 3.6-4.8 (4.2±0.41) | 2.6-3.3 (3.0±0.34) | 1.6-2.6 (2.0±0.29) | 4(L) 3 (S) | na | Szekely, et al. [6] |
| M. bhadrensis | L. rohita | Muscles | Unequal | 8.0-11.0 -9.5 | 7.0-8.0 -7.14 | na | 3.0-4.0 (3.5) | 2.0-4.0 (2.5) | 1.0- 2.0 -1.75 | 1.0- 2.0 -1.75 | na | na | Seenappa and Manohar [37] |
| M. calbasui | C. mrigala | Intestine | Unequal | 13.0-14.56 (13.79±0.49) | 8.84-10.66 (9.79±0.61) | 6.76-7.8 (7.31±0.28) | 4.68-5.98 (5.16±0.39) | 2.34-3.02 (2.69±0.21) | 3.12-3.9 (3.5±0.22) | 1.3-2.34 (1.87±0.34) | 5-6 (L) 3-4 (S) | na | Present study |
| M. calbasui | C. reba | Liver | Unequal | 14.3-15.23 (14.83±0.29) | 9.88-10.92 (10.39±0.27) | 6.8-7.7 (7.3±0.26) | 5.2-6.76 (5.9±0.49) | 2.6-3.48 (3.03±0.27) | 2.6-3.9 (3.31±0.41) | 1.52-2.20 (1.85±0.24) | 6-7 (L) 4-5 (S) | na | Present study |
| M. calbasui | C. mrigala, L. rohita, L. bata, and L. dyocheilus | Gill, Scales, Fins, Gall bladder | Unequal | 12 - 15 (13.32) | 8 - 10 (8.94) | na | 5 - 7 (6) | 3 - 4 (3.56) | 3 – 5 -4.18 | 2 - 3 (2.32) | 7 (L) Not seen (S) | 52-64 (L) 16-24 (S) | Gupta, et al. [38] |
| M. calbasui | L. calbasu, L. rohita and C. mrigala | Gall bladder | Unequal | 12.4-15 | 8.2-10.0 | na | 6.18 | 4.12 | 4.12 | 3.09 | na | 125 (L) 64 (S) | Chakravarty, et al. [39] |
| M. opthalmu sculata | C. mrigala | Eye muscles | Unequal | 12.4-13.97 (13.13) | 7.45-9.18 (8.04) | na | 4.9-6.12 (5.47) | 2.35-3.37 (3.03) | 2.65-3.37 (3.06) | 1.84-2.14 (1.99) | 7-9 (L) 2-3 (S) | 53.04- 114.24 (83.13) (L) 20.4-32.64 (24.94) (S) | Basu, et al.[40] |
| M. mrigalhitae | C. mrigala- L. rohita hybrid | Gills | Unequal | 10.8-11.3 (10.8) | 7.6-8.1 (7.9) | na | 4.3-5.2 (4.8) | 2.9-3.2 (3.0) | 2.7-3.2 (2.1) | 2.0-3.2 (2.1) | 5-6 (L) 3-4 (S) | na | Basu, et al. [41] |
| M. buccoroofus | L. bata | Roof of buccal cavity | Unequal | 11.6-12.7 (12.1) | 6.4-8.1 (7.10 | na | 4.5-5 (4.9) | 2.0-.2.9 (2.5) | 2.7-3.0 (2.9) | 1.3-1.7 (1.5) | 6-8 (L) 4-5 (S) | na | Basu, et al. [42] |
| M. patialensis | L. rohita | Caudal fin | Unequal | 10.74-11.82 (11.28±0.76) | 6.27-7.07 (6.67±0.56) | na | 4.6-5.0 (4.8±0.28) | 1.6-1.8 (1.7±0.140 | 2.9-3.3 (3.1±0.28) | 1.22-1.8 (1.5±0.41) | 5-6 (L) 2-3 (S) | na | Kaur, et al. [43] |
| M. catlae | C. mrigala | Gills | Equal or slightly unequal | 17.42-19.24 (18.47±0.58) | 7.28-8.32 (7.93±0.32) | 6.74-7.28 (7.0±0.18) | 10.14-11.4 (10.92±0.34) | 10.14-10.92 (10.51±0.26) | 2.88-2.92 (2.92±0.15) | 2.6-2.86 (2.70±0.10) | 15- Nov | na | Present study |
Table 2: Myxobolus infection present or absent in the organs of carps examined from India.
- M. catlae
- C. reba
- Gills
- Equal or slightly unequal
- 18.2-19.12
- (19.12±0.51)
- 7.28-8.06
- (7.70±0.26)
- 6.24-7.02
- (6.61±0.32)
- 11.7-13.5
- (12.75±0.56)
- 11.4-13.0
- (12.10±0.48)
- 2.60-2.86
- (2.70±0.08)
- 2.34-2.60
- (2.48±0.10)
- 15-
- Nov na
- Present study
- M. catlae
- C. cirrhosus
- Gills
- Equal or slightly unequal
- 16.6-17.6
- (17.1±0.45)
- 6.5-6.65
- (6.6±0.05)
- 4.8-6.0
- (5.4±0.47)
- 8.4–11.2
- (10.4±0.47)
- 8.4–10.6
- (9.9±0.62)
- 2.5–2.6
- (2.5±0.05)
- 2.3–2.4
- (2.4±0.05)
- 15-
- Nov na
- Szekely et al. [6]
- M. catlae
- C. catla, L. rohita and
- Gills
- Equal
- 14.5-16.5
- 6.18
- 5.15
- 10.3-12.36
- 2.06-3.01 na
- 150
- Chakravarty, et al.
- [44]
- M. kalavatiae
- L. rohita
- Gills
- Equal
- 11.4-12.74
- (12.12±0.39)
- 5.84-6.76
- (6.34±0.28)
- 4.94-5.46
- (5.22±0.19)
- 4.94-5.72 (5.45±0.23)
- 1.56-1.82 (1.67±0.094)
- 5 na
- Present study
- M. kalavatiae
- L. rohita
- Gills
- Equal
- 7.3±0.27
- (6.8-7.7)
- 5.3±0.5 (4.8-
- 5.8)
- 3.7±0.09
- (3.5-3.8)
- 3.1±0.16 ( 2.9-3.4)
- 1.9±0.18 (1.6-2.0) na na
- Szekely et al. [6]
- Myxobolus haldari
- L. rohita
- Kidney
- Unequal
- 10.7-11.6
- (11.16±0.27)
- 7.9-8.63
- (8.3±0.19)
- 5.52-5.8
- (5.67±0.08)
- 4.6-4.8
- (4.68±0.06)
- 3.56-3.68
- (3.62±0.4)
- 2.24-2.39
- (2.32±0.04)
- 1.86-1.98
- (1.94±0.04)
- 6-7 (L)
- 4-5 (S) na
- Present study
- C. mrigala,
- L. bata, L. dyocheilus,
- L. rohita,
- Fins and gills
- Unequal
- 9.0-10.0
- -9.31
- 7.0-8.5 -6.25 na
- 4.0-5.0 -4.31 2.5-3.0 -2.95
- 2.5-3.0
- -2.97
- 1.5-2.0
- -1.98 na
- 31-70 (L)
- 18-31 (S)
- Gupta, et al. [45]
- M. haldari
- Myxobolus saranae
- L. bata
- Gills
- Unequal
- 10.6-11.96
- (11.29±0.39)
- 7.0-8.1
- (7.52±0.34)
- 5.2-5.8
- (5.51±0.22)
- 4.94-6.2
- (5.52±0.35)
- 3.12-3.9
- (3.52±0.24)
- 2.6-3.38
- (2.92±0.26)
- 1.56-1.86
- (1.7±0.09)
- 5-6 (L)
- 3-4 (S)
- Na
- Present study
- M. saranae
- L. calbasu,
- Puntius sarana
- Gills
- Unequal
- 6.0-9.0
- (7.72)
- 6.0-7.0 (6.2) na
- 4.0-5.0
- (4.24)
- 1.5-3.0 -1.98
- 2.5-4.0
- -3.04
- 1.0-2.0 -1.3 na
- 23-32 (L)
- 5.0-9.0 (S)
- Gupta, et al. [46]
- M. saranae
- L. rohita
- Caudal fin Unequal
- 8.0-9.0
- (8.5±0.70)
- 5.5-6.5
- (6.0±0.70) na
- 3.97-4.57
- (4.27±0.42)
- 1.9-2.5
- (2.2±0.42)
- 2.31-2.91
- (2.61±0.42)
- 1.74-2.14
- (1.94±0.28)
- 5-6 (L)
- 2-3 (S) na
- Kaur et al. [47]
- M. harpreetae
- L. bata
- Gill lamellae
- Unequal
- 11.4–13.8
- -13.2
- 4.9–7.8 -6.51 na
- 8.7–9.2
- -9.149
- 7.6–8.3 -7.94
- 2.9–3.4
- -3.45
- 2.7–3.0 -2.9 na na
- Ghosh, et al. [48]
- Myxobolus hosadurgensis C. mrigala
- Kidney
- Unequal 12.37-13.52
- (12.86±0.41)
- 7.54-8.32
- (7.94±0.25)
- 5.46-5.98
- (5.7±0.17)
- 4.65-5.3
- (4.9±0.23)
- 2.96-3.44
- (3.18±0.15)
- 2.6-3.1
- (2.85±0.15)
- 1.56-1.82
- (1.7±0.10)
- 5-6(L)
- 3-4(S) na
- Present study
- M. hosadurgensis C. mrigala
- Muscles Unequal
- 9.0-11.0
- (10.5)
- 5.0-8.0
- (6.25) na
- 4.0-6.0
- (5.37)
- 2.5-4.0 (3.3) 2.0-3.0 (2.3)
- 1.0-2.5
- (1.43) na na
- Seenappaand
- Manohar [37]
Table 3: Comparative measurements shown as range (mean ± SD) of Myxobolus species collected in the present study with morphological similar species (PC: Polar capsule, LOS:
DNA Isolation and Amplification
For genomic DNA extraction, ethanol-preserved plasmodia/cysts were processed using a QIAGEN DNeasyTM tissue kit (animal tissue protocol, Qiagen, Hilden, Germany) according to the protocol recommended by the manufacturer. The ssrDNA was amplified using the universal primers ERIB1 and ERIB10 [49] at the first round of PCR. The reaction was carried out according to the protocol followed as PCR was carried out in a 25-µl reaction mixture with the following thermocycling profile: denaturation at 95°C for 3 min; followed by 35 cycles of amplification at 95°C for 45 s, 56°C for 1 min and 72°C for 1 min, completed with terminal extension at 72 °C for 7 min and then stored at 4 °C. This was followed by a second round of PCR with primers Myx1F- SphR [50, 51]. The total volume of the reactions was 50 µl with the following PCR amplification profile: 95 °C for 3 min, then 35 cycles at 95 °C for 50 s, 56 °C for 50 s, 72 °C for 1 min, terminated with an extension at 72 °C for 7 min and then resting at 4 °C. The PCR amplification products were electrophoresed in 1% agarose gel (Tris-Acetate-EDTA buffer) stained with 1% ethidium bromide and examined under ultraviolet light. PCR products were purified with the PurelinkTM Quick Gel Extraction and PCR Purification Combo Kit (Invitrogen). Amplicons were sequenced from both strands with the PCR primers mentioned above using the Big Dye Terminator vr. 3.1 cycle sequencing kit in ABI 3130 Genetic Analyzer (Applied Biosystems, Foster City, CA).
Phylogenetic Analysis
Obtained contiguous sequences were manually assembled, edited using BioEdit, version 7.2.5 [52] checked and submitted to GenBank for accession numbers and put forward to NCBI for BLASTn and comparison with other myxozoan sequences. Newly generated sequences of the ssrDNA of myxozoan species were aligned together with published sequences of myxozoan species, available on GenBank. The sequences were aligned by using Clustal W [53] implemented in the MEGA X [54]. The evolutionary history was analyzed by using the Maximum Likelihood method and the General Time Reversible model. The nodal support was estimated by bootstrapping (n=1,000). The best evolutionary model of HYPERLINK "https://www. sciencedirect.com/topics/biochemistry-genetics-and- molecular-biology/nucleotide"nucleotide substitution using the Akaike Information Criterion (AICs) was determined with the MEGA X. A discrete Gamma distribution was used to model evolutionary rate differences among sites (5 categories (+G, parameter = 0.4686)). Genetic distance estimation was carried out using the p-distance model of substitution in MEGA X. A total of 59 nucleotide sequences with over 91% similarity were used for phylogenetic analysis. There were a total of 2075 positions in the final dataset. Evolutionary analyses were conducted in MEGA X. The Chlorormyxum cristatum (AY604198) was selected as out-group in the final analysis.
Results
Seven Myxobolus species were collected from the locally available cyprinids fish and we have not recorded any external symptoms of infection. All Myxobolus sp. were collected from gills, liver, gall bladder, kidney and intestine. The collected species were morphologically identified as belonging to the genus Myxobolus Bütschli, 1882 (Myxozoa: Myxobolidae) as Myxobolus bhadrensis, Myxobolus calbasui, Myxobolus catlae, Myxobolus kalavatiae, Myxobolus haldari, Myxobolus hosadurgenesis and Myxobolus saranae Figures 1 & 2. Details regarding comparative morphometrical data of the above-mentioned species with other related species are presented in Table 2.
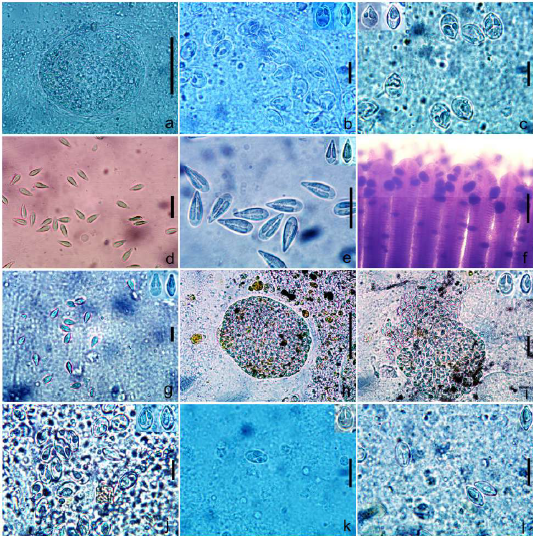
Figure 1: a. Cyst of Myxobolus bhadrensis. b. Spores of Myxobolus bhadrensis. c. Spores of Myxobolus calbasui. d, e. Spores of Myxobolus catlae. f. Gills with cyst of Myxobolus kalavatiae. g. Spores of Myxobolus kalavatiae. h. Cyst of Myxobolus haldari i. Spores of Myxobolus haldari j. Spores of Myxobolus saranae. k, l. Spores of Myxobolus hosadurgensis. Scale bars. a,h=50 µm; b=12 µm; c,g=15 µm; d,e=20 µm; f=200 µm; i,j=10 µm; k,l=15 µm.
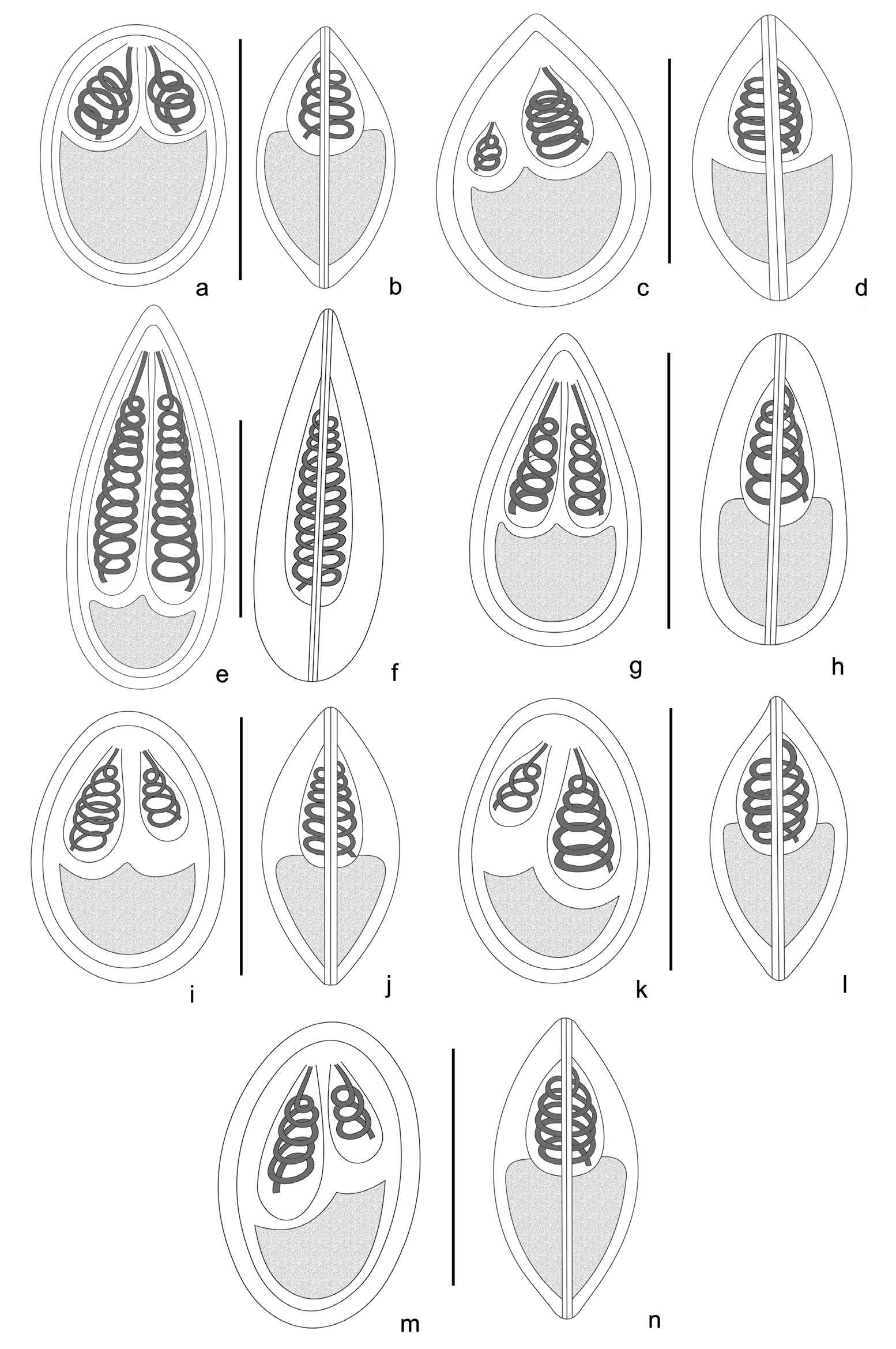
Figure 2: Line drawings of Myxobolus parasites collected during the study. a. Frontal view of Myxobolus bhadrensis. b. Sutural view of Myxobolus bhadrensis. c. Frontal view of Myxobolus calbasui. d. Sutural view of Myxobolus calbasui. e. Frontal view of Myxobolus catlae. f. Sutural view of Myxobolus catlae. g. Frontal view of Myxobolus kalavatiae. h. Sutural view of Myxobolus kalavatiae. i. Frontal view of Myxobolus haldari. j. Sutural view of Myxobolus haldari. k. Frontal view of Myxobolus saranae. l. Sutural view of Myxobolus saranae. m. Frontal view of Myxobolus hosadurgensis. n. Sutural view of Myxobolus hosadurgensis. Scale bars. a-n =10 µm.
Myxobolus bhadrensis Seenappa and Manohar [37], Szekely, et al. [6]
Spores description. Spores oval to ellipsoidal in frontal view with anterior and posterior rounded ends. In sutural view, spores pyriform at both the ends and convex at both sides. Length of spore 11.02-12.48 (11.87±0.44) (N=30); width, 7.28- 8.58 (7.95±0.38) (N=30) and thickness, 6.45- 7.02(6.74±0.21) (N=10). Polar capsules pyriform shape and unequal in size. Larger capsule 4.16-5.72 (4.86±0.46) long (N=30) and 1.82-2.86 (2.39±0.29) wide (N=30) while smaller capsule 2.86-4.16 (3.54±0.37) long (N=30) and 1.56-2.35 (1.94±0.25) wide (N=30). 3 polar tubule coils in smaller capsule and 4 coils in larger capsule. Sporoplasm large, binucleated having small iodinophilous vacuole. Intercapsular process not seen (Table 2; Figures 1a,1b & 2a,2b).
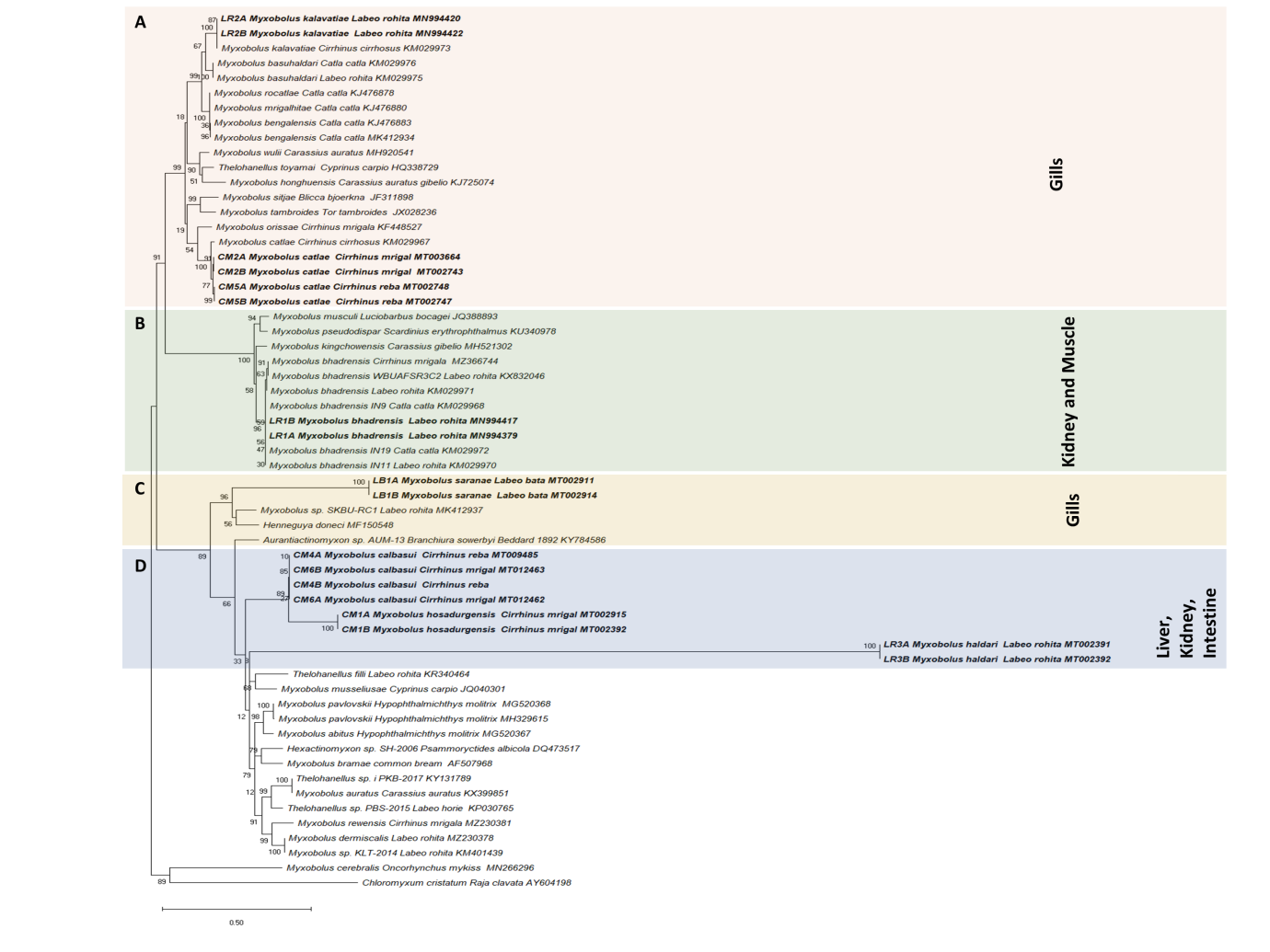
Figure 3: Myxobolus species phylogenetic relationships inferred by maximum likelihood analysis based on the ssrDNA sequences. Species in bold were newly sequenced in this study. GenBank accession numbers and their host name are listed along the species names. Numbers at nodes indicate the bootstrap confidence values (ML). Chloromyxum cristatum was used as an outgroup.
Present host and locality: Labeo rohita, Local name: Rohu; locality: Parikshitgarh Fish Farm (28°59′ N, 77° 56′ E), Meerut (U.P.), India.
Site of infection: Kidney.
Prevalence of infection: 22/42 in the 9-14 cm size group with a prevalence of 52%.
Material deposited: Digitized photographs were deposited in the Museum, Department of Zoology, Chaudhary Charan
Singh University, Meerut, under voucher number HSS/ZOO/ MYX/03/19.
Representative DNA sequence: The ssrDNA sequences of M. bhadrensis were submitted to NCBI with accession numbers MN994379 and MN994417.
• Remarks Spores of M. bhadrensis showed close resemblance in shape and size with spores of other Myxobolus species having unequal sized polar capsules. Spores of M. Mrigalae Chakravarty [39] showed morphological similarities with spores of M. bhadrensis but presence of several triangular marking in the valve of M. Mrigalae differentiates these two species. Spores M. indicum Tripathi [55] also resemble in size and shape but marked difference is seen in the size of polar capsules of both. Morphologically spores of M. bhadrensis shows close resemblance with spores of M. hosadurgensis Seenappa and Manohar [37] but spores of M. bhadrensis were oval to ellipsoidal and wider than spores of M. hosadurgensis which were elongated ovoidal in shape with anterior narrow end. Spores of M. vedavatiensis Seenappa and Manohar [37], M. lalithae Gupta, et al. [56] and M. haldari Gupta, et al. [45] also showed resemblance with spores of M. bhadrensis but spores of M. lalithae were oval or elliptical in shape while spores of M. vedavatiensis and M. haldari had an intercapsular process or ridge.
Myxobolus calbasui Chakravarty [21]
Spores description. Spores large size, rounded-oval in shape, anterior end pointed and posterior end much rounded in frontal view. Spores pyriform in sutural view with both sides convex. Length of spores 13.0-14.6 (13.8±0.4), width 8.8-10.7 (9.8±0.61), thickness 6.76-7.8 (7.3±0.28). Polar capsules oval-pyriform in shape, unequal sized. Larger capsule much larger than smaller capsule, showed marked differentiation in size, and reaches up to the anterior end of spore, but smaller capsule lies at some distance below the larger capsule. Larger capsule 4.68-6.0 (5.2±0.39) long and 3.12-3.9 (3.49±0.22) wide. Smaller capsule 2.34-3.0 (2.7±0.21) long and 1.3-2.3 (1.86±0.35) wide. Polar tubule coils seen 5-6 in larger capsule and 3-4 in smaller capsule in host fish C. mrigala while in spores obtained from C. reba no. of coils seen 6-7 in larger capsule and 4-5 in smaller capsule (Table 2; Figures 1c & 2c,2d).
Present host and locality: Cirrhinus mrigala (Ham.1822), Local name: Mrigal; locality: Parikshitgarh Fish Farm (28º59′N, 77°56′E), Meerut and C. reba (Ham. 1822), Local name: Raia; locality: Village Kheri Manihar Fish Pond (29º10′N, 77°92′E), Mawana, Meerut (U.P.), India.
Site of infection: Cirrhinus mrigala and C. reba: intestine and liver respectively.
Prevalence of infection: 26/43 and 11/17 in the 11-16 cm size group of Cirrhinus mrigala and C. reba, with a prevalence of 60% and 64.7% respectively.
Material deposited: Digitized photographs obtained from both host C. mrigala and C. reba were deposited in the Museum, Department of Zoology, Chaudhary Charan Singh University, Meerut, under voucher number HSS/ZOO/ MYX/11/19 and HSS/ZOO/MYX/10/19 respectively.
Representative DNA sequence: The ssrDNA sequences obtained from both the host C. mrigala and C. reba were deposited in GenBank with accession numbers MT012462, MT012463 and MT009485, MT012423.
• Remarks The characteristic large, rounded-oval spores of this species, with pointed anterior end and much rounded posterior end shows resemblance with those Myxobolus species, having different size polar capsules, infecting the cyprinid fishes. Spores of Myxobolus calbasui were identified on the basis of available morphological and morphometric data. Initially, this species was reported from the gall bladder of L. calbasu, L. rohita and C. mrigala at Calcutta district, West Bengal, India by Chakravarty [39]. Earlier, Ray [57] reported this species from liver of C. mrigala. Later, Gupta [38] (thesis) redescribed this species from different organs of four species of cyprinid fishes, L. rohita, L. bata, L. dyocheilus and C. mrigala. Spores of M. indicum Tripathi [55] described from the liver, intestinal wall and muscles of same host C. mrigala show close similarity in morphological characteristics but further studies are needed for their identification as a synonymous species due to the poor illustration of the spores. Spores of M. opthalmusculata Basu and Haldar [40], M. mrigalhitae Basu and Haldar [41], M. buccoroofus Basu and Haldar [42] obtained from different carp hosts are close in morphology with spores of M. calbasui. But the presence of a small depression at the anterior end of spores of M. opthalmusculata, the presence of parietal folds in the posterior region of the spores of M. mrigalhitae and narrow and bent spores of M. buccoroofus at anterior side, differentiate these species from M. calbasui. Spores of M. patialensis Kaur and Singh [43] are smaller in size with asymmetrical spore valve, having 4-5 parietal folds in the posterior part of spores as compared to spores of M. calbasui.
Myxobolus catlae [45] Szekely, et al. [6]
Spore description. Spores large-sized and elongated in shape. Anterior end of spores pyriform, sharply tapered and blunt in both frontal and sutural view. Posterior ends of spores ovo-rounded in both views. Spores measurement lies between: length 17.4-19.2 (18.5±0.58), width of 7.28-8.3 (7.9±0.32) and thickness 6.74-7.28 (7.00±0.18). Spore valves thin, uniform and symmetrical. Polar capsules (PC) very long, elongated-pyriform shape and filled most of the spore cavity, leaving a small space for sporoplasm at the posterior part of spore. Size of large PC: length 10.14-11.4 (10.92±0.34) and width 2.7-3.12 (2.92±0.15), size of small PC: length 10.14- 10.92 (10.51±0.26) and width 2.6-2.86 (2.70±0.10). Both capsules parallel to the longitudinal axis of body. In most of the spores, polar capsules equal in size but in some spores slightly unequal in size. The number of polar tubule coils ranged from 11 to 15 in each polar capsule. Sporoplasm small, binucleated and lies at the posterior end of the spore cavity. Intercapsular process and mucous envelop were not seen. A small, variable-sized iodinophilous vacuole was also present in the posterior part of the sporoplasm (Table 2; Figures 1d, 1e & 2e, 2f).
Present host and locality: Cirrhinus mrigala (Ham.1822). Local name: Mrigal; locality: Parikshitgarh Fish Farm (28º59′N, 77°56′E), Meerut and C. reba (Ham. 1822). Local name: Raia; locality: Village Kheri Manihar Fish Pond (29º10′N, 77°92′E), Mawana, Meerut (U.P.), India.
Site of infection: Gills lamellae of Cirrhinus mrigala and C. reba.
Prevalence of infection: 30/43 and 09/17 in the 11-16 cm size group of Cirrihinus mrigala and C. reba with a prevalence of 69.7% and 53% respectively.
Material deposited: Digitized photographs obtained from both the host C. mrigala and C. reba were deposited in the Museum, Department of Zoology, Chaudhary Charan Singh University, Meerut, HSS/ZOO/MYX/07/19 and HSS/ZOO/ MYX/08/19 respectively.
Representative DNA sequence: ssrDNA sequences of M. catlae obtained from both the host C. mrigala and C. reba were submitted to NCBI with accession numbers MT002743, MT003664 and MT002747, MT002748 respectively.
• Remarks Spores of M. catlae can be differentiated from other known Myxobolus species easily because of their large, elongated shaped spores with the large number of polar tubule coils in the polar capsules. Identification was confirmed on the basis of morphological as well as the molecular data also. Spores of M. bengalensis Chakravarty, et al. [58] were much smaller in length as compared to spores of M. catlae. In M. cuttaki Haldar, et al. [59]9 number of coils is less as compared to M. catlae while the spores of M. catmrigalae Basu, et al. [42] much broader and have more coils in polar capsules. Spores of M. zoohuri Majumder, et al. [60] were comparatively smaller in size and have more coils in the polar capsule as compared to the M. catlae.
Myxobolus kalavatiae Szekely, et al. [6]
Spore description. Spores medium size, ovoidal, elongated both in frontal and sutural view, with anterior end, tapering and blunt while posterior end broad and rounded. Length of the spore 11.4-12.74 (12.12±0.39) (N=25); width 5.84-6.76 (6.34±0.28) (N=25); and thickness 4.94-5.46 (5.22± 0.19) (N=15). Spore wall thick, uniform and symmetrical. Polar capsules long, pyriform shaped and equal- sized, obliquely located on either side of the midline, filling around more than half of the spore cavity. Capsules 4.94-5.72 (5.45±0.23) long (N=20) and 1.56-1.82 (1.67±0.094) (N=20) wide.Polar tubule coils are 5 in each capsule. Sporoplasm small, binucleate having a small iodinophilous vacuole. Intercapsular process and mucous envelope not seen (Table 2; Figures 1F, 1g & 2g, 2h).
Present host and locality: Labeo rohita (Ham. 1822), local name: Rohu; locality: Parikshitgarh Fish Farm (28º59′N, 77°56′E) Meerut (U.P.), India.
Site of infection: Gills.
Prevalence of infection: 37/42 in the 9-14 cm size group with a prevalence of 88%.
Material deposited: Digitized photographs were deposited in the Museum, Department of Zoology, Chaudhary Charan Singh University Meerut, under voucher number HSS/ZOO/ MYX/04/19.
Representative DNA sequence: The ssrDNA sequences were submitted to NCBI with accession numbers MN994420 and MN994422.
• Remarks Originally Szekely, et al. [6] collected spores of M. kalavatiae from gills of the host fish C. cirrhosus but we have collected spores of M. kalavatiae from the gills of the host fish L. rohita. As compared to the original description, spores were larger in size, having polar tubule coils, which were not seen by Székely, et al. [6]. Comparison of spores of M. kalavatiae was done with other Myxobolus sp. having equal polar capsules. Comparison of morphological characters of M. kalavatiae showed that it is different from M. shetti Seenappa and Manohar [37], M. venkateshi Seenappa and Manohar [37], M. basuhaldari Szekely, et al. [6], M. meerutensis Szekely, et al. [6]. As spores of M. shetti were smaller and broader than spores of M. kalavatiae, having a small intercapsular process and lacking the polar tubule coils. Spores of M. venkateshi were egg-shaped with a well-developed intercapsular process. Spores of M. basuhaldari showed close resemblance with spores of M. kalavatiae, as both have two equal size polar capsules, equipped with 5 coils, except spores of M. basuhaldari were smaller in size and ovoidal in shape and had a small knob at the anterior end, as compared to the spores of M. kalavatiae. In the original description given by Székely, et al. [6] these two spp. showed close resemblance in their size but spores of M. kalavatiae, obtained in the present study were longer and elongated than the spores of M. basuhaldari. In sutural view spores of M. meerutensis are lemon-shaped while spores of M. kalavatiae are elongated. No. of polar tubule coils seen 6 in M. meerutensis and 5 in M. kalavatiae. A small knob-like intercapsular appendix was also present in the spores of M. meerutensis but not seen in the spores of M. kalavatiae.
Myxobolus haldari Gupta, et al. [45]
Spores description. Spores long, ovoidal or elliptical in frontal view, biconvex in sutural view with round anterior and posterior ends. Length of spore 10.7-11.6 (11.16±0.27) (N=25), width 7.9-8.63 (8.3±0.19) (N=25) and thickness 5.52-5.8 (5.67±0.08) (N=15). Polar capsules (PC) unequal sized and pyriform shaped. Large PC length 4.6-4.8 (4.7±0.06) (N=25); width - 2.24-2.39 (2.32±0.04) (N=25) and small PC length 3.56-3.68 (3.63±0.05) (N=25); width 1.84-2.0 (1.94±0.05) (N=25). Number of polar tubule coils 4-5 in smaller capsule, while 6-7 coils in larger capsule. Intercapsular process was not seen (Table 2; Figures 1h, 1i & 2i, 2j).
Present host and locality: Labeo rohita (Ham.1822), local name: Rohu; locality: Parikshitgarh Fish Farm (28º59′N, 77°56′E), Meerut (U.P.), India.
Site of infection: Kidney.
Prevalence of infection: 33/42 in 9-14 cm size group with a prevalence of 78.6%.
Material deposited: Digitized photographs were deposited in the Museum, Department of Zoology, Chaudhary Charan Singh University, Meerut, under voucher number HSS/ZOO/ MYX/05/19.
Representative DNA sequence: The ssrDNA sequences of Myxobolus haldari were submitted to NCBI with accession numbers MT002391 and MT002392.
• Remarks Spores of Myxobolus haldari were found very close to the spores of M. haldari in morphometric traits and other Myxobolus species with unequal polar capsules. Spores of Myxobolus haldari can be differentiated from spores of M. mrigalae Chakravarty [39], M. seshadri Lalita Kumari [61], M. carnaticus Seenappa and Manohar [62] and M. vedavatiensis Seenappa and Manohar [37] due to their size differences and by the presences of several triangular markings in shell valve of M. mrigalae, presences of inter capsular ridge in spores of M. seshadri, M. carnaticus and M. vedavatiensis. Spores of Myxobolus haldari and M. bhadrensis Seenappa and Manohar [37] showed differences in the length of their polar capsules.
Myxobolus saranae Gupta, et al. [46]
Spore description. Spores elongated, oval-shaped with blunt- oval anterior and posterior ends in frontal view. In a sutural view, spores lentiform or convexo-convex in shape. Spores 10.6-11.96 (11.3±0.39) long (N=25); 7.0- 8.1 (7.52±0.34) wide (N=25) and 5.2-5.8 (5.5±0.22) thick (N=15). Polar capsules unequal sized, longer, pyriform or pear-shaped, converging towards the anterior end of spores. Larger capsule 4.94-6.2 (5.52±0.35) long (N=25) and 2.6- 3.38 (2.92±0.26) wide (N=25) whereas length of small polar capsule 3.12-3.9 (3.52±0.24) (N=25) and width 1.56-1.86 (1.7±0.09) (N=25). Polar tubule coils were seen in both polar capsules and no. of coils ranges 5-6 in larger capsule and 3-4 in smaller capsule. Sporoplasm small, cresentric shaped, uni or binucleated. Intercapsular process and mucous envelop not seen. A variable-sized iodinophilous vacuole was seen in the posterior part of sporoplasm (Table 2; Figures 1j & 2k, 2l).
Present host and locality: Labeo bata (Ham. 1822), Local name: Bata; locality: Bairaj, Bijnor, (29°23′ N, 79°11′ E), (U.P.), India.
Site of infection: Gills.
Prevalence of infection: 23/30 in the 15- 20 cm size group with a prevalence of 76.6%. Material deposited: Digitized photographs deposited in the Museum, Department of Zoology, Chaudhary Charan Singh University, Meerut, under voucher number HSS/ZOO/ MYX/06/19.
Representative DNA sequence: The ssrDNA sequences of Myxobolus saranae are submitted to NCBI with accession numbers MT002911 and MT002914.
• Remarks The morphometric details were similar to the M. saranae, which was described by Gupta, et al. [46] from the gills of Puntius sarana and Labeo calbasu. Later on, Kaur, et al. [47] re-described it from caudal fin of L. rohita. ssrDNA sequences generated from Myxobolus saranae did not resemble other available myxosporean sequences at the GenBank. Morphologically M. saranae shows some similarities with other Myxobolus species as Spores of M. mrigalae Chakravarty [39] have several triangular markings on the shell valves as compared to the spores of M. saranae. The presence of intercapsular ridge in spores of M. seshadri Lalitha Kumari [61] makes a differentiation in both the species. Spores of M. bhadrensis Seenappa and Manohar [37] were broader and its polar capsules were smaller than M. saranae. Spores of M. vedavatiensis Seenappa, et al. [37] larger in size and possesses a large inter capsular ridge. Spores of M. chakravartyi Halder, et al. [63] were larger in size with anterior flat end and a triangular thickening at the anterior end. In M. chakravartyi number of polar filament coils 8-9 in the larger capsule and 6-7 in the smaller capsule. Spores of harpreetae Ghosh, et al. [48] have long and less wide spores than M. saranae. Polar capsules of Myxobolus saranae are smaller in length as compared to M. harpreetae.
Myxobolus hosadurgensis Seenappa and Manohar [37]
Spores description. Spores long, ovoid or ellipsoidal in frontal view and biconvex in sutural view with round anterior and posterior ends. Length of spore 12.37-13.52 (12.86±0.41) (N=25), width 7.54-8.32 (7.94±0.25) (N=25) and thickness 5.46-5.98 (5.70±0.17) (N=10). Polar capsules (PC) unequal sized and pyriform in shape. Large PC length 4.65-5.3 (4.9±0.23) (N=25), width 2.66-3.1 (2.86±0.14) (N=25) and small PC length 2.96-3.43 (3.18±0.15) (N=25), width 1.56-1.82 (1.71±0.09) (N=25). Number of polar tubule coils, 3 in smaller polar capsule and 5 in larger polar capsule. Sporoplasm binucleated, having a small iodinophilous vacuole. Intercapsular process was not seen (Table 2; Figures 1k, 1l & 2m, 2n).
Present host and locality: Cirrhinus mrigala (Ham.1822) Local name: Mrigal; locality: Parikshitgarh Fish Farm (28°59′ N, 77° 56′ E), Meerut (U.P.), India.
Site of infection: Kidney.
Prevalence of infection: 21/43 in the 11-16 cm size group with a prevalence of 48.8%.
Material deposited: Digitized photographs were deposited in the Museum, Department of Zoology, Chaudhary Charan Singh University, Meerut, under voucher no. HSS/ZOO/ MYX/09/19.
Representative DNA sequence: ssrDNA sequences of Myxobolus hosadurgensis are submitted to NCBI with accession numbers MT002915 and MT002924.
• Remarks The spores of Myxobolus hosadurgensis showed close resemblance in morphometric traits with spores of Myxobolus hosadurgensis Seenappa and Manohar, 1981. Because of unequal sized polar capsules M. hosadurgensis showed its morphological similarities with M. mrigalae Chakravarty [39], M. pinnaurati Lalitha Kumari [61], M. carnaticus Seenappa and Manohar [37], M. bhadrensis Seenappa and Manohar [37] and M. vedavatiensis Seenappa and Manohar [37] with some difference in their size, shape and number of polar tubule coils in polar capsules. However, recently generated ssrDNA sequences of this species did not match with any other previous myxozoan sequences available at NCBI.
Phylogenetic Analysis
Molecular analysis of the ssrDNA gene confirmed the phylogenetic position of the studied species (Figure 3). Maximum likelihood (ML) generated a well-resolved tree topology and bootstrap values are presented (Figure 3).
Myxobolus kalavatiae infecting the gills of rohu in the present study shows that ssrDNA sequences of both isolates did not show any intraspecific divergence and shared high identity (99.4%) with other isolates of the same species described by Székely, et al. [6] from gills of mrigal (Cirrihinus cirrohosus) (Figure 3). The phylogenetic analyses inferred from ssrDNA data set also show that M. kalavatiae is a sister to isolates of M. basuhaldari with strong homology (96.4%) reported from the gills of rohu and katla by Székely C, et al. [6]. The present phylogenetic analysis shows that M. kalavatiae was positioned within the well-supported clade that belongs to the gill infecting species in a clade A. Myxobolus catlae collected in the present study from gills of mrigal and reba was originally described by Chakravarty, 1943 from various hosts katla, rohu and mrigal. Later M. catlae was also reported by Székely, et al. [6] from the gills of Cirrihinus cirrhosus with a more comprehensive description. In the present study, we have collected M. catlae from mrigal and reba and ssrDNA sequences of these M. catlae isolates exhibited a similarity of 99.8 % with each other collected from two different hosts (Figure 3). Our isolates showed a 98.2 % similarity with M. catlae, which was isolated from the gills of Cirrihinus cirrhosus. In the clade B, Myxobolus bhadrensis ssrDNA sequence of two isolates collected from the kidney of rohu were found to be identical and showed a 99.2 to 99.4% similarity with other isolates of same species reported from the kidney and muscles of rohu, katla reported by Székely C, et al. [6] while showing similarity of 99.1% with isolate WBUAFSR3C2 infecting the kidney of rohu from West Bengal, India (Figure 3). The phylogenetic tree also showed that M. bhadrensis species formed a sister relationship with M. kingchowensis reported from the muscles of Carassius gibelio from China
with maximum homology (96.8 %) (Figure 3). According to the Székely, et al. [6] both the Myxobolus species (M. kingchowensis and M. bhadrensis) shared the same subclade and show affinity to tissue tropism towards the muscle and kidney respectively. Regarding Myxobolus saranae reported from gills of bata (Labeo bata) can be readily differentiated from other Myxobolus species studied here in the study based on molecular data of the ssrDNA gene and placed in the clade C (Figure 3). The tree topology strongly supported the independent branch formed by isolates of Myxobolus hosadurgensis shows no intraspecific variation among isolates. Myxobolus saranae forms a branch close to Myxobolus sp. (MK412937) infecting gills lamellae of rohu (Labeo rohita) shows only 81.3% similarity with each other and both are members of the same host family Cyprinidae (Figure 3).
Myxobolus haldari was originally described from the fins and gills of mrigal, rohu, bata and kali (Labeo dyocheilus) by Gupta, et al. [45]. Nonetheless, the molecular study of this species has not been performed previously. We have collected M. haldari from kidney of rohu and the molecular analysis of the ssrDNA gene confirmed its validation. The phylogenetic placement of M. haldari isolates clustered them together in an independent branch under subclade D and then clustered with sequences of Myxobolus hosadurgensis and M. calbasui isolates, parasites of kidney, liver and intestine of cyprinid host mrigal and reba (Figure 3). The retrieved similarity values of Myxobolus haldari isolates are 87.6% with isolates of Myxobolus hosadurgensis and 85.3% with M. calbasui isolates. Myxobolus hosadurgensis isolates share a common clade with M. calbasui collected in the present work from liver and intestine of reba and mrigal carp respectively in a clade D (Figure 3). Both the species share some morphological similarities as their myxospores have larger and shorter polar capsules. In turn, Myxobolus haldari which forms a branch close to Myxobolus hosadurgensis and M. calbasui have also significantly large and small polar capsules. The genetic distance estimation revealed the highest similarity of Myxobolus spp. to M. calbasui isolates (92.3%) (Figure 3). M. calbasui collected in the present study from the liver and intestine of reba and mrigal both have unequal polar capsules. Originally, this species was described by Chakravarty MM [39] from gall bladder of calbasu (Labeo calbasu), rohu and mrigal with small polar capsules have three to four filament turns while the large ones have five to six turns respectively. In all, M. calbasui isolates of reba and mrigal ssrDNA gene sequences revealed 100% similarity with each other (Figure 3). M. calbasui isolates share a clade with Myxobolus hosadurgensis from the kidney of mrigal and the genetic distance similarity values range 91.8% (Figure 3).
Discussion
The Myxobolus Bütschli, 1882 is the largest genus among the myxosporean and the diversity of this genus keeps increasing day by day all over the world. The phylogenetic placement of the seven species of Myxobolus using ssrDNA sequences in the present study was consistent as the species in the tree are closely related, all infecting the hosts of the family Cyprinidae. Although, previously many studies also proved that myxozoan species generally lean towards to cluster according to the family of the fish host [64, 65, 66] and it can be used as a character to discriminate closely related species. Now a day, it is already accepted that for the identification of myxosporean species morphology alone is not sufficient and the studies should be supplemented with molecular data [6, 8, 21, 28, 67, 68] as many congener species have morphological similarities with each other and cannot be readily distinguishable. However, the unavailability of molecular data (ssrDNA), restricts the genetic and evolutionary comparison of these species. Although, especially in India focus on molecular aspects is less in comparison to morphology and except for a few, the majority of species were described based on morphology only [14, 15, 16, 17].
During the present study, seven Myxobolus species were obtained from the carp fish of the Meerut region. Out of these, three species were collected from the renal tissue (M.bhadrensis, M. haldari, M. hosadurgensis) three from gills (M. kalavatiae, M.saranae, M.catlae,) and one from the liver (M. calabasui) and intestine (M. calabasui). According to Cech, et al. [24] in general Myxobolus species are host specific but may also infect some closely related fishes. Recovery of spores of M. calbasui and M. catlae from two different hosts, C. mrigala and C. reb_a are supported by the findings of Székely [6] which has mentioned that genetically closely related Indian major carps and their hybrids may share the same Myxobolus species. In the molecular phylogeny of Indian fish species, which also included all the carp genera: Labeo, Catla and Cirrhinus show a close phylogenetic relationship as sister species. In the present study Chakraborty M, et al. [69, 70] all the species infected the host of order Cypriniformes and the family _Cyprinidae. In terms of phylogeny, M. kalavatiae and M. catlae despite some genetic diversity with other isolates available on Genbank, are placed in the tree with different isolates along with other species in clade A, that are gill infecting myxobolids. However, M. bhadrensis collected from the kidney of host fish in the present study and previously reported from muscle and kidney of host fish forms a clade B with other muscle infecting species i.e., M. terengganuensis, as kidney and muscle infecting species share a common subclade, also reported in previous studies [28]. According to Molnár K, et al. [71] dispersed spores and large groups of Myxobolus species are commonly found in the renal tissue; both the coelozoic as well as histozoic species are capable of causing kidney pathology. Regarding Myxobolus saranae, it is clustered in a clade C with Myxobolus and Henneguya species forming a group that is composed of exclusively gill infecting parasites of Cypriniformes. In the clade D, where Myxobolus haldari, Myxobolus hosadurgensis and M. calbasui is dispersedly grouped in a cluster primarily of liver, kidney and intestine infecting species, while other gills infecting species belong to Myxobolus and Thelohanellus occurred in another cluster under clade D.
Thus, the findings of the present study corroborate the idea given in previous studies that the host and inclination for a particular organ/tissue development is an important factor for the identification of species that should be considered for myxozoan phylogenetic studies [33, 72]. Eszterbauer E [33] suggested that genetic differentiation based on tissue/organ tropism is a more ancient evolutionary characteristic than host specificity. Phylogenetic relationships related to tissue tropism have been reported for myxobolids as stronger evolutionary signals [67, 73, 74, 75]. Most of the Myxobolus species are host specific but as in our study, one species infected two different hosts as also reported earlier that myxosporeans can infect other closely related fish species [33, 76, 77, 78].
The findings presented above reveal the fact that myxosporean species infected cyprinid hosts are phylogenetically close, so as a side as tissue tropism shows the strongest evolutionary signal, phylogenetic affinity according to the fish hosts also suggests an important factor. Future work of myxosporeans with more molecular data is required to better understand evolutionary relationships and their taxonomy that was previously based purely on morphology for most Indian species.
Conclusion
In this present study, collected Myxobolus species data supported by morphological measurement and phylogenetic analysis of ssrDNA sequences data. The M. saranae, M. hosadurgensis, M. haldari and M. calabasui showed different clades from other available species on Genbank, due to a lack of previous data available on these species. This study adds the knowledge of the diversity of Myxobolus species in cyprinid fishes. Also, proves that Myxobolus species is the most dominant species among other species in cyprinids fishes in India.
- Conflicts of Interest The authors declare no competing interests.
- Acknowledgements We are grateful to the Head, Department of Zoology, Chaudhary Charan Singh University, Meerut _(_Uttar Pradesh), 250004, India, for providing laboratory facilities.
- Ethical Approval All applicable institutional and national guidelines for the care and use of animals were followed.
- Funding This work was supported by a grant from Chaudhary Charan Singh University to the first author (AG) as Research Grant for PhD students.
References
-
Okamura B, Hartigan A, Naldoni J (2018) Extensive Uncharted Biodiversity: The Parasite Dimension. Integr Comp Biol 58(6): 1132-1145.
-
Zhang ZQ (2011) Animal biodiversity: an outline of higher-level classification and taxonomic richness. Zootaxa pp 3148(1): 1-3.
-
Eiras JC, Molnár K, Lu YS (2005) Synopsis of _Myxobolus_ Bütschli, 1882 (Myxozoa: Myxosporea, Myxobolidae). Syst Parasitol 61: 1-46.
-
Eiras JC, Zhang J, Molnár K (2014) Synopsis of the species of Myxobolus Bütschli, 1882 (Myxozoa: Myxosporea, Myxobolidae) described between 2005 and 2013. Syst Parasitol 88: 11-36.
-
Kaur H, Singh R (2012) A synopsis of the species of _Myxobolus_ Bütschli, 1882 (Myxozoa: Bivalvulida) parasitizing Indian fishes and a revised dichotomous key to myxosporean genera. Syst Parasitol 81: 17-37.
-
Székely C, Cech G, Chaudhary A, Borzák R, Singh HS, et al. (2015) Myxozoan infections of the three Indian major carps in fish ponds around Meerut, UP, India, with descriptions of three new species, _Myxobolus basuhaldari_ sp. n., _M. kalavatiae_ sp. n. and _M. meerutensis_ sp. n., and the redescription of _M. catlae_ and _M. bhadrensis_. Parasitol Res 114(4): 1301-1311.
-
Gupta A, Kaur H (2017) A new pathogen, _Myxobolus_ _holzerae_ (Myxosporea: Myxozoa) causing severe gill disease in an Indian major carp Labeo rohita in a cold water wetland, Punjab (India). Microb Pathog 111: 244- 251.
-
Chaudhary A, Goswami U, Gupta A, Cech G, Singh HS, et al. (2018) Morphological, histological, and molecular description of _Myxobolus ompok_ n. sp. (Myxosporea: Myxobolidae), a kidney myxozoan from Pabdah catfish _Ompok pabda_ (Hamilton, 1822) (Siluriformes: Siluridae) in India. Parasitol Res 117(6): 1899-1905.
-
Gupta A, Kaur H (2018) _Myxobolus okamurae_ sp. nov. (Myxosporea: Myxozoa) causing severe gill myxoboliosis in the cyprinid _Labeo bata_ in a cold water wetland, Punjab (India). Microb Pathog 115: 86-92.
-
Ahmed I, Ahmad I, Dar S, Awas M, Kaur H, et al. (2019). _Myxobolus himalayaensis_ sp. nov. (Cnidaria: Myxozoa) parasitising _Schizothorax_ _richardsonii_ (Cyprinidae: Schizothoracinae) from River Poonch in North West Himalaya, India. Aquacult Rep 14: 100192.
-
Lom J, Dykova I (2006) Myxozoan genera: Definition and notes on taxonomy, life-cycle terminology and pathogenic species. Folia Parasitol 53(1): 1-36.
-
Okamura B, Gruhl A, Bartholomew J L (2015) An introduction to myxozoan evolution, ecology and development. In: Okamura BGB, Gruhl A, et al. (Eds.), Myxozoan Evolution, Ecology and Development. Springer International Publishing, Switzerland, pp: 1-20.
-
Zhang B, Zhai Y, Liu Y, Gu Z (2017) _Myxobolus pseudowulii_ sp. n. (Myxozoa: Myxosporea), a new skin parasite of yellow catfish _Tachysurus fulvidraco_ (Richardson) and redescription of _Myxobolus voremkhai_ (Akhmerov, 1960). Folia Parasit 64: 30.
-
Kalavati C, Nandi N C (2007) Handbook on myxosporean parasites of Indian fishes. Zoological Survey of India, Kolkata, New Delhi, India, pp: 1-294.
-
Ahmad I, Kaur H (2017) _Mxyobolus vascularis_ n. sp. (Cnidaria: Myxozoa: Myxosporea), a new parasite infecting fingerlings of Indian major carps in aquaculture in Punjab, India. Bull Pure and Appl Sci 36(2): 57-70.
-
Fariya N, Abidi R, Chauhan UK (2018) _Myxobolus_ _deformis_ sp. nov. (myxozoa, myxosporea, myxobolidae), a new myxosporean parasite infesting the gills of _Cyprinus_ _carpio_. Species 19: 4-14.
-
Ahmad I, Kaur H (2018) Prevalence, site and tissue preference of myxozoan parasites infecting gills of cultured fingerlings of Indian major carps in District Fatehgarh Sahib, Punjab (India). J Parasit Dis 42(4): 559- 569.
-
Abraham TJ, Banerjee S, Patra A, Sarkar A, Adikesavalu H, et al. (2015) Molecular phylogeny of _Myxobolus orissae_ (Myxosporea: Myxobolidae) infecting the gill lamellae of mrigal carp _Cirrhinus mrigala_ (Actinopterygii: Cyprinidae). Mol Biol Res Commun 4(1): 15-24.
-
Kato E, Kasai A, Tomochi H, Li YC, Sato H (2017) Four _Myxobolus_ spp. (Myxosporea: Bivalvulida) from the gill lamellae of common carp (_Cyprinus carpio_) and Japanese silver crucian carp (_Carassius langsdorfii_) in the western part of Japan, with the description of three new species (_M. tanakai_ n. sp., _M. paratoyamai_ n. sp., and _M. ginbuna_ n. sp.). Parasitol Res 116: 2427-2441.
-
Yokoyama H, Ogawa K (2015) The resurrection of _Myxobolus toyamai_ with a validation of a stunted polar capsule based on morphological evidence. Parasit Int 64(4): 43-47.
-
Zhang JY, Yokoyama H, Wang JG, Li AH, Gong XN, et al. (2010) Utilization of tissue habitats by _Myxobolus_ _wulii_ Landsberg & Lom, 1991 in different carp hosts and disease resistance in allogynogenetic gibel carp: redescription of _M. wulii_ from China and Japan. J Fish Dis 33(1): 57-68.
-
Szekely C, Shaharom Harrison F, Cech G, Mohamed K, Zin NA, et al. (2012) Myxozoan infection of the Malaysian mahseer, _Tor tambroides_, of Tasik Kenyir Reservoir, Malaysia: description of a new species _Myxobolus_ _tambroides_ sp. n. Parasitol Res 111(4): 1749-1756.
-
Banerjee S, Patra A, Adikesavalu H, Joardar SN, Abraham TJ (2015) Molecular and phylogenetic characterization of _Thelohanellus_ _qadrii_ (Myxozoa, Myxosporea, Bivalvulida) infecting the secondary gill epithelium of Indian major carp, _Catla catla_ (Hamilton, 1822). Mol Biol Res Commun 4(2): 83-91.
-
Cech G, Molnár K, Székely C (2012) Molecular genetic studies on morphologically indistinguishable _Myxobolus_ spp. infecting cyprinid fishes, with the description of three new species, _M. alvarezae_ sp. nov., _M. sitjae_ sp. nov. and _M. eirasianus_ sp. nov. Acta Parasitol 57(4): 354-366.
-
Cech G, Borzák R, Molnár K, Székely C (2015) Three new species of _Myxobolus_ Bütschli, 1882 (Myxozoa: Myxobolidae) infecting the common nase _Chondrostoma_ _nasus_ (L.) in the River Danube. Syst Parasitol 92(2): 101- 111.
-
Eszterbauer E (2002) Molecular biology can differentiate morphologically indistinguishable myxosporean species: _Myxobolus elegans_ and _M. hungaricus_. Acta Vet Hung 50(1): 59-62.
-
Molnár K, Marton S, Eszterbauer E, Székely C (2006) Comparative morphological and molecular studies on _Myxobolus_ spp. infecting chub from the river Danube, Hungary, and description of _M. muellericus_ sp. n. Dis Aquat Org 73(1): 49-61.
-
Székely C, Shaharom Harrison F, Cech G, Ostoros G, Molnár K (2009) Myxozoan infections in fishes of the Tasik Kenyir Water Reservoir, Terengganu, Malaysia. Dis Aquat Org 83(1): 37-48.
-
Zhang J, Mo X, Li NN, Chen W, Yang CZ (2015) Supplementary description of _Henneguya zikawiensis_ Sikama, 1938 and its molecular phylogeny. Neijiang Shifan Xueyuan Xuebao 4: 42-46.
-
Rajesh SC, Banerjee S, Patra A, Dash G, Abraham TJ (2014) Molecular characterization of _Myxobolus_ _cuttacki_ (Myxozoa, Myxosporea, Bivalvulida) infecting gill lamellae of minor carp _Labeo bata_ (Ham.). Mol Biol Res Commun 3(4): 231-239.
-
Banerjee S, Patra A, Mondal A, Adikesavalu H, Ramudu KR, et al. (2017) Molecular characterization of _Myxobolus_ _catmrigalae_ (Myxosporea: Myxobolidae) infecting the gill lamellae of carp _Cirrhinus mrigala_ (Hamilton). J Parasit Dis 41(1): 62-70.
-
Molnár K, Cech G, Székely C (2008) _Myxobolus_ species infecting the cartilaginous rays of the gill filaments in cyprinid fishes. Acta Parasitol 53: 330-338.
-
Eszterbauer E (2004) Genetic relationship among gill- infecting Myxobolus species (Myxosporea) of cyprinids: molecular evidence of importance of tissue-specificity. Dis Aquat Org 58(1): 35-40.
-
Zhang B, Gu Z, Liu Y (2018) Morphological, histological and molecular characterization of three _Myxobolus_ species (Cnidaria: Myxosporea) from silver carp _Hypophthalmichthys molitrix_ Valenciennes and bighead carp _Hypophthalmichthys nobilis_ Richardson in China. Parasitol Int 67(4): 509-516.
-
Whipps CM, Murray KN, Kent ML (2015) Occurrence of a myxozoan parasite _Myxidium streisingeri_ n. sp. in laboratory zebra fish _Danio rerio_. J Parasitol 101(1): 86- 90.
-
Lom J, Arthur JR (1989) A guideline for the preparation of species descriptions in Myxosporea. J Fish Dis 12(2): 151-156.
-
Seenappa D, Manohar L (1981) Five new species of _Myxobolus_ (Myxosporea: Protozoa), parasitic in _Cirrhinus_ _mrigala_ (Hamilton) and _Labeo rohita_ (Hamilton), with a note on a new host record _M. curmucae_ Seenappa & Manohar, 1980. J Protozool 28(3): 358–360.
-
Gupta S (1987) Histozoic and Coelozoic Myxozoan Parasites (Protozoa: Myxozoa: Myxosporea) of freshwater fishes of Northern India. Unpublished PhD thesis submitted to Panjab University, Chandigarh, pp: 189.
-
Chakravarty MM (1939) Studies on myxosporidia from fishes of Bengal, with a note on myxosporidian infection in aquaria fishes. Arch Protistenk 92: 169-178.
-
Basu S, Haldar DP (2002) _Myxobolous opthalmusculata_, a new species of histozoic myxozoan (Myxozoa: Bivalvulida) from _Cirrhinus mrigala_ (Hamilton). Proc Zool Soc Calcutta 55: 43-48.
-
Basu S, Haldar DP (2003) Three new species of _Myxobolus_ Butschli, 1882 from different food fishes of West Bengal, India. Acta Protozool 42: 245-251.
-
Basu S, Haldar DP (2004) Description of three new species (Myxozoa: Myxosporea: Bivalvulida) of the genera _Myxobilatus_ Davis, 1944 _Myxobolus_ Butschli, 1882. Acta Protozool 43: 337-343.
-
Kaur H, Singh R (2011) Two new species of _Myxobolus_ (Myxozoa: Myxosporea: Bivalvulida) infecting Indian freshwater fishes in Punjab Wetlands (India). Parasit Res 108: 1075-1082.
-
Chakravarty MM (1943) Studies on myxosporidia from the common food fishes of Bengal. Proc Indian Acad Sci 18: 21-35.
-
Gupta S, Khera S (1989) Observations on _Myxobolus_ _haldari_ sp. nov. (Myxozoa: Myxosporea) from freshwater fishes of northern India. Res Bull (Science) Punj Univ 40: 281-291.
-
Gupta S, Khera S (1990) On three species of the genus _Myxobolus_ Butschli, 1882 (Myxozoa: Myxosporea) from freshwater fishes of northern India. Indian J Parasitology 14: 1-8.
-
Kaur H, Dar SA, Singh R (2013) On new and three already known myxosporean parasites of Indian major carps in Punjab (India). Species 4: 17-24.
-
Ghosh S, Bandyopadhyay PK (2017) Morphotaxonomical description of three new species of _Myxobolus_ Bütschli, 1882 (Myxozoa: Myxosporea: Bivalvulida) from edible freshwater fishes of West Bengal, India. J Parasit Dis 41: 97-105.
-
Barta JR, Martin DS, Liberator PA, Dashkevicz M, Anderson JW, et al. (1997) Phylogenetic relationships among eight _Eimeria_ species infecting domestic fowl inferred using complete small subunit ribosomal DNA sequences. J Parasitol 83(2): 262-271.
-
Hallett SL, Diamant A (2001) Ultrastructure and small- subunit ribosomal DNA sequence of _Henneguya lesteri_ n. sp. (Myxosporea), a parasite of sand whiting _Sillago_ _analis_ (Sillaginidae) from the coast of Queensland, Australia. Dis Aquat Org 46(3): 197-212.
-
Eszterbauer E, Székely C (2004) Molecular phylogeny of the kidney parasitic _Sphaerospora renicola_ from common carp (_Cyprinus carpio_) and _Sphaerospora_ sp. from goldfish (_Carassius auratus auratus_). Acta Vet Hung 52(4): 469-478.
-
Hall TA (1999) BioEdit: a user–friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser 41: 95-98.
-
Thompson JD, Higgins DG, Gibson TJ (1994) Clustal W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res 22(22): 4673-4680.
-
Kumar S, Stecher G, Knyaz M Li C, Tamura K (2018) Mega X: molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol 35(6): 1547-1549.
-
Tripathi YR (1952) Studies on the parasites of Indian fishes. І. Protozoa: Myxosporidia together with a Checklist of parasitic protozoa described from Indian fishes. Rec Indian Museum 50: 63-88.
-
Gupta S, Khera S (1988) On one new and one already known species of the genus _Myxobolus_ from freshwater fishes of India. Res Bull (Science) Punj Univ 39: 173-179.
-
Ray HN (1933) Myxosporidia from India. Proc Indian Sci Congr 1: 259.
-
Chakravarty MM, Basu SP (1948) Observations on some myxosporideans parasitic in fishes with an account of nuclear cycles in one of them. Proc Zool Soc Bengal India 1: 23-33.
-
Haldar DP, Samal KK, Mukhopadhyay D (1996) Studies on the protozoan parasites of fishes in Orissa: eight species of _Myxobolus_ Butschli (Myxozoa: Bivalvulida). J Beng Nat Hist Soc 16: 3-24.
-
Majumder S, Panda S, Ghosh S, Bandyopadhyay PK (2015) Description of a new species of _Myxobolus_ Butschli, 1882 from the _Cirrhinus mrigala_ Hamilton, 1822 an edible fish of India. J Parasit Dis 39(3): 456-60.
-
Lalitha Kumari PS (1969) Studies on parasitic protozoa (Myxosporidia) of freshwater fishes of Andhra Pradesh, India. Riv Parasitol 30(3): 153-226.
-
Seenappa D, Manohar L (1980) Two new species of _Myxobolus_ (Myxoporidea: Protozoa) parasitic on _Cirrhinus mrigala_ (Ham.) and _Puntius curmuca_ (Hamilton). Curr Sci 49: 204-206.
-
Haldar DP, Das MK, Sharma BK (1983) Studies on Protozoan parasites from fishes. Four new species of the genera _Henneguya_ Thelohan, 1882, _Thelohanellus_ Kudo, 1933 and _Myxobolus_ Butschli, 1882. Arch Protistenk 127: 283-296.
-
Holzer AS, Bartošová Sojková P, Born Torrijos A, Lövy A, Hartigan A, et al. (2018) The joint evolution of the Myxozoa and their alternate hosts: A cnidarian recipe for success and vast biodiversity. Mol Ecol 27(7): 1651- 1666.
-
Naldoni J, Zattib SA, Silva MRM da, Maia AAM, Adriano EA (2019) Morphological, ultrastructural, and phylogenetic analysis of two novel _Myxobolus_ species (Cnidaria: Myxosporea) parasitizing bryconid fish from São Francisco River, Brazil. Parasitol Int 71: 27-36.
-
Liu Y, Lövy A, Gu Z, Fiala I (2019) Phylogeny of Myxobolidae (Myxozoa) and the evolution of myxospore appendages in the _Myxobolus_ clade. Int J Parasitol 49(7): 523-530.
-
Rocha S, Azevedo C, Alves Â, Antunes C, Casal G (2019) Morphological and molecular characterization of myxobolids (Cnidaria, Myxozoa) infecting cypriniforms (Actinopterygii, Teleostei) endemic to the Iberian Peninsula. Parasite 26: 48.
-
Borkhanuddin MH, Cech G, Molnár K, Shaharom Harrison F, Khoa TND, et al. (2020) _Henneguya_ (Cnidaria: Myxosporea: Myxobolidae) infections of cultured barramundi, _Lates calcarifer_ (Perciformes: Latidae) in an estuarine wetlands system of Malaysia: description of _Henneguya setiuensis_ n. sp., _Henneguya voronini_ n. sp. and _Henneguya calcarifer_ n. sp. Parasitol Res 119(1): 85- 96.
-
Chakraborty M, Ghosh SK (2014) An assessment of the DNA barcodes of Indian freshwater fishes. Gene 537(1): 20-28.
-
Khedkar GD, Jamdade R, Naik S, David L, Haymer D (2014) DNA barcodes for the fishes of the Narmada, one of India’s longest rivers. PloS One 9(7): e101460.
-
Molnár K (2007) Site preference of myxozoans in the kidneys of Hungarian fishes. Dis Aquat Org 78(1): 45-53.
-
Molnár K, Ranzani Paiva MJ, Eiras JC, Rodrigues EL (1998) _Myxobolus macroplasmodialis_ sp. n. (Myxozoa: Myxosporea), a parasite of the abdominal cavity of the characid teleost, _Salminus maxillosus_, in Brazil. Acta Protozool 37: 241-245.
-
Fiala I (2006) The phylogeny of Myxosporea (Myxozoa) based on small subunit ribosomal RNA gene analysis. Int J Parasitol 36(14): 1521-1534.
-
Carriero MM, Adriano EA, Silva MRM, Ceccarelli PS, Maia AAM (2013) Molecular phylogeny of the _Myxobolus_ and _Henneguya_ genera with several new South American species. PLoS One 8 (9): e73713.
-
Molnár K, Eszterbauer E (2015) Specificity of infection sites in vertebrate hosts. In: Okamura BGB, Gruhl A, et al. (Eds.), Myxozoan Evolution, Ecology and Development. Springer International Publishing, Switzerland, pp: 295- 313.
-
Molnár K (1994) Comments on the host, organ and tissue specificity of fish myxosporeans and on the types of their intrapiscine development. Parasitol Hung 27: 5-20.
-
Molnár K, Marton S, Eszterbauer E, Székely C (2006) Comparative morphological and molecular studies on _Myxobolus_ spp. infecting chub from the river Danube, Hungary, and description of _M. muellericus_ sp. n. Dis Aquat Org 73(1): 49-61.
-
Molnár K, Cech G, Székely C (2011) Histological and molecular studies of species of _Myxobolus_ Bütschli, 1882 (Myxozoa: Myxosporea) in the gills of _Abramis_, _Blicca_ and _Vimba_ spp. (Cyprinidae), with the redescription of _M. macrocapsularis_ Reuss, 1906 and _M. bliccae_ Donecet Tozyyakova, 1984. Syst Parasitol 79: 109-121.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind