Water, Feeding Efficiencies, and Development of Dermestes maculatus DeGeer (1774)
Although dermestids have varying ecological roles, most Dermestes are generally associated with decomposition of dry animal tissues. However, Dermestes maculatus DeGeer (1774) can eat soft, hydrated tissues, so it is unclear if the apparent preference of Dermestes maculatus for dry tissue reflects physiological adaptation or competitive exclusion by blowflies and other invertebrate scavengers that arrive at carrion earlier and may have faster immature development rates. To address this question, we examined development of D. maculatus larvae on hydrated and dry food, and we determined development rates at a constant temperature of 25°C. Specifically, we conducted multiple replicated experiments in incubators. Initial experiments used with a factorial treatment arrangement of food type (wet versus dry) and larval density (1, 5, 10, 15, and 25 per 5 g of food) with 4 replications. Additionally, for dry food we determined growth, consumption, and conversion rates for 3-6 stage larvae, and consumption and conversion rates for adults. Hydrated food significantly reduced survivorship of D. maculatus larvae and adults (ca. 20% survivorship) as compared to dried food with survivorship rates of ca. 80-90%. Adults feeding on hydrated food were observed to void water, likely to overcome problems with osmotic regulation. Given dry food with little water and high conversion rates (much greater than 10%) in larvae, evidence indicated D. maculatus obtains most water metabolically and not through diet. These adaptations indicate that D. maculatus (and likely related species) have evolved physiologically to exploit dry carrion thereby avoiding competition with other carrion-feeding species. However, D. maculatus is not adapted for low humidity environments or to avoid extreme water loss, these adaptations allow use of a unique resource but do not represent adaption to xeric conditions generally. Moreover, the very adaptations that allow D. maculatus to survive on dry food, are potentially lethal when only hydrated food is available.
Introduction
“Thousands have lived without love, not one without water.” ̶ [1].
Dermestid beetles are unique among decompositional insects because of their consumption of dry tissue. Most insects decomposing animals either feed on soft tissues (e.g., blow flies, flesh flies, filth flies, carrion beetles, cheese skippers, and coffin flies), are predators on decompositional insects (e.g., rove beetles, clown beetles, and wasps), or are drawn to animal carcasses because of an associated change in environmental conditions (often increased humidity) (e.g., springtails) or the need for salts (e.g., butterflies) [2].
The varied species in this family live in multiple habitats, provided there are reliable resources available [3]. Dermestids vary greatly in the length of their life cycles, with some species going from egg to adult in six weeks and others taking as long as a year or more to complete development [4]. Like all holometabolous insects, dermestids have four life stages (egg, larvae, pupa, and adult), with a variable number of larval stages but typically 5-6. While many beetle species have long generation times, after one year or longer, many dermestids can complete a generation in weeks to a couple months, which lets them better use temporary resources. However, like other beetles dermestids are relatively long lived (weeks to months) as compared to many other insects.
For most dermestids larva size ranges between less than 1 millimeter after hatching and up to 12mm pre-pupation. In the 15 to 20 days following hatching, larvae spend most of their time going through rapid growth and feeding. Under ideal circumstances this developmental stage spans between 18 and 20 days. Dermestids have different numbers of molts depending on species, and within species some may have variable numbers of larval molts (e.g., 5-9 based on rearing temperature in Dermestes alter DeGeer) [5], facultative diapause (dormancy) (in Trogoderma granarium Everts), and even a rare variation in metamorphosis called retrogression (also in Trogoderma granarium) arising from limitations in food availability, such as through competition [6]. Retrogression is a form of “reverse molting” in which the individual has reduced weight and size after molting and requires additional molts to return to a normal developmental sequence based on limitations in food availability, competition, and completion of other larvae during development.
Although dermestids have varying ecological roles, most in the Dermestes are generally associated with decomposition of dry animal tissues [7], with Dermestes maculatus DeGeer probably the most ubiquitous in North America. Because these beetles are typically found on dried carrion, it is generally assumed that their feeding preference is exclusively for dry tissue. Indeed, dermestids, including D. maculatus, are regarded as specialists on dry tissues in the context of the array of insects feeding on dead animals [8]. Additionally, Dermestes can survive long periods without food and through periods of near desiccation. However, when used for de-fleshing bones, dermestids feed on a combination of wet and dry tissue, and D. maculatus and related species are sometimes found on human bodies early in decompositional succession. Consequently, whether or not D. maculatus is adapted for feeding solely on dry tissue is unclear.
To address this question, a logical approach is to look for differences in development rates between D. maculatus reared on wet and dry food. The development of D. maculatus was examined by Zanetti, et al. [9] who reported rates by larval stage for 6 temperatures (15, 20, 22, 24, 27, 30). As described, Zanetti, et al. [9] reared larvae on “…beef with or without bone, boiled during 10 min to minimize colonization by fungi, mites, and other pathogens.” which presumably represents a hydrated food source. Thus, in addition to direct experimental comparison of development on hydrate and dry food, comparisons of new data with that of Zanetti et al. offer another avenue for evaluating the influence of dietary water on D. maculatus larval growth and survival.
Development is a dynamic process; many factors beyond food influence the speed and efficiency of development, including temperature, humidity, enzyme regulation, and photoperiod [10]. Further differences may be credited to the fact that oviposition in insects on a specific host is determined by various factors that may determine its suitability as a breeding medium, such as nutritional quality, host abundance [11], morphology, environmental conditions, age and size of individual and competition [12, 13]. Consequently, these factors can confound experiments to measure development, unless properly considered as part of the experimental design. Measurements in development can become difficult in a laboratory setting when these factors are not taken into consideration.
Besides providing a means for looking at differences in food use, consumption and conversion rates have not been previously examined for D. maculatus. Knowing these rates could be of use in forensic science for relating dermestid populations and time to tissue removal on decomposed bodies. So, we examined water content of food and its influence on D. maculatus development and survival.
Materials and Methods
Dermestes maculatus for experiments were obtained from colonies and maintained at the University of Nebraska- Lincoln (established in summer 2015) from a combination of purchased and feral beetles. Colonies were maintained at 16:8 L:D, 25°C, and fed dried pork (pig ears). For experiments, colony adults were placed on a diet consisting of 1 gram of wet liver and moistened sections of paper towel to stimulate oviposition. Eggs were laid on the moist paper towel, were transferred to a separate rearing container and maintained until reaching the third larval stage. For colonies and experiments we used DigiTherm® 38-liter Heating/Cooling Incubators which allowed for precision temperature regulation within 0.1°. These incubators have microprocessor-controlled temperature regulation, internal lighting, and a recirculating air system.
We conducted a series of preliminary experiments to establish appropriate larval densities, weight of food, and other experimental conditions before testing the influence of wet versus dry food. In efforts to both reduce error and address variations within stages of insects sampled, we initially examined consumption in first and second larval stages versus later stages and adults. Because the larvae in the first two stages eat so little food (ca. <0.001g dry weight) and this amount represent less than 5% of total consumption, we decided to conduct future experiments with 3rd stage larvae. This decision also reduced problems with survivorship, in that survival rates in the first two stages are highly variable.
We also tested various food sources. As colonies were maintained on pork, we decided early on to use some form of pork in our experiments. Pork liver has long been a substrate of choice in studies of development of carrion insects, however, in looking at wet vs. dry food, liver was unsuitable because of difficulty in establishing water content differences, problems in recovering larvae that had burrowed in the liver, and difficulties in excluding fungal and bacterial contamination in some instances. Other problems were separating frass and shed peritrophic membranes from remaining food. Fortunately, dried pig ears (used as dog treats) proved to be an ideal substrate for testing. We could easily re-hydrate pig ears to provide a workable substrate existing in wet and dry conditions. One potential problem was that in testing small amounts of substrate (specifically, less than 0.5 g) dried pig ears might absorb sufficient moisture from the air so as to confound our treatment and contribute to high variability in measured consumption. We solved this problem by increasing the number of larvae and amount of pig ear per experimental unit, so that the surface area to mass ratio of the pig ear was reduced and therefore absorbed water minimized.
This testing lead to the following observations relative to establishing our pig ear treats: (1) saturated pig ears, which were those soaked water at 25°C for 2 hours to achieve Food % survival Δ larval wt (g) food consumed (g) % conversion feeding rate/larva/day (mg) mean SE mean SE mean SE mean SE mean SE dry 79.5 2.7 0.051 0.019 -0.218 0.065 93.7 64.8 2.1 1 wet 23.2 5.5 0 0.003 -0.815 0.196 4.2 2.7 30 6 food water content: dry vs wet df 1, 25 1, 21 1,21 1, 14 1,21 F 56.27 6.58 12.34 21.35 30.53 P>F <0.0001 0.018 0.0009 0.0004 <0.0001 maximum absorbance (as evidence by constant weight) and a total weight typically double that of a dry pig ear; (2) wet pig ears, which are soaked at 25°C for ca. 30 min to achieve an increase in weight of about 50%; and (3) dry pig, which were ca. <10% moisture content. Preliminary tests showed that when beetles were fed saturated pig ears, the mortality rate in our testing was routinely 100%. To provide measurable consumption on hydrated tissue, we decided to use wet, rather than saturated pig ears for comparisons with dry ears. Additionally, in a natural setting where carrion starts losing fluids immediately after death, the wet pig ear are probably more representative of tissue beetles might natural encounter. One final consideration is the potential influence of intraspecific competition on the larval consumption and associated values. We decided to look at this explicitly by varying starting food amounts to represent different potential resource limitations for larvae.
The wet vs. dry food experiment was a randomized complete block with factorial treatment arrangement. Main effects were food water content (wet pig ears vs. dry pig ears) and food quantity (starting food weights of 6.0, 4.8, 3.6, 2.4, and 1.2 g) with 4 replications. The experimental unit was a 1.7 L plastic box with 10 new molted 3rd stage larvae on pig ear as specified for individual treatments. Experimental units were blocked inside growth chambers by shelves, and maintained at 25.0°C with a light: dark cycle of 12:12 and relative humidity (65±5 %), which are reported as optimal conditions for development [11]. Individuals were weighed at the beginning of the experiment and every 2 days thereafter. Final larval and consumption rates (used to calculate change in larval weight and food consumption) were based on values from the larval measurement immediately before pupation (Table 1). This procedure avoided errors in larval measurements estimated from initial pupal weights, in that pupation can cause a 10-20% reduction in weight [14].
food quantity: starting pig ear weight (6, 4.8, 3.6, 2.4, and 1.2g) for 10 larvae df 4, 25 4, 21 4, 21 1 ,14 4, 21 F 0.58 1.6 1.24 1.45 2.43
- P>F ns ns ns ns ns interaction of food water content vs. food quantity df
- 4, 25
- 4, 21
- 4, 21
- 1, 14
- 4, 21
- F
- 1.04
- 1.74
- 2.2
- 0.06
- 2.84
- P>F ns ns ns ns
- 0.0498
Table 1: Results from a factorial experiment with food water content (wet and dry pig ears) and food quantity (6, 4.8, 3.6, 2.4,
- conditions for this experiment were the same as previously described with the follow exceptions. Single larvae or adults were place on 0.5 g of dried pig ear, and only treatments with dry pig ears were used. N = 23 for larvae and n = 21 for adults
- (Table 2). larvae (3rd to pupa) adult time in stages (h) food consumption larva food consumption mean SE
- Δwt
- Δwt/day
- Δwt
- Δwt/day
- % conversion
- Δwt
- Δwt/day
- 548
- 0.092
- 0.0042
- 0.035
- 0.0015
- 44.50%
- 0.039
- 0.0077
- 26
- 0.011
- 0.0005
- 0.002
- 0.0001
- 4.60%
- 0.002
- 0.0004
Table 2: Weight gain, food consumption, and conversion percentage for larval (3rd to pupa) and adult food consumption of
Initial testing at all three levels provided enough detail to determine that saturated levels (generally unseen in nature outside of flood events or similar) were deemed excessive. We therefore excluded these treatments from further study and experiments.
Analysis
As previously stated, the goal of this study was to address and determine whether preferences or associations with moist and dry tissue are mitigated through resource availability, then further determine whether this influenced growth and development. For statistical analysis of the wet vs. dry experiment, we used SAS University Edition (https://www.sas.com/en_us/home.html), and mixed models procedures to accommodate missing points in the data set. Response variables were survivorship, change in larval weight, change in food weight, percent conversion, and feeding rates per day (Table 1). We also did linear regressions of proportion survivorship versus food weight for dry food and for wet food in Graph Pad Prism 8.01 (Graph Pad Software https://www.graphpad.com/).
Results and Discussion
We found a pronounced difference between how much D. maculatus (chiefly larvae) feed and their feeding rates on wet versus dry tissues. Our findings illustrate not only a preference for dry tissues, but a dependence on them. Indeed, hydrated tissue presents a potentially lethal challenge to D. maculatus. Adults have approximately 50% the daily consumption rates of larvae (averaged over 3rd to the last larval stage). Finally, conversion rate results indicate that D. maculatus must acquire moisture from non-dietary sources.
Survivorship
Table 1 and Figure 1 illustrate the influence of food moisture content on D. maculatus consumption rates and survival. As summarized in Table 1, wet and dry food treatments were significantly different for all variables measured. Although some larvae could complete development on moist tissue, high levels (ca. 75%) of mortality occurred on this food. In preliminary experiments with saturated tissues, 100% mortality occurred, and both larvae and adults were observed to “defecate” water droplets, presumably to maintain osmotic balance.
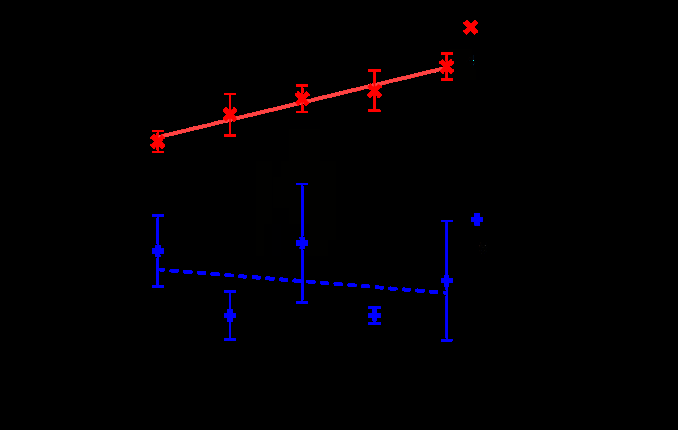
As compared to limitations in food quantity, we observed no differences in larval survivorship even with food per larvae as low as 0.12g, however, Figure 2 indicates that there was a linear reduction in survivorship with reduced dry food. In contrast, the linear regression for wet food quantity and survivorship was not significant, which is consistent with the significant interaction noted in our mixed models analysis (Table 1). One limitation in our experimental design is that wet treatments had less food content than dry treatments (because wet and dry treatments used the same weight and were not corrected for water content). Consequently, the wet treatment might have been more food limited than dry treatments. However, food limitation does not account for the low survivorship of wet treatments, in that the highest weight wet treatments still had dramatically lower survivorship than the lowest weight dry treatments (Figure 1). Moreover, our observations of purging water from the anus strongly indicates problems with excess water and osmoregulation.
Because they have large body surface relative to their volume, insects are susceptible to desiccation. While possessing little water in their bodies relative to size causes insects to dry out quickly. Desiccation tolerance in insects is accomplished through numerous physiological and behavioral adaptations including waxy epicuticles, furthered divisions in proto-cuticles and endo-cuticles, glycogen stowage, and eclosion during development. Water conservation is a common issue among all insects and is amplified among those associated with dry habitats. Typically, water conservation in digestion occurs through osmotic regulation in the Malpighian tubules of the hind gut, so insect waste (frass) is usually dry. However, insects processing fluid diets with excess water (such as true bugs and blood feeding insects) may defecate water to allow processing of sufficient food to meet nutritional demands (usually protein for plant feeding insects) and to maintain osmotic balance.
The surprising results and observations that D. maculatus regarding defecated water on hydrated food suggest that while D. maculatus has evolved to survive on an extremely dry diet, it has lost the ability to survive on normal, hydrated tissue.
Consumption and Conversion Rates
The same trend noted in the wet vs dry experiment is seen from consumption as compared to the amount (weight) of tissues consumed (Table 2). As a rule, only about ten percent of the energy of one trophic level can be transmitted to the next trophic level, the so-called rule of ten. Results of experiment 2 seem to contract this principle, because D. maculatus exceeded this ecological limit, averaging a conversion rate of over 44% (Table 2). The explanation for this result is, of course, that the calculated conversion rate is confounded with uptake of water in developing larvae. More specifically, these results indicate larvae are not obtaining the bulk of their water from their food. Consequently, calculating conversion rates only based on food weight over estimates the conversion rate.
Taken in conjunction with the survivorship findings of experiment one, the conversion data confirm that D. maculatus does not depend on dietary sources for water. Between wet and dry tissues there were noticeable differences in consumption and survivorship. When we looked at the dry tissues, we found that weight gain is disproportionately higher than would be explained by only the consumption of dry tissues consumed. Like all organisms, insects need water. Looking at the weight gains in our experiments, considering the lack of moisture in the dry tissues, and keeping in mind water requirements, these dermestids are clearly obtaining their water needs from some other source. Indeed, excess dietary water is potentially lethal. So where does the water come from? There are two possibilities: the source may be atmospheric water [15], or water produced metabolically, as observed in desert-adapted beetles.
Work by Fraenkel, et al. [14] showed that Tribolium confusum (Coleoptera: Tenebrionidae), Ephestia kuehniella (Lepidoptera: Pyralidae), and Dermestes vulpinus (Coleptera: Dermestidae) obtain water from a combination of atmospheric water and metabolic water, with metabolic contributions more important at low relative humidities. The same combination of sources almost certainly applies to D. maculatus. What has not been examined, to the best of our knowledge, is the impact of excess dietary water, as we did here in experiment one.
When we look at conversion rate it becomes clear that having to process extra water is unproductive. It is also inefficient when compared to biomass conversion. Moreover, larvae ate more hydrated food, presumably to obtain sufficient nutrients, but in the process exacerbated their problems with excess water. Because larval feeding behaviors did not adapt in response to the danger posed by hydrated food, we assume this is not a commonly experienced phenomenon. Thus, food choice seems likely to prevent mortality as long as sufficient acceptable, i.e. dried, food is available.
Why does excess water kill dermestids? The question itself fits well with one of the observations made during our study: beetles with hydrated food defecated water. As we noted previously, insects typically do not excrete water or liquid waste, and instances of insects defecating water are associated with osmoregulation. But osmoregulation comes at a significant energy cost, and when D. maculatus only have access to hydrated food, the more they eat to address energy needs for osmoregulation, the more they exacerbate their osmoregulatory problem. This interpretation is supported by data in Table 1: larvae feeding on wet food have much higher consumption rates (30 mg/larva/d on wet versus 2.1 mg/larva/d), but have a huge difference in larval weight gain ( <0.001 g/d on wet versus 0.051 g/d on dry).
In arid environments Zachariassen [16] points out that water conserving physiological adaptations among terrestrial insects, such as the beetles in our study, are accomplished through series of tradeoffs and compromises. Often, there is a substantial lethality due to dehydration hence a strong selection pressure for efficient physiological mechanisms to limit water loss in such species [16]. In addition to behavioral and circadian responses, there are a series of physiological responses which help insects deal with shortages of free water, extremes of temperature, and desiccation. Reduced cuticular water loss is associated with lowered metabolic rates and variances in oxygen consumption. Oxidative metabolic processes result in the formation of ATP and increases in sodium ion pumping. Most investigators agree that a very substantial part of cellular ATP turnover is spent in trans-membrane sodium pumping which in turn gives rise to a higher than usual electrochemical energy gradient [16]. These same costs in ATP expenditure and regeneration for osmoregulation will apply for excess hydration and removal of water to maintain osmotic balance.
The high energy costs associated with dehydration are avoided when water can be metabolically produced. Species using metabolic water as a primary water source presumably need access to adequate or even a surfeit of food. Larvae of T. confusum and E. kuehniella both meet this requirement, and the diet of Dermestes like D. maculatus and D. vulpinus includes calorie-rich proteins. However, the dermestids do not face the same challenges as desert-adapted species because they are not exposed to highly xeric environments. Ironically, although D. maculatus can thrive on an extremely dry diet, it is no better adapted to avoid desiccation than most insects. And the very adaptations that allow D. maculatus to survive on dry food, are potentially lethal when only hydrated food is available.
Conclusions
Rather than representing a physiological adaptation to occupy a specific biome, the physiological preference D. maculatus has evolved seem to show strong evidence of how they’ve niche specialized for a feeding guild, the carrion insects. When contrasted against virtually all other carrion insects, this adaptation for an exclusively dry diet illustrates how D. maculatus are able to avoid competition which is usually a challenge for other decomposers species. On decomposing soft tissues we see great competition among insects, whereas dermestids seem to capitalize on a resource inaccessible to other insects and thereby avoid conflict [17, 18].
References
-
Auden WH (1957) First things first. The New Yorker, USA, pp: 938.
-
Haskell NA, Williams NA (2008) Entomology and Death: A Procedural Guide. In: 2nd (Edn.), East Park Printing, Clemson, SC.
-
Kiselyova T, Mchugh J (2006) A phylogenetic study of Dermestidae (Coleoptera) based on larval morphology. Systematic Entomol 31: 469-507.
-
Triplehorn CA, Johnson NF (2020) Introduction to the Study of Insects. In: 7th (Ed.), Cengage Learning. Independence, KY.
-
Bujang NS, Kaufman PE (2010) Featured creatures: common name: black larder beetle, incinerator beetle scientific name: Dermestes ater DeGeer (Insecta: Coleoptera: Dermestidae). University of Florida Cooperative Extension Service, Pub. No. EENY-480.
-
Beck SD (1971) Growth and retrogression in larvae of Trogoderma glabrum (Coleoptera: Dermestidae). 1. Characteristics under feeding and starvation conditions. Annals Entomol Soc Amer 64: 149-155.
-
Parkinson AH (2012) Dermestes maculatus and Periplaneta americana: bone modification criteria and establishing their potential as climatic indicators. M.S. Dissertation, University of the Witwatersstrand, Johannesburg, South America.
-
Smith KS (1986) A Manual of Forensic Entomology. Cornell University Press, Ithaca, NY.
-
Zanetti NI, Visciarelli EC, Centeno ND (2015) The effect of temperature and laboratory rearing conditions on the development of Dermestes maculatus (Coleoptera: Dermestidae). J Forensic Sci 61(2): 375-3891.
-
Nabity PD, Higley LG, Heng-Moss TM (2007) Light- induced variability in the development of the forensically important blow fly, Phormia regina (Diptera: Calliphoridae). J Med Entomol 44(2): 351-358.
-
Usman Z, Ayertey JN, Cobblah MA (2013) Development of Dermestes maculatus (DeGeer, 1774) (Coleoptera, Dermestidae) on different fish substrates. Jordan J Biological Sci 6: 5-10.
-
Johnson CD, Kistler RA (1987) Nutritional ecology of Bruchid beetles. Nutritional ecology of insects, mite, spider and related invertebrates. John Wiley & Sons, New York, USA.
-
Siemens DH, Johnson CD, Woodman RL (1991) Determinants of host range in Bruchid beetles. Ecology 72: 1560-1566.
-
Fraenkel G, Blewett M (1944) The utilization of water in insects. Bull Entomol Res 34: 127-139.
-
Edney EB (1966) Absorption of water vapour from unsaturated air by Arenivaga sp. (Polyphagidae, Dictyoptera). Comparative Biochemistry and Physiology 19(2): 387-408.
-
Zachariassen KE (1996) The water conserving physiological compromise of desert insects. European J Entomol 93(3): 359-368.
-
Ahearn GA, Hadley NF (1969) The effects of temperature and humidity on water loss in two desert tenebrionid beetles, Eleodes armata and Cryptoglossa verrucosa. Comparative Biochemistry and Physiology 30: 739-749.
-
Byrd J, Castner J (2001) Forensic Entomology. The Utility of Arthropods in Legal Investigations. CRC Press, Boca Raton, FL.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind