An Expansion of the Invasive Species Gussevia asota Kritsky, et al., 1989 (Monogenea: Dactylogyridae) In Western Uttar Pradesh, India
Non-native fish species and their parasites are a threat to aquatic ecosystems, posing a risk to inherited communities by diminishing biodiversity and causing severe commercial and public health impacts. Along with their fish hosts, monogenean parasites are often co-introduced into new areas. In the western part of the state of Uttar Pradesh, India, a range expansion for the non-native monogenean parasite was recorded. This study confirmed the co-introduction of the dactylogyrid, Gussevia asota Kritsky et al. (1989), into India with their fish host, the invasive Oscar, Astronotus ocellatus Agassiz, 1831. The present species were also distinguished based on molecular analysis of their 18S rDNA sequence and the haptoral parts and male copulatory organ. Phylogenetic analysis of G. asota showed that it clustered with other dactylogyrid species, supporting the contention that it co-introduced parasites, increasing the number of monogeneans acquired in Indian water.
Introduction
Non-native host species introduce their non-native parasites into a new range of geographical locations that can help settle them into new habitats [1]. Several non- native parasite species are co-introduced into new regions with their original host species. These non-native parasites frequently persist as a part of the host’s fauna and can extend their range as the host increases their range [2, 3]. Sometimes a few co-introduced parasite species can switch to a new host species that is native to that particular region, though it spreads in the new environment [4, 5, 6, 7, 8]. The majority of non- indigenous fish hosts in India were imported for aquaculture and as ornamental fish, which can cause disease transmission and the co-introduction of new parasite species that might further increase the threats of unintended introduced non-native species for local fauna [1, 9]. As important fish parasites, Monogeneans are common parasites that are very host-specific. Reports on monogenean parasites co-introduced into India include various monogenean species, and most of them are host-specific and generally do not switch to other fish host species [10, 11, 12, 13, 14, 15, 16, 17]. The genus Gussevia was proposed as an ancryocephaline species by Kohn, et al. [18]. Gussevia Kohn, et al. [18] comprises three monogenean species Gussevia asota, Gussevia astronoti, and Gussevia rogersi that were described by Kritsky, et al. [19] from Astronotus ocellatus collected from countries Brazil and the United States. In all of the above mentioned species, particularly G. asota was found an important pathogen that may well kill their host [19, 20].
The main aim of the present study was to identify monogenean species Gussevia asota Kritsky, et al. [19] infecting invasive Oscar, Astronotus ocellatus Agassiz, 1831, in the Meerut region, a district of Uttar Pradesh, India, using ribosomal 18S DNA data and morphological analysis that make available to understand the phylogenetic relationships with other monogenean species.
Materials and Methods
Host-Parasite Collection and Morphological Analysis
Non-native Oscar Astronotus ocellatus was collected in the Meerut region, a district of Uttar Pradesh, India. In total, 38 Oscars were collected and transported alive in aerated boxes to the molecular taxonomy laboratory, department of Zoology, Chaudhary Charan Singh University, Meerut, Uttar Pradesh, India, and kept in aerated aquariums. Prior to the collection of helminth parasites, the fish specimens were subjected to euthanasia utilizing clove oil within three days of capture. They were further dissected for monogenean parasites, but before dissection, every fish host was measured for standard length (SL).
Collected dactylogyrid monogenean parasites from gill filaments were mounted in glycerine-ammonium-picrate [21] for semi-permanent slide preparation for morphological and morphometric analyses, while for permanent slides, the protocol of Verma, et al. [22] was followed. Monogeneans mounted in Canada balsam are deposited in the Museum of the Department of Zoology, Chaudhary Charan Singh University, Uttar Pradesh, India, and the Museum d’Histoire naturelle, Geneva, Switzerland. Morphological observations were made using an Olympus CH30 (Shinjuku, Tokyo, Japan) fitted with the image analysis software Axio Cam ERc5s. Drawings of haptoral hard parts were made with the aid of a drawing attachment. The morphological identification was based on the shape and measurements of the sclerotized parts of the haptor and reproductive organs, according to Kritsky, et al. [19]. All the measurements were taken in micrometers as the mean ± standard deviation, followed by the range in parentheses unless otherwise stated. A subsample of the collected parasites was preserved in 95% ethanol for further molecular analysis. Prevalence (in %) was calculated according to Bush, et al. [23].
DNA Extraction, PCR Amplification and Sequencing
Parasites collected in the present study, placed in 95% ethanol for genomic DNA extraction, were used to extract the DNA using a DNeasy Blood & Tissue Kit (QIAGEN, Hilden, Germany), following the method. The region of the 18S ribosomal DNA (rDNA) was amplified using the primers WormA+1270R and 930F+WormB [24]. The PCR reaction was performed at a final volume of 25 μL, according to Shinad, et al. [25], and the cycling conditions were as follows: initial denaturation for 2 min at 95 °C, 35 cycles of 55 s at 95 °C, 50 s at 55 °C, 1 min at 72 °C, and a final extension for 7 min at 72 °C. Obtained PCR products were electrophoresed on 1.5% agarose gels and then purified using the PurelinkTM Quick Gel Extraction and PCR Purification Combo Kit (Invitrogen, Löhne, Germany), following the manufacturers protocol. The purified PCR products were then sequenced directly in both directions using the same primers as mentioned above using a Big Dye® Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, Thermo Fisher Scientific, Foster City, California, USA). Generated DNA sequences were assembled and edited using the Geneious Pro 5.4 platform [26], and the newly generated monogenean sequences were deposited in GenBank for accession numbers.
Molecular phylogeny was analyzed using maximum likelihood (ML) and Bayesian inference (BI) methods. Sequences of other monogenean species for phylogenetic reconstruction were retrieved from GenBank. For the alignment of 18S gene sequences, MUSCLE algorithm implemented in Molecular Evolutionary Genetics Analysis (MEGA) version 11 [27] was used with default parameters. Model Finder [28] was used to infer the ideal evolutionary model for the 18S gene using the Bayesian information criterion. The GTR + I + G evolutionary model were selected in the monogenean phylogeny of the current species. To conduct ML analysis, the program MEGA version 11 was used with nodal support assessed by 1,000 bootstrapping replicates. The BI phylogenetic analysis was performed with TOPALi version 2.5 software [29], using four simultaneous chains of the Markov Chain Monte Carlo (MCMC) algorithm run twice for 1,000,000 generations. For every 100 generations, the tree topologies were sampled, whereby the first 25% of trees from each run were discarded as burn-in. Finally, the genetic divergence was calculated using uncorrected p-distances for the 18S gene in MEGA version 11. The species of Scutogyrus and Cichlidogyrus was used as outgroup.
Results
Family Dactylogyridae Bychowski, 1933 Genus Gussevia Kohn and Paperna, 1964 Gussevia asota Kritsky, Thatcher and Boeger, 1989 Description (n= 27) (Figure 1: Gussevia asota microphotograph). Body-elongated, fusiform, moderate- sized worms with a narrow cephalic region; maximum width attained at the level of gonads. Cephalic lobes poorly developed, having two terminals and two bilateral. Two pairs of eyes, equidistant; the posterior pair larger than anterior pair. Ovate shaped pharynx present. Well-developed haptor present with anterior and posterior lobes having small glandular patches. Dorsal anchor, variable in length with superficial root, shaft curved with elongate point. Dorsal bar rod-shaped have slightly expanded ends. Ventral anchor comprises poorly differentiated roots, short anterior projection arising from tip of superficial root, shaft short with acute point. Filamentous ventral bar has ends curling around ventral anchor base. Hook pairs 1, 2, 3, 4, 6, and 7 similar with a slender shank; hook pair 5 modified, usually elongate. Eggs spherical in shape and smooth outline. Cirrus comprises a base from which a double-walled, coiled tube arises, having a coil of about 1.5 rings. Accessory piece basally articulated with cirrus having terminal flabellate piece possessing sclerotized ring. Vagina not observed. Vitellaria dense, dispersed along through intestinal caeca.
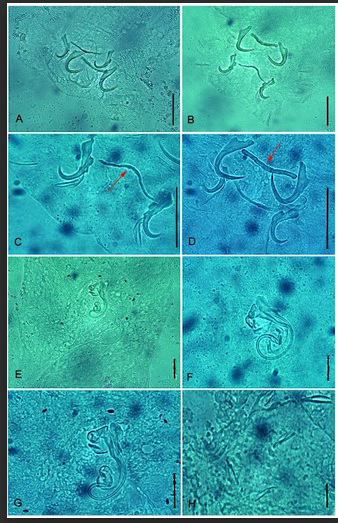
Figure 1: Micrographs of Gussevia asota Kritsky et al. (1989) from Astronotus ocellatus. (A) and (B) Haptoral armature; (C) An enlarged view of the ventral anchors and ventral bar (VB), VB indicated by red arrow; (D) An enlarged view of the dorsal anchors and dorsal bar (DB), DB indicated by red arrow; (E) Male copulatory organ; (F) and (G) Two different enlarged views of male copulatory organ; (H) Marginal hooks. Scale bars: 35 µm (A, D), 32 µm (B), 30 µm (C) 23 µm (E) 12 µm (F) 17 µm (G) 11 µm (H).
Taxonomic Summary
Host: Oscar, Astronotus ocellatus Agassiz, 1831 (Cichliformes: Cichlidae). Locality: Meerut (29˚01′N and 77˚45′E), Uttar Pradesh, India.
Site of infection: Gill filaments. Prevalence: 71.05% (27 host infected out of 38); a range of >50 to 200 parasites per fish. Material deposited: Voucher specimens were submitted to the collection of the Museum, Department of Zoology, Chaudhary Charan Singh University, Meerut (U.P.), India
(HS/mon/2022/13) and to the Museum d’Histoire Naturelle, Geneva, Switzerland (MHNG-PLAT-0144330). Molecular sequence data: For 18S gene, we have generated sequences from two isolates of Gussevia asota: PP352272 (1787 bp), PP352273 (1695 bp).
Remarks: This is the second record of G. asota from India based on morphological and molecular data. It is characterized primarily by having a dextral vagina, cirrus with a long spiral tube, and an accessory piece attached to the distal end of the cirrus. Gussevia asota was originally described from the gill filaments of Astronotus ocellatus from Brazil. This parasite was also reported from various geographical regions: East Asia [30]; Europe [20]; North America [19]; South America [19, 31, 32, 33]. From the Indian region, this parasite was reported by Tripathi, et al. [17] and exhibited some more features, like the presence of a vaginal pore that is present in the dextro-marginal, funnel-shaped, and large median seminal receptacle (Table 1).
Study of the present specimen showed that this is conspecific to previous descriptions of G. asota [17, 19, 31]. In our study, this parasite was morphologically compared with isolates of other geographical regions. Among them, it shows maximum similarity (in respect of body length) with isolates from Peru [33] (Table 1). The present species comprises features that are a little bit different from the original description in having: large worms, well-developed anterior and posterior lobes in the haptoral region, and small glandular patches, as also shown in numerous studies [17, 20, 31] (Table 1).
| Body parts | G. asota Kritsky et al. (1989) Brazil | G. asota Mendoza- Franco et al. (2010) Peru | G. asota Smigh et al. (2016) Slovakia | G. asota Tripathi and Matey (2023) India | G. asota Present study India |
|---|---|---|---|---|---|
| Body: | |||||
| Body length | 394(380-462) | 630 | 544(540-564) | - | 634.46±19.93 (606.67- 666.67) |
| Body width | 88(73-111) | 103 | 92(81-121) | - | 146.18±14.74 (126.47- 166.62) |
| Pharynx diameter | 25(21-30) | 23-29 | - | - | 30.59±2.45 (26.67- 33.33) |
| Haptor length | 42(34-54) | - | 90-97 | - | 123.73±7.25 (113.75- 130.76) |
| Haptor width | 66(61-78) | 160 | 105(102-111) | - | 145.65±3.90 (140.33- 150) |
| Dorsal anchor: | |||||
| Length | 27(24-30) | 26(25-27) | 27(24-28) | 27 (27-28) | 28.27±2.23 (25-31.33) |
| Base width | 12(10-13) | 12(10-13) | 7(6-8) | - | 6.86±0.88 (5.33-8) |
| Dorsal bar: | |||||
| Length | 31(23-36) | 33(31-38) | 23-36 | 32 (31-36) | 32.5±1.57 (31-35) |
| Width | - | - | - | - | 2.99±0.62 (2.33-4) |
| Ventral anchor: | |||||
| Length | 26(25-28) | 25(24-27) | 26(24-29) | 24 (23-26) | 26.16±1.30 (24-27.33) |
| Base width | 14(12-16) | 18(17-19) | 7(5-8) | - | 5.64±0.99 (4.33-7) |
| Ventral bar | |||||
| Length | 31(24-35) | 32(30-33) | 26-35 | 31 (26-36) | 33.66±3.31(29-36.33) |
| Width | - | - | - | - | 1.55±0.41 (1-2) |
| Female reproductive organs: | |||||
| Ovary length | |||||
| Ovary width | 109(85-132) | - | - | - | 142.30±6.90 (135.89- 153.84) |
| Egg length | 45(36-57) | - | - | - | 43.07±4.41 (35.89- 48.71) |
| Egg width | - | - | - | - | 70.91±2.08 (68.75- 74.16) |
| - | - | - | - | 63.24±1.90 (60.83- 66.25) | |
| Male reproductive organs: | |||||
| Testis length | 90(71-108) | - | - | - | 101.11 ±11.38 (82.5- 115.38) |
| Testis width | 40(33-51) | - | - | - | 41.66±4.92 (35.89- 48.71) |
| Cirrus length | 61-62 | 30(28-35) | 54-73 | 57 (56-62) | 56.91±5.05 (52.67- 65.33) |
| Proximal ring diameter | 14(10-16) | - | - | - | 13.67±1.44 (11-15.33) |
| Accessory piece length | 27(23-29) | 29(26-31) | - | 22 (24-40) | 31.58±3.70 (27-36.67) |
| Hooks length: | |||||
| 1,2,3,4,6,7th pair | 11(10-12) | 11(10-12) | - | 11 (11-12) | 11.55±0.98 (10.16-13) |
| 5th pair | 14(13-15) | 15(14-15) | - | 14 (13-15) | 14.38±0.47 (13.83-15) |
Table 1: A comparison of the measurements of Gussevia asota Kritsky, et al. (1989) parasitizing Astronotus ocellatus.
Phylogenetic Analysis
The final alignment of the 18S rRNA gene was used for phylogenetic reconstruction, and the species of the genera Scutogyrus and Cichlidogyrus were used as an-outgroup. Identical sequences of 18S rDNA were obtained from two specimens in this study. For alignment of the 18S dataset, selected monogenean parasite data from GenBank was retrieved. The topologies generated using the ML and BI methods were similar; therefore, ML reconstruction with bootstrap support and posterior probabilities outcomes from both analyses were shown along their nodes (Figure 2). The 18S rDNA phylogeny using both ML and BI demonstrated that the sequences generated in the present study and sequences for other isolates of the same species from the same host, A. ocellatus, Peru, form a well-supported group (Figure 2). A sister clade to the second grouping, comprised of G. astronoti from the same host A. ocellatus, Peru was also shown. The intraspecific range for isolates of G. asota was 0.07%, while the interspecific range of isolates of G. asota with G. astronoti was 0.30%.
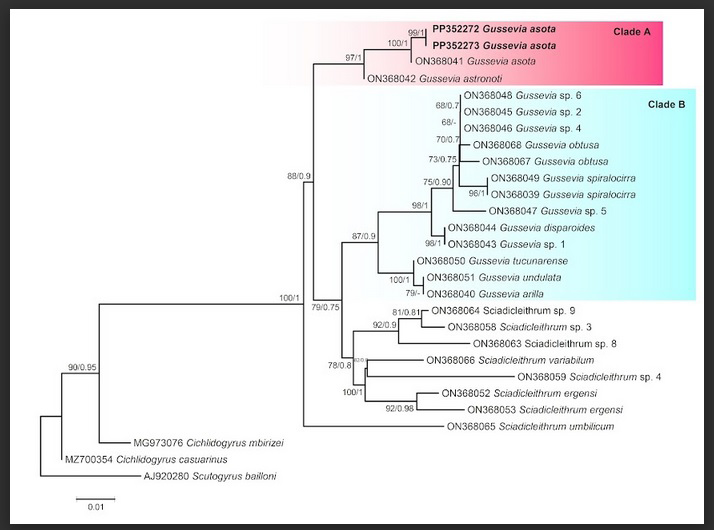
Figure 2: Maximum likelihood phylogenetic tree of Gussevia asota parasitizing cichlid fish based on 18S rRNA sequences. Values alongside branches designate bootstrap values and posterior probabilities resulting from maximum likelihood (ML) and Bayesian inference (BI) analyses. A and B letters symbolized the well-supported clades. Values below 0.70 (BI) are shown as dashes. The GenBank accession numbers of the species used in the phylogenetic analysis are also shown. The branch-length scale bar indicates the number of substitutions per site.
The phylogenetic reconstruction for Gussevia species in the present study revealed two well-supported clades parasitizing Neotropic cichlids. Gussevia asota and G. astronoti parasitizing cichlid A. ocellatus from Peru belongs to South America, formed clade A, while Gussevia species that also parasitizing cichlids of South American countries, Peru and Venezeula, were nested together in clade B (Figure 2). Representatives of the genus Gussevia show a polyphyletic assemblage in the present analysis, whereas G. asota and G. astronoti, with Indian isolates of G. asota (clade A) and other Gussevia species of South American cichlids (clade B), formed a well-supported monophyletic group.
Discussion
The present study reports the presence of an neotropical cichlid, Astronotus ocellatus, in Meerut, western Uttar Pradesh, India. This cichlid, A. ocellatus was originally found in the Amazon basin, in the countries of Peru, Colombia, and Brazil [34, 35]. This species has growing economic interest as an ornamental fish and is abundantly consumed for its delicious meat [35, 36, 37]. Insufficient controls of fish species imports and exports at quarantine places enable the spreading of novel and undocumented pathogen or parasite species [30].
In spite of the great economic value of A. ocellatus, studies appraising monogenean infection in Indian water for non- native fish parasites are still scarce. This is the first report of high infection by G. asota in A. ocellatus (found >50 to 200 parasites per fish) in western Uttar Pradesh, India, and it establishes that this fish pathogen is spreading in the global ornamental fish trade. In the present study, for the first time, we have amplified the 18S ribosomal RNA gene for molecular characterization and to supplement the morphology of the parasite. The occurrence of G. asota was confirmed in this study, i.e., in the western UP district of Meerut, and in a study by Tripathi, et al. [17] in the northern UP district Lucknow, in New Delhi, and in the eastern part, i.e., Kolkata, India. These reports show the occurrence of this parasite species, which agrees with our results, indicate high prevalence in both studies, and show the potential and probability of this parasite species co-introduction into new areas. According to a study on the Neotropical cichlid monogeneans by Seidlová, et al. [38], we also have similar observations. Our phylogenetic analysis indicated the inclusion of G. asota Indian isolates clustered with Peru isolates of the same species and another species, G. astronoti parasitizing cichlid host (Figure 2). Seidlová, et al. [38] have suggested a monophyletic origin for the monogenean parasites of Neotropical cichlids which is also consistent with our results. Furthermore, Seidlová, et al. [38] proposed Gussevia as monophyletic group with which the current results shows not in agreement and indicating the requirement for the taxonomic revision of Gussevia.
India has obtained less responsiveness for investigations regarding parasite co-introduction by exotic fishes as compare to other geographical regions [10, 11, 12, 13, 14, 15, 16, 17]. Although the National Fisheries Development Board (NFDB), Ministry of Fisheries, the Government of India implemented guidelines to import ornamental fishes in India and to control the entry of exotic ornamental fish along with associated parasites [39]. As per the conventions of the National Fisheries Development Board, Government of India [39], Oscar is one of the 92 commercially significant freshwater ornamental fish that are authorized to be imported in India. The Oscars have the capability to survive and reproduce in non-native environmental tolerances. For example, hypoxia, the ability to colonize interrupted habitats, generalist feeding behaviors, and reproductive potential in association with fast growth [40, 41, 42, 43]. Moreover, to reduce the risk of the introduction of non-native parasites and their associated diseases, quarantining is one of the essential biosecurity measures. In the course of this period, imported fish are detained in a quarantine facility in which they can be monitored, made a diagnosis, and treated successfully beforehand before being discharged to a farm [44]. According to the International Union for Conservation of Nature (IUCN), A. ocellatus is listed as a species of “least concern.” It is widely distributed across South America, including the western Amazon and Orinoco basins, but has been translocations throughout elsewhere under the ornamental fish trade [45], and as a result, it is now present in the various geographical localities, which also shows the tight host specificity of G. asota.
Conclusion
In conclusion, our study confirmed the presence of the monogenean fauna of Neotropical cichlid A. ocellatus in India. Morphology, morphometry, and molecular data generated subsidize the existing literature about the invasive monogenean parasite G. asota. Additionally, the generated taxonomic evidence for this parasite may be employed in future studies to track the cause and range of its invasive hosts in the Indian freshwater system and worldwide.
Conflicts of Interest
The authors declare no competing interests.
Acknowledgements
We are grateful to the Head, Department of Zoology, Chaudhary Charan Singh University, Meerut, Uttar Pradesh India, for providing laboratory facilities.
References
-
Taraschewski H (2006) Hosts and parasites as aliens. Journal of Helminthology 80: 99-128.
-
Lymbery AJ, Morine M, Kanani HG, Beatty SJ, Morgan DL (2014) Co-invaders: the effects of alien parasites on native hosts. International Journal for Parasitology: Parasites and Wildlife 3: 171-177.
-
Kvach Y, Ondrackova M, Kutsokon Y, Dzyziuk N (2018) New Record of Monogenean Parasites on Non-Indigenous Fishes in the Ukrainian Danube Delta. BioInvasions Records 7: 65-72.
-
Škoríková B, Scholz T, Moravec F (1996) Spreading of introduced monogeneans Pseudodactylogyrus anguillae and P. bini among eel populations in the Czech Republic. Folia Parasitologica 43: 155-156.
-
Ondračková M, Matějusová I, Šimková A, Gelnar M (2004) New reports of dactylogyridean species (Monogenea) for Central Europe. Helminthologia 41: 139-145.
-
Simková A, Morand S, Jobet E, Gelnar M, Verneau O (2004) Molecular phylogeny of congeneric monogenean parasites (Dactylogyrus): a case of intrahost speciation. Evolution 58: 1001-1018.
-
Galli P, Strona G, Benzoni F, Crosa G, Stefani F (2007) Monogenoids from freshwater fish in Italy, with comments on alien species. Comparative Parasitology 74: 264-272.
-
Ondračková M, Seifertová M, Tkachenko MY, Vetešník L, Liu H, et al. (2023) The parasites of a successful invader: monogeneans of the Asian topmouth gudgeon Pseudorasbora parva, with description of a new species of Gyrodactylus. Parasite 30: 22.
-
Kmentová N, Van Steenberge M, Thys van den Audenaerde DFE, Nhiwatiwa T, Bukinga FM, et al. (2019) Co-introduction success of monogeneans infecting the fisheries target Limnothrissa miodon differs between two non-native areas: the potential of parasites as a tag for introduction pathway. Biological Invasions 21: 757- 773.
-
Sharma P, Agarwal N, Kumar S (2011) Ribosomal DNA and morphological analysis of Dactylogyrus species from freshwater fishes of India. Journal of Parasitic Diseases 35: 210-214.
-
Chaudhary A, Verma C, Varma M, Singh HSS (2014) Identification of Thaparocleidus caecus (Mizelle & Kritsky, 1969) (Monogenea: Dactylogyridae) using morphological and molecular tools: a parasite invasion in Indian freshwater. BioInvasions Records 3: 195-200.
-
Chaudhary A, Verma C, Singh H (2016) First report on the molecular characterization of Diaphorocleidus armillatus Jogunoori et al. 2004 (Monogenea: Dactylogyridae) infecting the gills of introduced fish, Gymnocorymbus ternetzi in India. Acta Parasitologica 61: 639-644.
-
Chaudhary A, Chiary HR, Singh HS (2017) First molecular confirmation of the Dactylogyrus anchoratus and D. vastator (Monogenea, Dactylogyridae) from Carassius auratus in western India. BioInvasions Records 6: 79-85.
-
Chiary HR, Chaudhary A, Singh HS, Goswami UC (2014) Molecular characterization of two non-native species of Dactylogyrus (Monogenea: Dactylogyridae) recovered from introduced hosts in India. BioInvasions Records 3: 297-300.
-
Tripathi A (2015) Monogenoidea on exotic Indian freshwater fish. 3. Are Indian guidelines for importation of exotic aquarium fish useful and can they be implemented; the case of Neotropical Gussevia spiralocirra Kohn and Paperna, 1964. Current Science 108: 2101-2105.
-
Tripathi A, Matey C, Agarwal N (2022) Monogenoidea on exotic Indian freshwater fish. 4. Dactylogyrus minutus from Platinum ogon, an ornamental variety of the common carp Cyprinus carpio (Cypriniformes, Cyprinidae). BioInvasions Records 11: 510-523.
-
Tripathi A, Matey C (2023) Monogenea on exotic Indian freshwater fish. 5. First report of pathogenic Gussevia asota (Platyhelminths) from Oscar Astronotus ocellatus (Agassiz 1831) (Perciformes: Cichlidae). Zootaxa 5237: 52-64.
-
Kohn A, Paperna I (1964) Monogenetic trematodes from aquarium fishes. Revista Brasileira de Biologia 24: 145- 149.
-
Kritsky DC, Thatcher VE, Boeger WA (1989) Neotropical Monogenea. 15. Dactylogyrids from the gills of Brazilian Cichilidae with proposal of Sciadicleithrum gen. n. (Dactylogyridae). Proceedings of the Helminthological Society of Washington 56: 128-140.
-
Šmiga L, Košuthová L, Koščo J, Košuth P, Pekárik L, et al. (2016) First report of Gussevia asota (Monogenea: Dactylogyridae), destructive parasite of Astronotus ocellatus (Perciformes: Cichlidae) in Europe. Biologia 71: 547-550.
-
Malmberg G (1970) The excretory system and the marginal hooks as a basis for the systematics of Gyrodactylus (Trematoda: Monogenea). Arkiv för Zoologi 23: 1-235.
-
Verma C, Chaudhary A, Shanker Singh H (2017) Redescription of two species of Thaparocleidus (Monogenea: Dactylogyridae), with the description of T. armillatus sp. n. from Wallago attu and a phylogenetic analysis based on 18S rDNA sequences. Acta Parasitologica 62: 652-665.
-
Bush AO, Lafferty KD, Lotz JM, Shostak AW (1997) Parasitology meets ecology on its own terms: Margolis et al. revisited. Journal of Parasitology 83: 575-583.
-
Littlewood DTJ, Olson PD (2001) Small subunit rDNA and the phylum Platyhelminthes: signal, noise, conflict and compromise. In: Littlewood DTJ, Bray RA (Eds.), Interrelationships of the Platyhelminthes. Taylor and Francis, London, UK, pp: 262-278.
-
Shinad K, Chaudhary A, Prasadan PK, Singh HS (2021) Phylogenetic position of Tremiorchis ranarum Mehra and Negi, 1926 (Trematoda: Plagiorchiidae) with remark on this monotypic genus. Parasitology International 84: 102398.
-
Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, et al. (2012) Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28: 1647- 1649.
-
Tamura K, Stecher G, Kumar S (2021) MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Molecular Biology and Evolution 38: 3022-3027.
-
Kalyaanamoorthy S, Minh BQ, Wong TKF, von Haeseler A, Jermiin LS (2017) ModelFinder: fast model selection for accurate phylogenetic estimates. Nature methods 14: 587-589.
-
Milne I, Lindner D, Bayer M, Husmeier D, McGuire G, Marshall DF, et al. (2009) TOPALi v2: a rich graphical interface for evolutionary analyses of multiple alignments on HPC clusters and multi-core desktops. Bioinformatics 25: 126-127.
-
Kim JH, Hayward CJ, Joh SJ, Heo GJ (2002) Parasitic infections in live freshwater tropical fishes imported to Korea. Diseases of aquatic Organisms 52: 169-173.
-
Abdallah VD, Azevedo RK, Luque JL (2008) Notes on the morphology of two species of Gussevia Kohn & Paperna (Monogenea: Dactylogyridae) parasitic on Astronotus ocellatus (Agassiz) (Perciformes: Cichlidae) from Brazil. Pan-American Journal of Aquatic Sciences 3: 101-104.
-
Mendoza-Franco EF, Aguirre-Macedo ML, Vidal- Martínez VM (2007) New and previously described species of Dactylogyridae (Monogenoidea) from the gills of Panamanian freshwater fishes (Teleostei). Journal of Parasitology 93: 761-771.
-
Mendoza-Franco EF, Scholz T, Rozkosná P (2010) Tucunarella n. gen. and other dactylogyrids (Monogenoidea) from cichlid fish (Perciformes) from Peruvian Amazonia. Journal of Parasitology 96: 491-498.
-
Pavanelli GC (2000) Sanidade de peixes, rãs, crustáceos e moluscos. In: Valenti WC, Poli R, et al. (Eds.), Aqüicultura no Brasil: bases para um desenvolvimento sustentável, CNPQ: Brasília, pp: 208-209.
-
de Carvalho AFS, Isadora de LA, Paiva IM, Mansur VFR, Castro TFD, et al. (27) Identification of sex in Astronotus ocellatus through the evaluation of aggressive behavior and sexual steroid plasma level. Aquaculture 481: 239- 244.
-
Delgado PM, Delgado JPM, Orbe RI (2014) Parasitism by Gussevia asota in gills of juveniles of Astronotus ocellatus cultured in the Peruvian Amazon. Journal of Fisheries Sciences 8: 61-66.
-
Tavares-Dias M, Sousa TJSM, Neves LR (2014) Parasitic infections in two Benthopelagic fish from Amazon: the Arowana Osteoglossum bicirrhosum (Osteoglossidae) and Oscar Astronotus ocellatus (Cichlidae). Bioscience Journal 330: 546-555.
-
Seidlová L, Benovics M, Šimková A (2022) Gill monogeneans of neotropical cichlid fish: diversity, phylogenetic relationships, and host-parasite cophylogenetic associations. International Journal for Parasitology 52: 603-615.
-
National Fisheries Development (NFDB) (2022) National Fisheries Development Board.
-
Ferreira EJG (1981) Alimentação dos adultos de doze espécies de cichlideos (Perciformes, Cichlidae) do rio Negro, Brasil. M.Sc. Thesis, INPA/FUA, Manaus, pp: 242.
-
Muusze B, Marcon J, van den Thillart G, Almeida-Val V (1998) Hypoxia tolerance of Amazon fish: Respirometry and energy metabolism of the cichlid Astronotus ocellatus. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology 120: 151-156.
-
Soares MGM, Costa EL, Siqueira-Souza FK, Anjos HDB, Yamamoto KC, et al. (2008) Peixes de lagos do médio Rio Solimões. Manaus, Amazonas, pp: 175.
-
CABI (2022) Astronotus ocellatus (Oscar) Invasive Species Compendium.
-
Hadfield CA, Clayton LA (2011) Fish quarantine: current practices in public zoos and aquaria. Journal of Zoo and Wildlife Medicine 42: 641-650.
-
Froese R, Pauly D (2022) FishBase. World Wide Web electronic publication.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind