Molecular Markers and Taxonomic Explosion in Herpetology: More or Less Robust Taxonomy? True and False Advantages of DNA Markers
Despite the increasing deficit of taxonomic expertise, the number of newly described species since the early 2010s has grown exponentially. This growth is related to the increased use of DNA markers in taxonomic descriptions. However, routine use of DNA markers in taxonomy did not bring practical taxonomy closer to the theory. Species are unique lineages with irreversible evolutionary pathways, and only the presence of distinct populations within the same geographic range, or at least the presence of narrow hybrid zones between the parapatric ranges is a conclusive evidence of evolutionary irreversibility. In the case of allopatric populations, only very high genetic distances, suggesting several tens of millions of years of independent evolution, can be used for validation of species status. This problem cannot be solved by the broader introduction of genomic phylogenies, which also fail to provide robust criteria for evolutionary irreversibility. We can hardly suppose that robust validation of species status is applicable to all or most of hundreds of thousands of animal species, including 20,000 amphibians and reptiles. Instead, practical taxonomy should concentrate on describing recognizable species, maintaining a trade-off between sufficiently detailed descriptions of world biodiversity and the applicability of these descriptions for practical use and metaanalyses, not pretending that formally described species reflect real lineages with independent and irreversible evolutionary pathways. Simultaneously, the non-critical elevation of the taxonomic status of individual geographic populations, contrary to the declared purpose of better-focusing conservation efforts, often has the opposite effect, leaving many formally described taxa outside the conservation umbrella.
Introduction
The invention of cheap PCR-based sequence technologies, established in the middle 1980s, triggered a revolution in taxonomy [1]. Discovery and description Conceptual Paper of new species since the time of Linnaeus and Pallas, were based on the individual experience of the naturalists. Long- lasting discussions were going in attempts to identify the most informative phenotypic characters that could help to separate “real” species from less meaningful traits that could vary among conspecific populations or morphs or make taxonomy more objective by introducing numerical analysis [2, 3, 4, 5, 6, 7, 8, 9]. By the XX century, a generation of taxonomists arose, specialized on smaller organismal groups, such as individual families of beetles or butterflies. It generally is considered that years of experience are required for a person to become a real expert on a certain group being able to make a reliable diagnosis of a species.
Generating molecular genetic data was supposed to make taxonomic practices more objective [10, 11, 12]. General biological knowledge is sufficient for seeing the DNA sequence differences between two phenotypically similar specimens. A zoologist working on a specific group of animals can easily convince a non-expert in his taxonomic suggestion, supporting his findings with some molecular genetic information. The new tool became widespread in taxonomic practices in the late 1990s; however, in original species descriptions of the 1990s and early 2000s, genetic data are rarely reported. In 2005, only in one [13] out of 31 descriptions of new lizard species from 67 most speciose genera the authors provided mitochondrial sequence information. It does not rule out that molecular data were used in the practical taxonomy of these years; however, this usually was done for already described species or elevation of taxonomic status of the earlier described subspecies [14, 15, 16, 17]. From this time, the inclusion of molecular data into species descriptions increased exponentially. In 2015, 65% of new lizard species descriptions referred to some molecular data (usually mitochondrial DNA sequences), and in 2023 this proportion increased to 98% [18].
Simultaneously, the number of annually described species increased sharply (Figure 1). In general, 50-100 species of lizards from 67 speciose genera were described within a period of five years since the middle XIX century until the end of the XX century; since the early 2000s, this number increased five times.
![Figure 1: The number of new species of lizards described per five year periods, since middle XVIII century to the present day. The diagram is based on the analysis of 67 lizard genera that include 20 or more described species, according to Uetz, et al. [18]. Only newly described taxa are considered; infraspecific taxa with elevated status or re-described species are ignored.](/fulltextimages/12265/fig_1.png)
Figure 1: The number of new species of lizards described per five year periods, since middle XVIII century to the present day. The diagram is based on the analysis of 67 lizard genera that include 20 or more described species, according to Uetz, et al. [18]. Only newly described taxa are considered; infraspecific taxa with elevated status or re-described species are ignored.
This taxonomic explosion may look surprising because it happened concurrently with the general decline of taxonomic expertise [19, 20, 21]. This fact suggests that the wide use of DNA sequencing was likely decisive for the fast growth of the nominal species’ number. The development of Barcoding of Life projects [22, 23, 24, 25, 26] was an additional factor for the taxonomic explosion. These projects accelerated efforts towards the documentation of biodiversity, and revealed the presence of a “barcoding gap”, i.e. some level of mitochondrial DNA divergence, which is in line with species boundaries described by experienced taxonomists [27].
Limitations of Mitochondrial DNA: Positive and Negative Sides
In a living cell, mitochondrial genes produce thousands of copies, different from most nuclear genes [28, 29]. This makes mitochondrial DNA (mtDNA) fragments easier to amplify than any other parts of the genome [30, 31]. As a result, mtDNA sequencing has become a routine method for phylogenetic and taxonomic studies since the late 1980s. In Figure 2, the growth of the amphibian and reptile studies using mtDNA analysis is shown between 1980 and 2023.
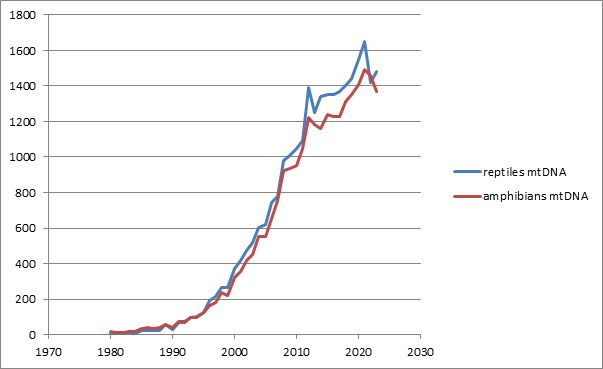
Mitochondrial gene sequences contain important information on species evolution. Mitochondrial phylogenies help to reconstruct patterns of geographic expansion of a taxon. Projecting phylogenies on geographic maps became a specific field of science called phylogeography [32, 33, 34, 35, 36]. Comparing mtDNA sequences helps to infer the geological time when the ancestral lineages of two populations started to diverge. The molecular clock hypothesis assumes that substitutions in individual genes are aggregated with a predictable speed [37, 38]. Molecular clocks can be calibrated by known and well-dated geological events (such as the separation of two islands by sea) [39, 40], or by comparing DNA sequences of current specimens with fossil DNA [41, 42]. Molecular clock hypothesis was repeatedly criticized, because fixation rates of new substitutions may depend on the population size, and can substantially differ among taxa [43, 44, 45, 46], but proper statistical analysis helps to estimate reasonable confidence limits [47, 48, 49, 50, 51].
Simultaneously, the use of mtDNA sequences for the analysis of species boundaries is problematic. One challenge is incomplete lineage sorting [52]. Each of the two populations descending from a common ancestor may be not genetically homogenous, and fixation of alleles at polymorphic loci in diverging populations can occur at different speeds; both the same or different alleles can be fixed, depending on a locus [47, 53, 54]. As a result, phylogenetic trees based on unlinked genes may not coincide, causing an inconsistency between gene evolution and species evolution [52, 55, 56, 57, 58, 59]. Different authors suggested combining sequences of mitochondrial and nuclear genes to infer more reliable phylogenies [60, 61, 62]. Another problem is a common horizontal transfer of mitochondrial genes among closely related species [63, 64, 65, 66, 67, 68, 69, 70]. Probably due to these problems, and the broader invention of genomic technologies, since the early 2010s, the growth rate of the publications presenting mt-DNA data in herpetological studies declined (Figure 2).
Attempts to Overtake Incomplete Lineage Sorting Problem: Right and Wrong Approaches
Since late 1990s, evolutionary biologists and taxonomists invented a routine complementing mitochondrial sequences with sequences of one or more nuclear genes, usually those which produce multiple copies in a genome, such as genes C-mos, RAG-1, BDNF in amphibians and reptiles [71, 72, 73, 74, 75]. Coding nuclear genes are far more conservative than mitochondrial DNA, probably because of continually acting stabilizing selection [76]. Nuclear gene sequences usually have too few informative positions for producing reliable, statistically well-supported phylogenetic patterns if closely related species are studied. Some authors use a concatenation of nuclear and mitochondrial genes, trying to increase the robustness of the generated phylogenies [77, 78, 79, 80, 81]. This is a conceptually incorrect approach. Concatenated sequences that include mitochondrial genes with a substantial number of informative positions and nuclear genes with just a few informative sites usually produce a tree simply coinciding with mitochondrial phylogeny. Meanwhile, the very sense of the analysis of incomplete lineage sorting is validation of a null hypothesis suggesting coinciding phylogenies, based on the unlinked loci analyzed separately. Other taxonomic papers provide separate phylogenies or haplotype networks based on individual nuclear genes: in this way, incomplete lineage sorting can indeed be detected [82, 83, 84, 85, 86, 87].
Species Concepts: from Dobzhansky and Mayr to the Present Day
The taxonomic explosion of the late 1980s occurred along with the resuming of the discussion about species concepts, which never ended since the publication of Charles Darwin’s “The Origin of Species” [88]. A very logical concept of polytypic biological species to which the majority of scientists appealed for several decades, has an important weakness: it cannot be practically applied to thousands of described taxa [89, 90]. Validation of pre- or post-zygotic isolation, a central issue of this concept, requires extensive analysis, which does not apply to most newly described species because of a lack of funds, time, and expertise. In fact, most taxonomists relied on a traditional taxonomic approach based on more or less subjective expert opinions [91]. However, the lack of a firm scientific theory behind the taxonomic practices challenges taxonomic conclusions. For this reason, concepts that were easier to validate, such as the phylogenetic species concept [92, 93, 94], replaced the biological species concept. The phylogenetic species concept considers monophyletic origin as a sufficient precondition for considering a population to be a separate species. The concept of genealogical concordance, based on the idea of incomplete lineage sorting is, in fact, an extended definition of phylogenetic species, because it refers to the ancestral evolutionary pathway and not to the current metapopulation structure of a taxon.
Some other species concepts have become popular in recent decades. The relatively old concept of Evolutionary species does not require reproductive isolation as a necessary precondition of species reality [95, 96, 97]. The unified species concept of DeQueiroz, defining species as a separately evolving lineage, accepts ways of validating species status [98, 99]. The departure from the biological species concept was largely a result of understanding that introgressive hybridization does not necessarily cause a merger of the evolutionary lineages and may occur concurrently with the ongoing divergent evolution [100, 101].
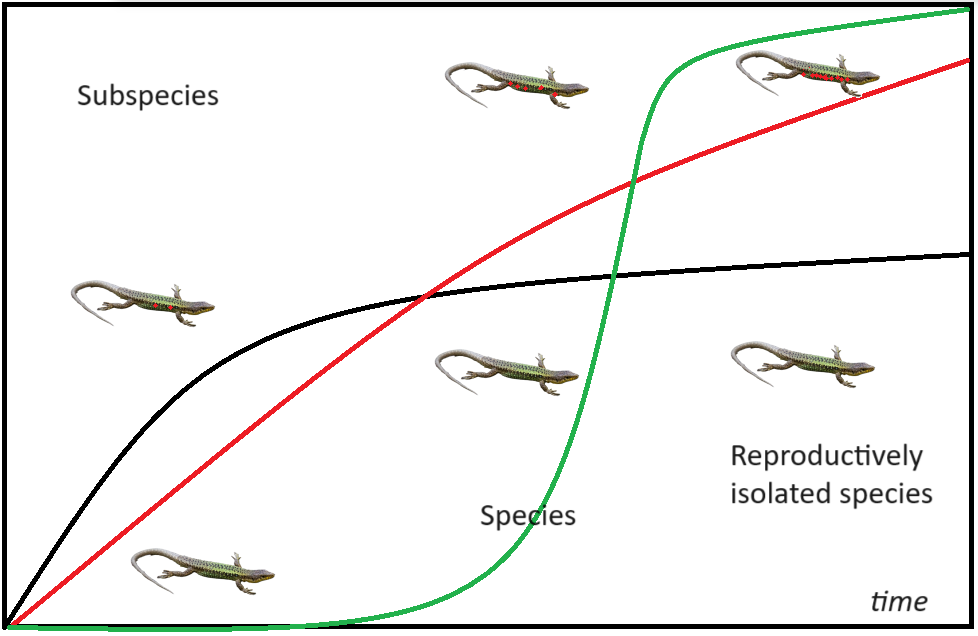
DeQueiroz did not thoroughly explain what “separately evolving lineage” means. Simultaneously, one of the central ideas of the theory of evolution and speciation is the irreversibility of evolutionary changes. In general, the evolutionary pathway is considered to be irreversible [102, 103, 104], although this hardly can be applied to ephemeral lineages that occasionally re-merge in case of secondary contact. I would suggest that the evolutionary pathway of a species cannot be reversed, and each species is unique: it can evolve further, split into two daughter species, or extinct, but not lose its distinctness from any other species. The irreversibility criteria is concurrent with the concept of evolutionary species and separately evolving lineage of DeQueiroz, which don’t rule out potential hybridization, but consider the presence of stable, species-specific complexes of genotypic and phenotypic characters.
Multiple genetic studies conducted in recent decades showed that hybridization and gene flow between diverging incipient species is commonplace [105, 106]. Why could this happen? Fixation of differential de novo mutations in two isolated populations, as a result of gene drift and selection, decreases the number of the shared alleles. Interbreeding of individuals with different alleles at the same locus increases heterozygosity, reduces the probability of inbreeding depression, and, consequently, can increase the fitness of the hybrid offspring. Multiple examples suggest superior fitness in hybrids of even formally described different species [107, 108, 109]. However, in some cases, heterozygotes have inferior fitness [110, 111, 112, 113, 114]. Moreover, over time, the differential mutations are aggregated that change genomic architecture, and cause dysfunction of genes in hybrids, or incompatibility between nuclear and organellar genomes [115]. Inferior fitness of heterozygotes can also be a result of adaptation of the isolated populations to different environmental conditions [116, 117]. These deleterious effects slowly aggregate, and above a certain threshold their effect overweighs the positive effect of heterosis (Figure 3). After this time point, the speciation process becomes irreversible.
Similar to the biological species concept, the irreversibility of speciation (i.e., presence of separate evolutionary pathways) can hardly apply to practical taxonomy. The inferior fitness of hybrids is hard to detect without exhaustive study on the population level. It is impossible to conduct such studies for hundreds of annually described nominal species; therefore, this criterion, albeit theoretically very convincing, can rarely be used as an effective tool. This builds a gap between evolutionary theory and formal taxonomy. Probably, it is counterproductive to apply strict species criteria for describing new species, but even more harmful is an incorrect application of theory. The monophyletic origin does not prove that populations, even with diagnostic phenotypic features, cannot merge in case of secondary contact. Neither the presence of genealogical concordance, nor geographic isolation can be taken as an evidence of irreversible speciation process. The level of differences sufficient for the development of full reproductive isolation varies among animal taxa. It could be taken as an indirect argument of species-level differences and not a conclusive evidence.
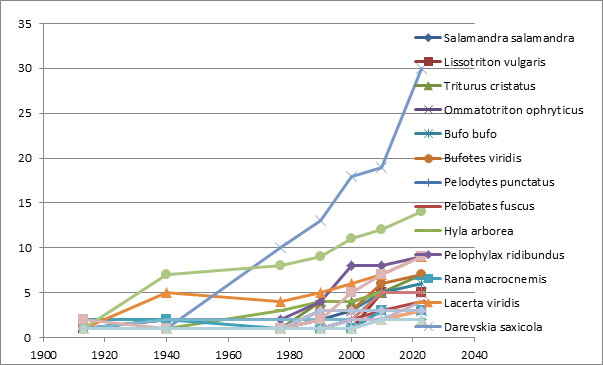
Can we give priority to one or another of these concepts? Irrespective of a theory, scientists need to name living beings for inventing terms that help to understand what we are speaking about. Scientific names, such as Bufo bufo, Lacerta agilis, or Natrix natrix save us time: we don’t need to write long sentences explaining what kind of animal we are speaking about, whether or not it has a tail and legs and if its body is covered with scales, or how it is colored, replacing long unnecessary explanations by a Latin binomial invented by Linnaeus. Hence, practical taxonomy, first of all, should pursue the nomenclature well describing diagnostic features of a group of organisms, and last but not least, easy to remember by ecologists and evolutionary biologists. Those purposes are rarely considered in recent taxonomic practices. Scientists describe thousands of “cryptic” species with no diagnostic phenotypic characters, based only on the differences in the sequence of a few genes [118]. Based on some sequence differences, they often split “traditional” species into several new ones which are difficult to remember by a non-expert in a specific taxonomic group. How this process of splitting is intensified in recent decades, in the example of the West Eurasian herpetological fauna is shown in Figure 4. In total, 25 species from 22 genera of amphibians and reptiles were split into 47 species by 1977, 86 species before 2000, and into 140 species by 2023.
Introgressive Hybridization Challenges Determination of Species’ Boundaries
Although introgressive hybridization may not prevent continuing divergence of incipient species [100], it has a certain impact on the evolution within each of the hybridizing lineages. Some genes may be horizontally transmitted from one lineage to another. Typical cases of DNA transmissions among the species, sometimes interpreted as “reticulate speciation”, is transfer of mitochondrial DNA among the species. In some cases, such as frogs Rana chensinensis and R. kukunoris [66], toads Bufo fowleri and Bufo americanus [63], horned lizards of genus Phrynosoma [65], skinks of genus Messalina [86], Alligator lizards from the genus Elgaria, or rock lizards Darevskia obscura and Darevskia portschinskii, in spite of a distinct phenotypic and allelic boundary between the species, mitochondrial DNA of a related species replaced original mitochondrial lineage in the peripheral part of their ranges. In the Carpathian newt (Lissotriton montandoni) the original mitochondrial lineage is completely replaced by the mitochondrial DNA of a more widespread smooth newt (Lissotriton vulgaris) [119, 120]. The same is true for leaf-tailed geckos of the genus Uroplatus, or some rock lizards from the genus Darevskia. Genetic studies showed discordance between mitochondrial and nuclear phylogeny of at least four very distinct and reproductively isolated species of this genus: Darevskia alpina, D. derjugini, D. praticola, and D. parvula. In the first two species, mitochondrial genes are closer to that of lizards with whom their geographic range overlap than with allopatric species to which they are closer phenotypically and by nuclear genes. In general, the discordance of nuclear and mitochondrial phylogenies is not an exception for natural genetic systems.
Actually, we don’t know how common the introgressive pattern of mitochondrial DNA in different organismal groups is. Unfortunately, combined analysis of mitochondrial and nuclear genes does not always aim to infer patterns of introgression. As a consequence, it remains a formal procedure pursuing the search of some theoretical ground for nominating species.
Practical and Conservation Outcomes: Taxonomic Inflation
Some authors defined the accelerated description of new species in the first quarter of the XXI century as “taxonomic inflation”. The very idea of this definition is that unlimited descriptions of new species, applying to highly variable arguments and datasets, result in the devaluation of a species as a part of nature deserving attention and conservation. Some negative consequences of taxonomic inflation are discussed by previous authors [121]. Here I would underline the three most important ones from my point of view.
(1) Complication of the task for decision-makers. Governmental bodies, which are in charge of environmental protection, must identify conservation priorities for effectively spending taxpayer’s money. A routine part of this procedure is identifying species that need more urgent conservation actions than the others. In national legislation, there are no definitions for upper taxa or “species groups”, and for this reason, the officials require scientists to provide them with scientific names of prioritized species or, at least, subspecies. In private conversations, taxonomists commonly suggest that describing new species is useful for increasing attention to taxa, especially local endemics, which need to be protected. I think that the actual effect is the opposite. Including five taxa in lists of protected species (such as national Red Lists) would not increase the funds that a government will spend for their conservation. Hundreds and thousands of nominal species suggested for conservation actions just frighten decision- makers who often have superficial knowledge of taxonomy. Further, splitting a species, formally protected by law, into several ones often causes a mess, and this “old” species remains protected by national legislation of international agreements only in a part of the range where the nominal form is found. Lizard Lacerta trilineata, snakes Dolichophis jugularis, Elaphe quatuorlineata, Vipera ammodytes, Vipera ursini, amphibians Bufo viridis and Hyla arborea are included under Annex II (species requiring special protection) of Bern convention on conservation of Wildlife and Natural Habitats (https://www.coe.int/en/web/bern-convention/home ). Simultaneously, subspecies of all these species from the Caucasus biodiversity hotspot were re-described as separate species in recent decades [122, 123, 124, 125, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151, 152]. None of these “new” species is prioritized in conservation programs supported by the European Commission.
(2)Nominal species commonly interbreed with their close relatives in the contact areas. Gene flow among the populations (either conspecific or formally non-conspecific) is a natural process affecting genetic diversity, exchange of adaptive alleles, and the effective population size. Typically, hybridization is not desirable because it may result in inferior fitness of hybrids. But simultaneously, common introgression of genes among populations may indicate the presence of positive effects of natural hybridization, including transfer of adaptive genes and increasing an effective population size, if gene flow is sufficiently high. Protection of a single nominal taxon and leaving unprotected adjacent populations of even formally different species may substantially reduce the effective size of its population and increase its vulnerability.
(3)For the understanding of the structure and dynamics of global biodiversity, it is important to develop integrated analyses of faunas and floras for large regions of the World, including biogeographic realms. Such analyses preferably cover evolutionarily distant taxonomic groups. Growth
of the number of formally described allopatric species misleads scientists conducting such an analysis, causing overestimation of differences and hiding similarities. For instance, herpetological fauna of central and Mediterranean parts of Europe has several widespread species, such as Lissotriton vulgaris, Triturus cristatus, Pelophylax ridibundus, Hyla arborea, Bufotes viridis, Bufo bufo, Lacerta trilieata, Anguis fragilis, Vipera ursini, Vipera ammodytes, Elaphe quatuorlineata, or Malpolon monspessulanus. All these species were assumed to exist in the Caucasus biodiversity hotspot before the status of Caucasian regional populations or subspecies was elevated to species [127]. As a result of taxonomic splitting, the region has now the species Triturus karelinii, Pelophylax bedriagae, Hyla orientalis, Bufotes variabilis, Bufo verrucosissimus, Anguis colchica, Vipera eriwanensis, Vipera transcaucasiana, Elaphe urartica, Malpolon insignitus. Most of these species are hardly recognizable from their Mediterranean relatives without a detailed study of their morphology or comparison of their DNA markers, and their ecosystem function hardly differs from that of their closest relatives. However, the nomenclatural differences give an impression of higher differences between the Caucasian and European herpetological faunas than in fact, it has. It also inflates the importance of real relict endemics, such as Caucasian salamander (Mertensiella caucasica) or Caucasian parsley frog (Pelodytes caucasicus), whose separate evolution counts tens of millions of years and who developed unique, highly specific phenotypes and ecological niches [127, 128, 129].
Do DNA Markers Make Taxonomy Less Biased?
As said, there is hardly a universal algorithm that could firmly validate species status (i.e., that of an evolutionary lineage whose evolutionary pathway became irreversible). Neither morphological study nor the analysis of mitochondrial DNA or even genomic phylogeny can solve this question without deep analysis, including hybridization experiments or a comparative study of viability for hybrids and purebreds. Such kind of analysis cannot cover hundreds of thousands of described species, including about 20,000 amphibians and reptiles. Practical taxonomists and evolutionary biologists commonly use any argument, based on the DNA marker analysis, to support their nomenclatural suggestions. We could classify these arguments: (1) Use of genetic distance based on the mitochondrial sequence analysis. This approach may be used as an indirect argument for treating an OTU (operative taxonomic unit) as a separate species, assuming that the molecular clock hypothesis works and it is likely that the number of genomic architectural mutations over time inevitably cause reproductive incompatibility. However, this time may substantially vary among the taxa. Besides, in some cases, pre-zygotic isolation may develop before post- zygotic [130]. (2) Mitochondrial (or nuclear) phylogeny as an argument for attaining species status to geographically separated populations. This argument is commonly used along with a morphological description of a taxon. However, monophyly is a weak argument if not supported by accounted genetic differences. Many local populations, at some stage after their establishment, are monophyletic groups of organisms. They often share common phenotypic features, which is not evident for the irreversibility of their evolution. Non-critical use of phylogenies for species description may cause extreme taxonomic inflation in the case of “nomenclatural harvesting” [131]. (3) Genealogical concordance, i.e., concurrent phylogeny based on unlinked genetic character. It is supposed that the lineages that achieved this stage of differentiation are isolated long enough to achieve an independent evolutionary pathway [132]. However, if the effective size of an isolated population is small, genealogical concordance can be reached in a short time, which may be insufficient for the development of pre- or post-zygotic isolation or irreversibility of evolution. Moreover, in most cases, scientists rely on combined analysis of mitochondrial and one or two nuclear genes, which usually have insufficient signals for building a robust phylogeny. Besides, concordant phylogenies based on very few loci are insufficient for driving conclusions about genealogical concordance throughout the entire genome. (4) The most expensive and informative approaches are those based on the high-throughput sequences of genomes, such as RAD sequencing [133, 134]. These approaches help to build more reliable phylogenetic trees and group species into genetic units using Bayesian inference [135]. However, even in this case, one cannot rule out the effect of a small effective population size, when genetic specificity is achieved in a small period of divergent evolution, insufficient for the development of effective reproductive isolation.
Generally speaking, none of the broadly used methods based on genetic or morphological differences between OTUs can be taken as conclusive evidence for separate evolutionary pathways, i.e. considering a population to be a separate species. The only safe approach remains one defined in early papers of Mayr, et al. [136, 137] the presence of two distinct and non-merging taxa within the same geographic range. An additional group of evidence is the presence of a narrow hybrid zone when two species come into contact but remain distinct throughout most of their range, such as in a classical example of fire-bellied toads [138, 139]. In the case of geographically isolated populations talking about the presence of different species can only be safe if the lineages are isolated obviously for tens of millions of years and show a very strong phenotypic hiatus – such as in the case of sister species of relict salamanders [140], or parsley frogs.
In all other cases nominating geographically isolated, closely related OTUs cannot be considered an objective way of describing species diversity if we agree that species are lineages with separate and irreversible evolutionary pathways. But what should we do with the myriads of binomials, which scientists introduced for the last two and a half centuries? Here I would return to “practically useful” taxonomy: it should help scientists, not only, to understand what are we speaking about when using a particular scientific name. In other words, it should keep a balance between a sufficiently detailed description of the observed biological diversity and the ability of a person to memorize the meaning of words.
Both describing more and more species and renaming earlier described taxa (both species and higher order taxonomic groups) do not help to memorize species names. For instance, 30 years ago, most Colubrid snakes from the Caucasus Ecoregion were aggregated into two genera (Coluber, Elaphe). Currently, species from the clade to which the nominal genus Coluber belongs are distributed among ten genera, and only two of them, Macroprotodon and Eirenis, were not members of this genus in early taxonomic systems. The inclusion of these two genera into Coluber would be a more parsimonious taxonomic solution, although taxonomists preferred splitting and turning three former genera into ten new ones.
In some recent papers [141, 142] broader use of the concept of geographic subspecies is suggested for allopatric, genetically, and morphologically close population groups. Nominating subspecies may be the way to a detailed description of biodiversity without the infinite complication of biological nomenclature. Understanding diversity within polytypic species is important for experts working on specific taxa, and retaining more inclusive species names would help to understand diversity for the general public, conservation bureaucrats and activists, and scientists analyzing biogeographic patterns throughout large geographic areas. Nominating subspecies does not meet conceptual difficulties that meet species descriptions. The criterion for describing separate subspecies is clear and simple: the ranges of different subspecies cannot coincide or overlap. If they are allopatric, the subspecies can be described based on even little genetic differences or the presence of private (not necessarily diagnostic) phenotypic characters. If the ranges are parapatric, i.e., conspecific individuals may move across the geographic boundaries between populations, the subspecies can still be described based on distinct diagnostic characters but no evidence for the formation of a tension hybrid zone [143, 144, 145] or otherwise evidence of selection against the subspecies hybrids. In this case, populations with intermediate characters or polymorphic by diagnostic characters do not belong to any subspecies, and their species name can be used without indication of a subspecies.
Conclusion
Use of Genetic Markers for Better Understanding Biodiversity
Molecular markers are indispensable for various kinds of biodiversity research, potentially including identification of species boundaries, but only rarely are they correcly used for the identification of these boundaries. It is correct to identify species based on the DNA marker analysis of individuals from the same range or contiguous ranges, but in the case of allopatric taxa, mtDNA sequences and even genomic data-based phylogenies could only be used as an indirect argument, in case if the differences are excessive. In all other cases, the description of a new species is still based on the experience and preferences of a taxonomic expert, and molecular markers are used for supporting a pre-defined suggestion and not for objective and unbiased validation of a hypothesis. The appearance of new species names in checklists does not depict “true” species diversity; at least if we assume that species is a real evolutionary lineage that attained an irreversible stage of divergence from its closest relatives (and hence has an independent evolutionary pathway). For practical reasons, however, the description of new species does not, and probably should not pursue strict evolutionary concepts. Instead, it can be accepted that, in most cases, formally named species are more or less subjective entities, helping to better understand the existing diversity. In this latter case, however, taxonomists should not oversplit the existing taxa, remembering potential problems associated with taxonomic inflation. Molecular markers, meanwhile, remain a powerful tool for understanding phylogeographic patterns, gene flow rates among the populations, or speciation process [146, 147, 148, 149, 150].
Acknowledgement
The paper was prepared while the author was in acceptance of grant by Shota Rustaveli National Science Foundation, project code FR-23-17324. The author appreciates students of Ecology at Ilia State University and Oleksiy Yanchukov for useful discussions.
References
-
Mullis K, Faloona F, Scharf S, Saiki R, Horn G, Erlich H (1986) Specific enzymatic amplification of DNA in vitro: the polymerase chain reaction. In Cold Spring Harbor symposia on quantitative biology. Cold Spring Harbor Laboratory Press 51: 263-273.
-
Simpson GG (1951) The species concept. Evolution 5(4): 285-298.
-
Hennig W (1966) Phylogenetic Systematics. University of Illinois Press, Urbana, Illinois.
-
Assis LC, Rieppel O (2011) Are monophyly and synapomorphy the same or different? Revisiting the role of morphology in phylogenetics. Cladistics 27(1): 94- 102.
-
Sokal RR (1966) Numerical taxonomy. Scientific American 215(6): 106-117.
-
Sokal RR (1986) Phenetic taxonomy: theory and methods. Ann Rev Ecol Syst 17(1): 423-442.
-
McNeill J (1974) The handling of character variation in numerical taxonomy. Taxon 23(5-6): 699-705.
-
Stevens PF (1991) Character states, morphological variation, and phylogenetic analysis: a review. Systematic Botany 16(3): 553-583.
-
Guisande C, Vari RP, Heine J, Garcia-Rosello E, Gonzalez- Dacosta J, et al. (2016) VARSEDIG: an algorithm for morphometric characters selection and statistical validation in morphological taxonomy. Zootaxa 4162(3): 571-580.
-
Lee MS (2004) The molecularisation of taxonomy. Invertebrate Systematics 18(1): 1-6.
-
Hillis DM (2019) Species delimitation in herpetology. J Herpetol 53(1): 3-12.
-
Dufresnes C, Poyarkov N, Jablonski D (2023) Acknowledging more biodiversity without more species. Proc Natl Acad Sci 120(40): e2302424120.
-
Günther R, Whiting A, Bauer A (2005) Description of a new species of African skink of the genus Trachylepis. Herpetozoa 18: 11-24.
-
Joger U, Lenk P, Baran İ, Böhme W, Ziegler T, et al. (1995) The phylogenetic position of Vipera barani and V. nikolskii within the Vipera berus complex. Herpetologia Bonnensis pp: 185-194.
-
Arntzen JW, Wallis GP (1999) Geographic variation and taxonomy of crested newts (Triturus cristatus superspecies): morphological and mitochondrial DNA data. Contributions to Zoology 68(3): 181-203.
-
Gvoždík V, Jandzik D, Lymberakis P, Jablonski D, Moravec J (2010a) Slow worm, Anguis fragilis (Reptilia: Anguidae) as a species complex: genetic structure reveals deep divergences. Mol Phylogenet Evol 55(2): 460-472.
-
Garcia-Porta J, Litvinchuk SN, Crochet PA, Romano A, Geniez PH, et al. (2012) Molecular phylogenetics and historical biogeography of the west-palearctic common toads (Bufo bufo species complex) Mol Phylogenet Evol 63(1): 113-130.
-
Uetz P, Freed P, Aguilar R, Reyes F, Kudera J, Hošek J (2023) The Reptile Database.
-
Hopkins GW, Freckleton RP (2002) Declines in the numbers of amateur and professional taxonomists: implications for conservation. In Animal conservation forum. Cambridge University Press 5(3): 245-249.
-
Krell FT (2004) Parataxonomy vs. taxonomy in biodiversity studies–pitfalls and applicability of ‘morphospecies’ sorting. Biodiversity & Conservation 13: 795-812.
-
Agnarsson I, Kuntner M (2007) Taxonomy in a changing world: seeking solutions for a science in crisis. Syst Biol 56(3): 531-539.
-
Smith MA, Poyarkov NA, Hebert PD (2008) DNA barcoding: CO1 DNA barcoding amphibians: take the chance, meet the challenge. Mol Ecol Res 8(2): 235-246.
-
Stoeckle MY, Hebert PD (2008) Barcode of life. Scientific American 299(4): 82-89.
-
Costa FO, Carvalho GR (2010) New insights into molecular evolution: prospects from the Barcode of Life Initiative (BOLI) Theory in Biosciences 129: 149-157.
-
Adamowicz SJ (2015) International Barcode of Life: Evolution of a global research community. Genome 58(5): 151-162.
-
Geiger MF, Astrin JJ, Borsch T, Burkhardt U, Grobe P, et al. (2016) How to tackle the molecular species inventory for an industrialized nation-lessons from the first phase of the German Barcode of Life initiative GBOL (2012– 2015) Genome 59(9): 661-670.
-
Čandek K, Kuntner M (2015) DNA barcoding gap: reliable species identification over morphological and geographical scales. Mol Ecol Res 15(2): 268-277.
-
Robin ED, Wong R (1988) Mitochondrial DNA molecules and virtual number of mitochondria per cell in mammalian cells. J Cell Physiol 136(3): 507-513.
-
Desalle R, Schierwater B, Hadrys H (2017) MtDNA: The small workhorse of evolutionary studies. Front. Biosci (Landmark Ed.) 22(5): 873-887.
-
Kocher TD, Thomas WK, Meyer A, Edwards SV, Paabo S, et al. (1989) Dynamics of mitochondrial DNA evolution in animals: amplification and sequencing with conserved primers. Proc Natl Acad Sci USA 86: 6196-6200.
-
Avise JC (2004) Molecular markers, natural history, and evolution, 2nd ed. Sinauer Associates, inc, Sunderland, Massachusetts pp: 401-466.
-
Avise JC, Arnold J, Ball RM, Bermingham E, Lamb T, et al. (1987) Intraspecific phylogeography: the mitochondrial DNA bridge between population genetics and systematics. Ann Rev Ecol Syst 18(1): 489-522.
-
Avise JC (1989) Gene trees and organismal histories: a phylo- genetic approach to population biology. Evolution 43:1192-1208.
-
Emerson BC, Hewitt GM (2005) Phylogeography. Current Biol 15(10): R367-R371.
-
Avise JC (2009) Phylogeography: retrospect and prospect. J Biogeogr 36(1): 3-15.
-
Poe S, Nieto-Montes de Oca A, Torres-Carvajal O, De Queiroz K, Velasco JA, et al. (2017) A phylogenetic, biogeographic, and taxonomic study of all extant species of Anolis (Squamata; Iguanidae) Syst Biol 66(5): 663- 697.
-
Thorpe JP (1982) The molecular clock hypothesis: biochemical evolution, genetic differentiation and systematics. Ann Rev Ecol Syst 13(1): 139-168.
-
Bromham L, Penny D (2003) The modern molecular clock. Nat Rev Genetics 4(3): 216-224.
-
Donoghue PC, Benton MJ (2007) Rocks and clocks: calibrating the Tree of Life using fossils and molecules. TREE 22(8): 424-431.
-
Loeza-Quintana T, Adamowicz SJ (2018) Iterative calibration: a novel approach for calibrating the molecular clock using complex geological events. J Mol Evol 86: 118-137.
-
Near TJ, Sanderson M J (2004) Assessing the quality of molecular divergence time estimates by fossil calibrations and fossil–based model selection. Phyl Transact Royal Soc London. Series B: Biological Sciences 359(1450): 1477-1483.
-
Müller J, Reisz RR (2005) Four well‐constrained calibration points from the vertebrate fossil record for molecular clock estimates. BioEssays 27(10): 1069- 1075.
-
Heads M (2005) Dating nodes on molecular phylogenies: a critique of molecular biogeography. Cladistics 21(1): 62-78.
-
Schwartz JH, Maresca B (2006) Do molecular clocks run at all? A critique of molecular systematics. Biol Theory 1: 357-371.
-
Pulquerio MJ, Nichols RA (2007) Dates from the molecular clock: how wrong can we be? TREE 22(4): 180-184.
-
Dietrich MR, Skipper RAJr (2007) Manipulating underdetermination in scientific controversy: the case of the molecular clock. Perspectives on Science 15(3): 295-326.
-
Ho SY, Duchêne S (2014) Molecular‐clock methods for estimating evolutionary rates and timescales. Mol Ecol 23(24): 5947-5965.
-
Tao Q, Tamura K, Mello B, Kumar S (2020) Reliable confidence intervals for RelTime estimates of evolutionary divergence times. Mol Biol Evol 37(1): 280- 290.
-
Tiley GP, Poelstra JW, Dos Reis M, Yang Z, Yoder AD (2020) Molecular clocks without rocks: new solutions for old problems. Trends in Genetics 36(11): 845-856.
-
Paradis E, Claramunt S, Brown J, Schliep K (2023) Confidence intervals in molecular dating by maximum likelihood. Mol Phylogenet Evol 178: 107652.
-
Kumar S, Suleski M, Craig JM, Kasprowicz AE, Sanderford M, et al. (2022) TimeTree 5: an expanded resource for species divergence times. Mol Biol Evol 39(8): 174.
-
Avise JC, Ball RMJr (1990) Principles of genealogical concordance in species concepts and biological taxonomy. in D. Futuyma and J. Antonovics, eds. Oxford Surveys in Evolutionary Biology. Oxford University Press, New York, USA, pp: 45-67.
-
Kimura M, Ohta T (1974) On some principles governing molecular evolution. Proc Natl Acad Sci 71(7): 2848- 2852.
-
Moore WS (1995) Inferring phylogenies from mtDNA variation: mitochondrial‐gene trees versus nuclear‐gene trees. Evolution 49(4): 718-726.
-
Neigel JE, Avise JC (1986) Phylogenetic relationships of mitochondrial DNA under various demographic models of spe- ciation. In: Karlin S, Nevo E, (Eds.), Evolutionary Processes and heory. Academic Press, Inc. pp: 515-533.
-
Hudson RR (1992) Gene trees, species trees, and the segregation of ancestral alleles. Genetics 131: 509-512.
-
Nichols R (2001) Gene trees and species trees are not the same. TREE 16(7): 358-364.
-
Rubinoff D, Cameron S, Will K (2006) A genomic perspective on the shortcomings of mitochondrial DNA for “barcoding” identification. J Hered 97(6): 581-594.
-
Kapli P, Yang Z, Telford MJ (2020) Phylogenetic tree building in the genomic age. Nat Rev Genet 21(7): 428- 444.
-
Hoelzer GA (1997) Inferring phylogenies from mtDNA variation: mitochondrial-gene trees versus nuclear-gene trees revisited. Evolution 51(2): 622-626.
-
Toews DP, Brelsford A (2012) The biogeography of mitochondrial and nuclear discordance in animals. Mol Ecol 21(16): 3907-3930.
-
Dong Z, Wang Y, Li C, Li L, Men X (2021) Mitochondrial DNA as a molecular marker in insect ecology: Current status and future prospects. Ann Entomol Soc America 114(4): 470-476.
-
Smith MA, Green DM (2004) Phylogeography of Bufo fowleri at its northern range limit. Mol Ecol 13(12): 3723-3733.
-
Babik W, Branicki W, Crnobrnja‐Isailović J, Cogălniceanu D, Sas I, et al. (2005) Phylogeography of two European newt species—discordance between mtDNA and morphology. Mol Ecol 14(8): 2475-2491.
-
Leaché AD, McGuire JA (2006) Phylogenetic relationships of horned lizards (Phrynosoma) based on nuclear and mitochondrial data: evidence for a misleading mitochondrial gene tree. Mol Phylogenet Evol 39(3): 628-644.
-
Qi Y, Lu B, Gao H, Hu P, Fu J (2014) Hybridization and mitochondrial genome introgression between Rana chensinensis and R. kukunoris. Mol Ecol 23(22): 5575- 5588.
-
Pabijan M, Zieliński P, Dudek K, Stuglik M, Babik W (2017) Isolation and gene flow in a speciation continuum in newts. Mol Phylogenet Evol 116: 1-12.
-
Haenel GJ (2017) Introgression of mt DNA in Urosaurus lizards: historical and ecological processes. Mol Ecol 26(2): 606-623.
-
Leavitt DH, Marion AB, Hollingsworth BD, Reeder TW (2017) Multilocus phylogeny of alligator lizards (Elgaria, Anguidae): testing mtDNA introgression as the source of discordant molecular phylogenetic hypotheses. Mol Phylogenet Evol 110: 104-121.
-
Freitas I, Ursenbacher S, Mebert K, Zinenko O, Schweiger S, et al. (2020) Evaluating taxonomic inflation: towards evidence-based species delimitation in Eurasian vipers (Serpentes: Viperinae). Amphibia-Reptilia 41(3): 285- 311.
-
Guayasamin JM, Castroviejo-Fisher S, Ayarzagüena J, Trueb L, Vilà C (2008) Phylogenetic relationships of glassfrogs (Centrolenidae) based on mitochondrial and nuclear genes. Mol Phylogenet Evol 48(2): 574-595.
-
Pyron RA, Burbrink FT, Wiens JJ (2013) A phylogeny and revised classification of Squamata, including 4161 species of lizards and snakes. BMC Evol Biol 13: 1-54.
-
Lourenço LB, Targueta CP, Baldo D, Nascimento J, Garcia PC, et al. (2015) Phylogeny of frogs from the genus Physalaemus (Anura, Leptodactylidae) inferred from mitochondrial and nuclear gene sequences. Mol Phylogenet Evol 92: 204-216.
-
Veith M, Bogaerts S, Pasmans F, Kieren S (2018) The changing views on the evolutionary relationships of extant Salamandridae (Amphibia: Urodela) PLoS One 13(8): e0198237.
-
Freitas S, Westram AM, Schwander T, Arakelyan M, Ilgaz Ç, et al. (2022) Parthenogenesis in Darevskia lizards: A rare outcome of common hybridization, not a common outcome of rare hybridization. Evolution 76(5): 899- 914.
-
Kimura M (1981) Possibility of extensive neutral evolution under stabilizing selection with special reference to non-random usage of synonymous codons. Proc Natl Acad Sci USA 78: 454-458.
-
Loewe L (2008) Negative Selection. Nature Education 1(1): 59.
-
Poe S, Latella I, Ayala-Varela F, Yañez-Miranda C, Torres- Carvajal O (2015) A new species of Phenacosaur Anolis (Squamata; Iguanidae) from Peru and a comprehensive phylogeny of Dactyloa-clade Anolis based on new DNA sequences and morphology. Copeia 103(3): 639-650.
-
Tilbury CR, Tolley KA (2015) Contributions to the herpetofauna of the Albertine Rift: Two new species of chameleon (Sauria: Chamaeleonidae) from an isolated montane forest, south eastern Democratic Republic of Congo. Zootaxa 3905(3): 345-364.
-
Marques MP, Parrinha D, Ceríaco LM, Brennan IG, Heinicke MP, et al. (2023) A new species of thick- toed gecko (Pachydactylus) from Serra da Neve and surrounding rocky areas of southwestern Angola (Squamata: Gekkonidae). Vertebrate Zoology 73: 325- 343.
-
Moreno-Arias RA, Méndez-Galeano MA, Beltrán I, Vargas-Ramírez M (2023) Revealing anole diversity in the highlands of the Northern Andes: New and resurrected species of the Anolis heterodermus species group. Vertebrate Zoology 73: 161-188.
-
Wiens JJ, Kuczynski CA, Arif S, Reeder TW (2010) Phylogenetic relationships of phrynosomatid lizards based on nuclear and mitochondrial data, and a revised phylogeny for Sceloporus. Mol Phylogenet Evol 54(1): 150-161.
-
Del Rosario Castañeda M, De Queiroz K (2011) Phylogenetic relationships of the Dactyloa clade of Anolis lizards based on nuclear and mitochondrial DNA sequence data. Mol Phylogenet Evol 61(3): 784-800.
-
Doughty P, Kealley L, Shoo LP, Melville J (2015) Revision of the Western Australian pebble-mimic dragon species- group (Tympanocryptis cephalus: Reptilia: Agamidae) Zootaxa 4039(1): 85-117.
-
Šmíd J, Moravec J, Kratochvíl L, Nasher AK, Mazuch T, et al. (2015) Multilocus phylogeny and taxonomic revision of the Hemidactylus robustus species group (Reptilia, Gekkonidae) with descriptions of three new species from Yemen and Ethiopia. Systematics and Biodiversity 13(4): 346-368.
-
Pizzigalli C, Crochet PA, Geniez P, Martínez‐Freiría F, Velo‐ Antón G, et al. (2021) Phylogeographic diversification of the Mesalina olivieri species complex (Squamata: Lacertidae) with the description of a new species and a new subspecies endemic from North West Africa. J Zool Syst Evol Res 59(8): 2321-2349.
-
Glaw F, Köhler J, Ratsoavina FM, Raselimanana AP, Crottini A, et al. (2023) A new large-sized species of leaf-tailed gecko (Uroplatus) from northern Madagascar. Salamandra 59(3): 239-261.
-
Wheeler QD (2000) The phylogenetic species concept (sensu Wheeler and Platnick) Species concepts and phylogenetic theory: a debate. Columbia University Press, New York, USA, pp: 55-69.
-
Dobzhansky T (1982) Genetics and the Origin of Species. Columbia University Press.
-
Mayr E (2000) A critique from the biological species concept perspective: what is a species, and what is not. Species concepts and phylogenetic theory: A debate. pp: 93-100.
-
Derraik JG, Early JW, Closs GP, Dickinson KJ (2010) Morphospecies and taxonomic species comparison for Hymenoptera. J Insect Sci 10(1): 108.
-
Cracraft J (1974) Phylogenetic models and classification. Syst Biol 23(1): 71-90.
-
Nixon KC, Wheeler QD (1990) An amplification of the phylogenetic species concept. Cladistics 6(3): 211-223.
-
Isaac NJ, Mallet J, Mace GM (2004) Taxonomic inflation: its influence on macroecology and conservation. TREE 19(9): 464-469.
-
Wiley EO (1978) The evolutionary species concept reconsidered. Syst Zool 27(1): 17-26.
-
Frost DR, Hillis DM (1990) Species in concept and practice: herpetological applications. Herpetologica 46: 87-104.
-
Wiley EO, Mayden RL (2000) A defense of the evolutionary species concept. Species concepts and phylogenetic theory. A debate. Columbia University Press, New York, USA, pp: 198-208.
-
DeQueiroz K (2007) Species concepts and species delimitation. Syst Biol 56(6): 879-886.
-
Padial JM, Miralles A, De la Riva I, Vences M (2010) The integrative future of taxonomy. Frontiers Zool 7(1): 1-14.
-
Mallet J (2005) Hybridization as an invasion of the genome. TREE 20(5): 229-237.
-
Moran BM, Payne C, Langdon Q, Powell DL, Brandvain Y, et al. (2021) The genomic consequences of hybridization. Elife 10: e69016.
-
Dollo L (1889) First note on the mosasaurians of Mesvin. Bulletin of the Belgian Society of Geology, Paleontology and Hydrology 3: 271-304.
-
Gregory WK (1936) On the meaning and limits of irreversibility of evolution. The American Naturalist 70(731): 517-528.
-
Bull JJ, Charnov, EL (1985) On irreversible evolution. Evolution 39(5): 1149-1155.
-
Schwenk K, Brede N, Streit B (2008) Introduction. Extent, processes and evolutionary impact of interspecific hybridization in animals. Phil Transact Royal Soc B: Biological Sciences 363(1505): 2805-2811.
-
Gabryś J, Kij B, Kochan J, Bugno-Poniewierska M (2021) Interspecific hybrids of animals-in nature, breeding and science–a review. Ann Anim Sci 21(2): 403-415.
-
Barton NH (2001) The role of hybridization in evolution. Mol Ecol 10(3): 551-568.
-
Arnold ML, Kentner EK, Johnston JA, Cornman S, Bouck AC (2001) Natural hybridisation and fitness. Taxon 50(1): 93-104.
-
Pfennig KS, Chunco AJ, Lackey AC (2007) Ecological selection and hybrid fitness: hybrids succeed on parental resources. Evol Ecol Res 9(2): 341-354.
-
Dobzhansky TH (1936) Studies on hybrid sterility. II. Localization of sterility factors in Drosophila pseudoobscura hybrids. Genetics 21(2): 113.
-
Shpak M (2005) The role of deleterious mutations in allopatric speciation. Evolution 59(7): 1389-1399.
-
Frankham R, Briscoe DA, Ballou JD (2002) Introduction to conservation genetics. Cambridge University Press.
-
Frankham R, Ballou JD, Eldridge MD, Lacy RC, Ralls K, et al. (2011) Predicting the probability of outbreeding depression. Conserv Biol 25(3): 465-475.
-
Muhlfeld CC, Kalinowski ST, McMahon TE, Taper ML, Painter S, et al. (2009) Hybridization rapidly reduces fitness of a native trout in the wild. Biol letters 5(3): 328- 331.
-
Burton RS, Pereira RJ, Barreto FS (2013) Cytonuclear genomic interactions and hybrid breakdown. Ann Rev Ecol Evol Syst 44: 281-302.
-
Wilson DS, Turelli M (1986) Stable underdominance and the evolutionary invasion of empty niches. American Natur 127(6): 835-850.
-
Calabrese GM, Pfennig KS (2020) Reinforcement and the proliferation of species. J Hered 111(1): 138-146.
-
Freudenstein JV, Broe MB, Folk RA, Sinn BT (2017) Biodiversity and the species concept—lineages are not enough. Syst Biol 66(4): 644-656.
-
Podnar M, Haring E, Pinsker W, Mayer W (2007) Unusual origin of a nuclear pseudogene in the Italian wall lizard: intergenomic and interspecific transfer of a large section of the mitochondrial genome in the genus Podarcis (Lacertidae) J Mol Evol 64(3): 308-320.
-
Nadachowska K, Flis I, Babik W (2010) Characterization of microsatellite loci in the Carpathian newt (Lissotriton montandoni). Herpetol J 20(2): 107-
-
Agapow PM, Bininda-Emonds OR, Crandall KA, Gittleman JL, Mace GM, et al. (2004) The impact of species concept on biodiversity studies. Quart Rev Biol 79(2): 161-179.
-
Nilson G, Tuniyev B, Andrén C, Orlov N (1999) Vipers of Caucasus: taxonomic considerations. Kaupia 8: 103-106.
-
Nagy ZT, Lawson R, Joger U, Wink M (2004) Molecular systematics of racers, whipsnakes and relatives (Reptilia: Colubridae) using mitochondrial and nuclear markers. J Zool Syst Evol Res 42(3): 223-233.
-
Gvoždík V, Moravec J, Klütsch C, Kotlík P (2010b) Phylogeography of the Middle Eastern tree frogs (Hyla, Hylidae, Amphibia) as inferred from nuclear and mitochondrial DNA variation, with a description of a new species. Mol Phylogenet Evol 55(3): 1146-1166.
-
Jablonski D, Kukushkin OV, Avcı A, Bunyatova S, Kumlutaş Y, et al. (2019) The biogeography of Elaphe sauromates (Pallas, 1814): with a description of a new rat snake species. Peer J 7: e6944.
-
Bannikov AG, Darevsky IS, Ishchenko VG, Rustamov AK, Szczerbak NN (1977) Guide to Amphibians and Reptiles of the USSR Fauna. Prosveshchenie, Moscow, pp: 414.
-
Weisrock DW, Macey JR, Ugurtas IH, Larson A, Papenfuss TJ (2001) Molecular phylogenetics and historical biogeography among salamandrids of the “true” salamander clade: rapid branching of numerous highly divergent lineages in Mertensiella luschani associated with the rise of Anatolia. Mol Phylogenet Evol 18(3): 434-448.
-
Veith M, Fromhage L, Kosuch J, Vences M (2006) Historical biogeography of Western Palaearctic pelobatid and pelodytid frogs: a molecular phylogenetic perspective. Contribut Zool 75(03-04): 109-120.
-
Tarkhnishvili D, Gokhelashvili R (1999) The amphibians of the Caucasus. Pensoft.
-
Servedio MR, Sætre GP (2003) Speciation as a positive feedback loop between postzygotic and prezygotic barriers to gene flow. Proc Royal Soc London, Series B: Biological Sciences 270(1523): 1473-1479.
-
Denzer W, Kaiser H (2023) Naming and gaming: The illicit taxonomic practice of ‘nomenclatural harvesting’and how to avoid it. J Zool 320(3): 161-168.
-
Koufopanou V, Burt A, Taylor JW (1997) Concordance of gene genealogies reveals reproductive isolation in the pathogenic fungus Coccidioides immitis. Proc Natl Acad Sci 94(10): 5478-5482.
-
Baird, NA, Etter PD, Atwood TS, Currey MC, Shiver AL, et al. (2008) Rapid SNP discovery and genetic mapping using sequenced RAD markers. PloS one 3(10): e3376.
-
Pante E, Abdelkrim J, Viricel A, Gey D, France SC, et al. (2015) Use of RAD sequencing for delimiting species. Heredity 114(5): 450-459.
-
Lawson DJ, Hellenthal G, Myers S, Falush D (2012) Inference of population structure using dense haplotype data. PLoS genetics 8(1): e1002453.
-
Mayr E (1947) Systematics and the origin of species.
-
Mayr E (1970) Populations, species, and evolution: an abridgment of animal species and evolution. Harvard University Press 19.
-
Szymura JM (1993) Analysis of hybrid zones with Bombina. In: Harrison RG (Ed.), Hybrid Zones and the Evolutionary Process, Oxford University Press: Oxford, USA, pp: 261-289.
-
Dufresnes C, Suchan T, Smirnov NA, Denoël M, Rosanov JM, et al. (2021) Revisiting a speciation classic: comparative analyses support sharp but leaky transitions between Bombina toads. J Biogeogr 48(3): 548-560.
-
Veith M, Steinfartz S, Zardoya R, Seitz A, Meyer A (1998) A molecular phylogeny of ‘true’salamanders (family Salamandridae) and the evolution of terrestriality of reproductive modes. J Zool Syst Evol Res 36(1‐2): 7-16.
-
Dufresnes C, Poyarkov NA, Jablonski D (2024) Reply to Lukhtanov: Polytypic species: Old wine in a new bottle. Proc Natl Acad Sci 121(9): e2321819121.
-
Lukhtanov VA (2024) Polytypic species concept and subspecies in the genomic era. Proc Natl Acad Sci 121(9): e2317038121.
-
Barton NH (1979) The dynamics of hybrid zones. Heredity 43(3): 341-359.
-
Barton NH, Hewitt GM (1989) Adaptation, speciation and hybrid zones. Nature 341(6242): 497-503.
-
Bierne N, Lenormand T, Bonhomme F, David P (2002) Deleterious mutations in a hybrid zone: can mutational load decrease the barrier to gene flow?. Genetics Research 80(3): 197-204.
-
Nikolsky AM (1913) Herpetologia Caucasica. The Caucasian Museum, Tiflis.
-
Stankowski S, Shipilina D, Westram AM (2021) Hybrid zones. eLS 2: 1-16.
-
Tarkhnishvili D, Hille A, Waller T, Todua M, Murtskhvaladze M, et al. (2022) Morphological trends and genetic divergence in anacondas, genus Eunectes Wagler, 1830 (Serpentes: Boidae). Amphibia-Reptilia 43(4): 379-393.
-
Terentyev PV, Chernov SA (1940) Field Guide of Reptiles and Amphibians of the USSR. Uchpedgiz, Leningrad.
-
Welch JJ, Bromham L (2005) Molecular dating when rates vary. TREE 20(6): 320-327.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind