On Some Acanthocephalans from Wildlife in Peru, with the Designation of Neoechinorhynchus (Neoechinorhynchus) cylindratus peruensis n. subsp. and Special Notes on the Status of “Polymorphus pacificus” sensu Karl, 1967
Cystacanths and adults of 8 valid species of acanthocephalans in 3 classes that we collected from various species of wildlife in Peru over the last few years are reported. Notes on each species are supplemented by light microscope images giving new perspectives even though some have been previously published using line drawings or SEM images. The presented images are novel and original for each species and enhance some of the former depictions of species that may have been previously published. This presentation includes a treatment of the cystacanth of a moniliformid species, Moniliformis moniliformis (Bremser, 1811) Travassos, 1915 (Class Archiacanthocephala) from Rattus rattus (Linn.) in Lima, Peru. A new form of Neoechinorhynchus (Neoechinorhynchus) cylindratus (Van Cleave, 1913) Van Cleave, 1919 (Class Eoacanthocephala) different from the usual North American form collected from the mosquitofish Gambusia affinis (Baird & Girard) in Pantanos de Villa Wildlife refuge is designated as a new subspecies: Neoechinorhynchus (Neoechinorhynchus) cylindratus peruensis n. subsp. Species in 3 families of Class Palaeacanthocephala, Echinorhynchidae, Illiosentidae, and Polymorphidae are reported. Pseudoacanthocephalus lutzi (Hamann, 1891) (Echinorhynchidae) was found in the coastal toad Chaunus limensis Werner off Lima. Illiosentis furcatus Van Cleave & Lincicome 1939 (Illiosentidae) is reported from the Lorna drum Sciaena deliciosa Tschudi in La Punta. The following species in Polymorphidae are also reported. Cystacanths of Polymorphus spindlatus Amin & Heckmann, 1991 in 4 species of killifish Orestias in Lake Titicaca and in the Nile Tilapia Oreochromis niloticus Linn. in Villa swamps. Adult P. spindlatus were described from the black-crowned night heron at Puno Bay, Lake Titicaca. Cystacanths of Profilicollis altmani (Perry, 1942) Van Cleave, 1947 were described from the Pacific sand crab Emerita analoga (Stimpson) in Chorrillos, Lima and the adults from Belcher’s gull Larus belcheri (Vigors) in Ancȯn, Lima. The cystacanths of Neoandracantha peruensis Amin & Heckmann, 2017 were described from the ghost crab Ocypod gaudichaudii Milne-Edwards & Lucas from the Pacific coast off Lima. The questionable validity of “Polymorphus (=Profilicollis) pacificus” sensu Karl, 1967 is discussed and that species appears to be an extreme variant of P. altmani.
Omar M Amin1*, César Peña2, Teresa Castro3 and Nataliya Rubtsova1
Keywords: Neoechinorhynchus; Polymorphus spindlatus; Peruvian Parasitology
Introduction
Our parasitology team has performed several investigations of wildlife parasitology over a number of years on various occasions designed around making field collections with the support of local institutions and parasitologists with common interests. Dr. Heckmann pioneered this involvement with the capturing of black-crowned night herons from Puno Bay, Lake Titicaca in the late 1980th leading to the study and description of adult Polymorphus spindlatus Amin, et al. [1]. Subsequent research of the host systems associated with the life cycle of P. spindlatus was published by Amin, et al. [2]. Dr. César Peña and Teresa Castro Barrientos as well as Dr. José Iannacone, President of the Associación Peruana de Helmintologia e Invertebrados Afines (APHIA), Lima, Peru have gifted us (OMA) a large collection of acanthocephalans while participating in two Peruvian parasitology conferences in Lima. The first was the October 2009: International Symposium of Aquatic Environment, Parasitology, at Lima, Peru. The second was the IV Congresso International de Parasitología Neotropical (IV COPANEO) in Lima in 2015. Studies of the earlier collections of Peña, Castro and Iannacone in the 1970s and 1980s provided much of the material used for the publication of our detailed work on Profilicollis altmani (Perry, 1942) Van Cleave, 1947 in Amin, et al. [3], larval stages of P. spindlatus in Oreochromis niloticus in Amin, et al. [4], Pseudoacanthocephalus lutzi (Hamann, 1891) in the coast toad Chaunus limensis Werner in Amin, et al. [5], and a new form of Neoechinorhynchus cylindratus (Van Cleave, 1913) Van Cleave, 1919 which is being designated as a new subspecies from the mosquito fish Gambusia affinis (Baird & Girard) from Pantanos de Villa Amin, et al. [6]. Specimens of Neoandracantha peruensis Amin and Heckmann, 2017 were described based on material partially provided by another Peruvian researcher, Asucena Naupay of the Universidad Nacional Mayor de San Marcos during the 2015 Congress. The tireless and well-designed field work (collecting and examining a large number of crabs in various coastal locations) provided by Dr. José Iannacone and his students at Universidad Nacional Federico Villarreal, San Marcos contributed to the collections of the N. peruensis material making possible the description of this important new genus of acanthocephalans by Amin, et al. [7]. Ianacone also provided specimens of Corynosoma sp. from the Peruvian rock bass Paralabrax humeralis (Valenciennes), cystacanths of Moniliformis moniliformis (Bremser, 1811) Travassos, 1915 from the black rat Rattus rattus (Lin.), and cystacanths of Profilicollis altmani (Perry, 1942) Van Cleave, 1947 from Emerita analoga (Stimpson).
The above introductory section is limited to remarks directly related to our work leading to the procuring of specimens used in this presentation. No reference is made here to the many other publications on other wildlife parasites mostly by peruvian researchers including Mateo, Pulido Capurro, Guzmán, Iannacone, Peña, Naupay, Gomez-Puerta, Tantaleán, Ortega, Gonzales-Viera, Garcia Varela, Pinacho- Pinacho, Obispo, Pérez-Ponce de León, Ancona, Luque, Brito- Porto, Riquelme, Montes, Srean, Asencios, Huffman, Calhoun, Cossios, Johnson, Leggett, Cordova, Castro, Welcomme, and Gomez-Puerta.
Materials and Methods
Processing for Microscopical Examination
Creating whole mounts of adults and cystacanths varied in different labs and locations. Specimens processed in Peru including P. altmani, P. spindlatus cystacanths, P. lutzi, N. cylindratus, and I. furcatus, were stained in Borax carmine and occasionally with fast green before being dehydrated in ethanol then whole mounted. Specimens processed by us in Scottsdale, Arizona including adult P. spindlatus, and N. peruensis, were stained in Mayer’s carmine, detained in 4% hydrochloric acid in 70% ethanol, dehydrated in ethanol, cleared in 100% xylene or in graded terpineol-100% ethanol, and mounted in Canada balsam.
Processing for Histopathological Studies
Infected host tissues were fixed in buffered 10% formalin. After dehydration and embedding in paraffin, sectioned at 4-6 um, fixed on glass slides and stained in hematoxylin and eosin (H & E) or Mallory’s triple stain (MT). The slides were studied with a Zeiss compound microscope or with a BH2 light Olympus microscope (Olympus Optical Co., Osachi- Shibamiya, Okaya, Nagano, Japan).
Line Drawings
Line drawings were made by using a Ken-A-Vision micro-projector (Ward’s Biological Supply Co., Rochester, New York) which uses cool quartz iodine 150W illumination with 10X, 20X, and 43X objective lenses. Images of stained whole mounted specimens were projected vertically on 300 series Bristol draft paper (Strathmore, Westfield, Massachusetts), then traced and inked with India ink. Projected images were identical to the actual specimens being projected. Microscope images were created using 10X and 40X objective lenses of a BH2 light Olympus microscope (Olympus Optical Co., Osachi-shibamiya, Okaya, Nagano, Japan) attached to an AmScope 1000 video camera (United Scope LLC, dba AmScope, Irvine, California), linked to an ASUS laptop equipped with HDMI high-definition multimedia interface system (Taiwan-USA, Fremont, California). Images from the microscope are transferred from the laptop to a USB and stored for subsequent processing on a computer.
Optical Microscopy
Images created for this presentation were acquired using a Zeiss Axioskop Transmitted Nomarski DIC Phase Contrast Microscope Trinocular (Munich, Germany) and a Canon T3i EOS 600D DSLR Camera (Melville, New York). Measurements are in micrometers unless otherwise noted; the range is followed by the mean values between parentheses when appropriate. Width measurements represent maximum width.
Optical microscope images in quoted already published works were acquired using a BH2 light Olympus microscope (Olympus Optical Co., Osachi-shibamiya, Okaya, Nagano, Japan) attached to an AmScope 1000 video camera (United Scope LLC, dba AmScope, Irvine, California), linked to an ASUS laptop equipped with HDMI high-definition multimedia interface system (Taiwan-USA, Fremont, California). Images from the microscope are transferred from the laptop to a USB and stored for subsequent processing on a computer.
Scanning Electron Microscopy (SEM)
Specimens previously processed for SEM were fixed and stored in 70% ethanol following standard methods. These included critical point drying (CPD) (Tousimis Automandri 931.GL) and mounting on aluminum SEM sample mounts (stubs) using conductive double-sided carbon tape. Samples were sputter coated with an 80%-20% gold-palladium target for 3 minutes using a sputter coater (Quorum (Q150T ES) www.quorumtech.com) equipped with a planetary stage, depositing an approximate thickness of 20 nm. Samples were placed and observed in an FEI Helios Dual Beam Nanolab 600 (FEI, Hillsboro, Oregon) Scanning Electron Microscope (FEI, Hillsboro, Oregon). Samples were imaged using an accelerating voltage of 5 kV, and a probe current of 86 pA, at high vacuum using a SE detector.
Deposits
Specimens were deposited in the University of Nebraska’s State Museum’s Harold W. Manter Laboratory (HWML) collection, Lincoln, Nebraska or at the US National Museum (USNM) Helminthological Collection, Beltsville, Maryland. Accession numbers are listed in Table 1.
| Parasite | Host | Distribution | Accession nos. |
|---|---|---|---|
| Class Archiacanthocephala | |||
| Family Moniliformidae | |||
| Moniliformis moniliformis (Bremser, 1811) Travassos, 1915 (cystacanth) | Rattus rattus (Linn.); black rat | San Juan de Miraflores, Lima | Harold W. Manter Lab. (HWML) coll. no. 217622 |
| Class Eoacanthocephala | |||
| Family Neoechinorhynchidae | |||
| Neoechinorhynchus (Neoechinorhynchus) cylindratus peruensis | Gambusia affinis (Baird & Girard); western mosquito fish | Pantanos de Villa Wildlife Refuge, desert coast, Peru | HWML coll. no. 216402 |
| Class Palaeacanthocephala | |||
| Family Echinorhynchidae | |||
| Pseudoacanthocephalus lutzi (Hamann, 1891) | Chaunus limensis Werner; coast toad | Coastal Lima, Peru | HWML nos. 74991- 74993 |
| Family Illiosentidae | |||
| Illiosentis furcatus Van Cleave & Lincicome 1939 | Sciaena deliciosa Tschudi; Lorna drum | La Punta (Callao), Peru | HWML coll. no. 217623 |
| Family Polymorphidae | |||
| Corynosoma australe Johnston, 1937 juveniles | Paralabrax humeralis (Valenciennes); Peruvian rock bass, paratenic host | Chorrillos, Lima, Peru | HWML coll. no. 217624 |
| Polymorphus spindlatus Amin & Heckmann, 1991 (adult) | Nycticorax nycticorax (Linn.); black- crowned night heron | Puno Bay, Lake Titicaca | HWML31700 and USNM 81263 |
| Polymorphus spindlatus Amin & Heckmann, 1991 (cystacanth) | Killifish: Orestias agassi Valenciennes Orestias luteus Valenciennes Orestias mulleri Valenciennes, Orestias olivaceous Garman | Puno Bay & open deeper waters, Lake Titicaca, Peru | US National Mus. (USNM) helminth. coll. no. 84237 & HWML 37935 |
| Polymorphus spindlatus Amin & Heckmann, 1991 (cystacanth) | Oreochromis niloticus Linn.; Nile Tilapia | Villa swamps, Chorrillos, Lima, Peru | Digital photos on a CD |
| Profilicollis altmani (Perry, 1942) Van Cleave, 1947 (adult) | Larus belcheri (Vigors); Belcher’s gull | Ancȯn, Lima, Peru | HWML coll. no. 217625 |
| Profilicollis altmani (Perry, 1942) Van Cleave, 1947 (cystacanth) | Emerita analoga (Stimpson); Pacific sand crab | Chorrillos, Lima, Peru | HWML coll. no. 216672 |
| “Polymorphus (Profilicollis) pacificus” Karl, 1967. Validity questionable | Cerorhinca monocerata (Pallas); Rhinoceros auklet & Aythya (= Marila) affinis (Eyton); lesser scaub | San Francisco, California | US National Mus. (USNM) helminth. coll. |
| “Polymorphus (Profilicollis) pacificus” Karl, 1967. Validity questionable | Larus pipixcan Wangler (adults) & Emerita analoga Stimpson) (cystacanths) | Chorrillos, Lima, Peru | Not deposited. Coll. by Pena; in Amin’s coll. |
| Neoandracantha peruensis Amin & Heckmann, 2017 (cystacanth) | Ocypod gaudichaudii Milne-Edwards & Lucas; ghost crab | Pacific Ocean off Lima | HWML139135 HWML139136 |
Table 1: Acanthocephala recovered from wildlife in Peru and their hosts and geographical distribution.
Results and Discussion
Cystacanths and adults of 8 species of acanthocephalans in 3 classes that we collected from various species of wildlife in Peru over the last few years are presented below. In addition, the status and the validity of Karl’s [8] Polymorphus (= Profilicollis) pacificus including comparative morphometrics are also discussed. Notes on each species are supplemented by light microscope images giving new perspectives even though some have been previously published using line drawings or SEM images. The presented images are novel and original for each species and enhance some of the former depictions of species that may have been previously published.
Moniliformis moniliformis (Bremser, 1811) Travassos, 1915 (cystacanths)
(Moniliformidae) (Figs. 1-3)
Two cystacanths on 2 separate slides labeled # 146, Rodriguez were given to us, probably in 2015 by J. Iannacone. They were collected from Rattus rattus (Linn.) in San Juan de Miraflores, Lima. The specimens (Figs. 1-3) appeared similar to the cystacanths of Moniliformis kalahariensis Meyer, 1931 examined from Meyer’s Berlin Museum für Naturkunde collection with our SEM images in Amin, et al. [9] (Figs. 8-12).
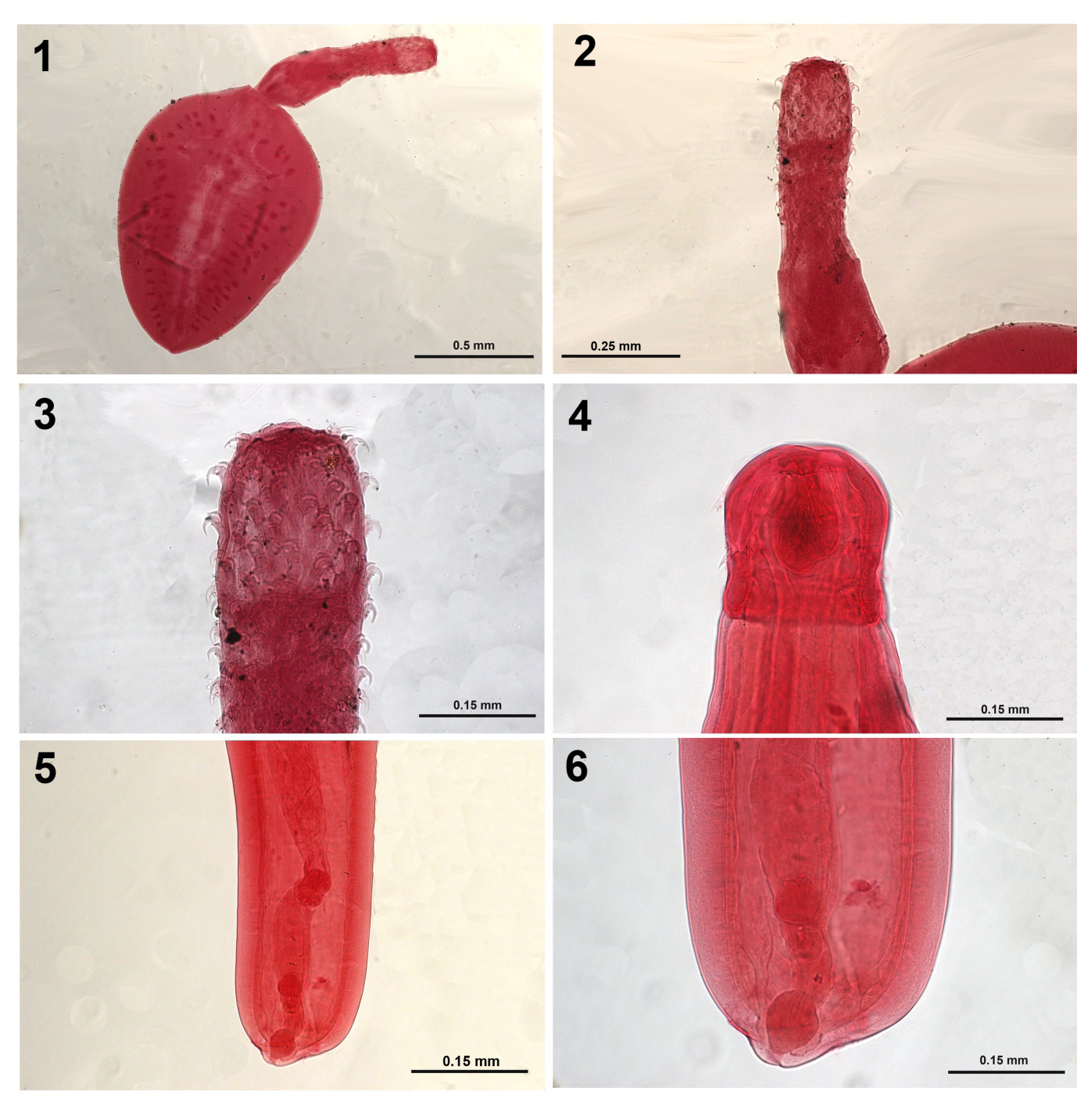
Figures 1-3: Moniliformis monilifo_rmis from _Rattus rattus.
- A whole cystacanth.
- Proboscis and neck.
- Higher magnification of the proboscis showing the shape and arrangement of hooks. Figures 4-6: Neoechinorhynchus (Neoechinorhynchus) cylindratus peruensis n. subsp. from Gambusia affinis.
- The proboscis showing the large apical organ.
- Posterior end of a female specimen showing the reproductive system with a near-terminal gonopore.
- A higher magnification of the posterior portion of the female reproductive system showing the shape of the vaginal bulb, the short uterus, the para-vaginal fibrillar strands, and the near terminal position of the gonopore.
Neoechinorhynchus (Neoechinorhynchus) cylindratus peruensis n. subsp.
(Neoechinorhynchidae) (Figs. 5-6)
Neoechinorhynchus (Neoechinorhynchus) cylindratus peruensis n. subsp., is reported from the mosquitofish Gambusia affinis (Baird & Girard) in Pantanos de Villa. It was first described by Amin, et al. [6] with 4-line drawings and 8 light microscopy images. Amin [10] first created two subgenera in the genus Neoechinorhynchus Stiles and Hassall, 1905 with species in the subgenus Neoechinorhynchus Stiles and Hassall, 1905 being distinguished from species in the subgenus Hebesoma Van Cleave, 1928 by having eggs with concentric shells without polar prolongation of fertilization membrane. In a concept uncommon in the description of species of the Acanthocephala, the species Neoechinorhynchus cylindratus is herein divided into two subspecies. By far, the most common subspecies, neoechinorhynchus is found in marine, freshwater or estuarine fishes, amphibians and reptiles in North America and Mexico. Females are typically with subterminal gonopore at a protruding ventral posterior knob. Females of the other subgenus peruensis have a large double apical organ (Fig. 4) and a near terminal gonopore and no posterior knob (Figs. 5,6), among other differences and is restricted to a small habitat in Peru.
- Designations
- Subspecies neoechinorhynchus n. subsp. With characters of the genus and subgenus Neoechinorhynchus as per Amin [10]. Posterior end of females with sub-ventral gonopore at ventral protruding knob. Common in fish, amphibians and reptiles in North America and Mexico. Type subspecies: Neoechinorhynchus (Neoechinorhynchus) cylindratus cylindratus. The species was originally described from Micropterus salmoides (Lacépède) in Pelican Lake, Minnesota as Neorhynchus cylindratus Van Cleave, 1914 (preoccupied) then Eorhynchus cylindratus Van Cleave, 1914.
• Subspecies peruensis n. subsp. With characters of the genus and subgenus Neoechinorhynchus as per Amin [10] and characters of the species N. cylindratus as per Amin, Peña, Castro [6]. Posterior end of females with terminal to near terminal gonopore and no protruding knobs. Described from imported fish in Pantanos de Villa Wildlife Refuge, Peru. Type subspecies: Neoechinorhynchus (Neoechinorhychus) cylindratus peruensis. Type host: Gambusia affinis (Baird & Girard).
Specimens. Harold W. Manter laboratory (HWML), Lincoln, Nebraska collection no. 216402 (2 males and 2 females on 2 slides).
ZooBank LSID#F4179915-0290-427B-AA2E-FF28F8854203
• Remarks The Peruvian subspecies is further distinguished from the North American and Mexican forms by having considerably wider and more robust lemnisci, paravaginal ligaments, and duct connecting the vaginal bulb with the uterine bell glands (selective apparatus) [6]. The terminal position of the gonopore is a relic of the juvenile developmental stage before the differential development of the dorsal vs. the ventral aspects of the body wall prompting the displacement of the gonopore to a ventral position in the adults which appears to be a function of diet that have become a fixed adult trait in the polluted and degraded environment of the Lima’s Pantanos de Villa Swamp. See Amin, et al. [6] for a detailed discussion.
Pseudoacanthocephalus lutzi (Hamann, 1891)
(Echinorhynchidae) (Figs. 7, 8)
Amin and Heckmann [5] described this acanthocephalan from Peru for the first time based on Iannacone’s donation in 2009 of one vial containing 70% ethanol-preserved specimens used for SEM studies and 7 carmine stained whole mounted specimens on 7 slides collected from the Peru coast toad Chaunus limensis. Specimens were collected in the Lima area, in July, 1987 and January, 1988. Our description included 11 SEM images showing the thick trunk, anterior trunk collar, thin anterior hooks, micropores on hooks and elsewhere in variable diameter and density, sub-ventral female gonopore, and fibrillar coat of the eggs. Pseudoacanthocephalus lutzi was reported but not described from other hosts in different locations in Peru by other observers; see Tantaleán, et al. [11]. Our SEM images [5] of P. lutzi provided good depictions of the external morphology but additional perspectives of internal structures are now revealed by our light microscope images including detail of the proboscis and hooks (Fig. 7) and a subterminal female gonopore (Fig. 8).
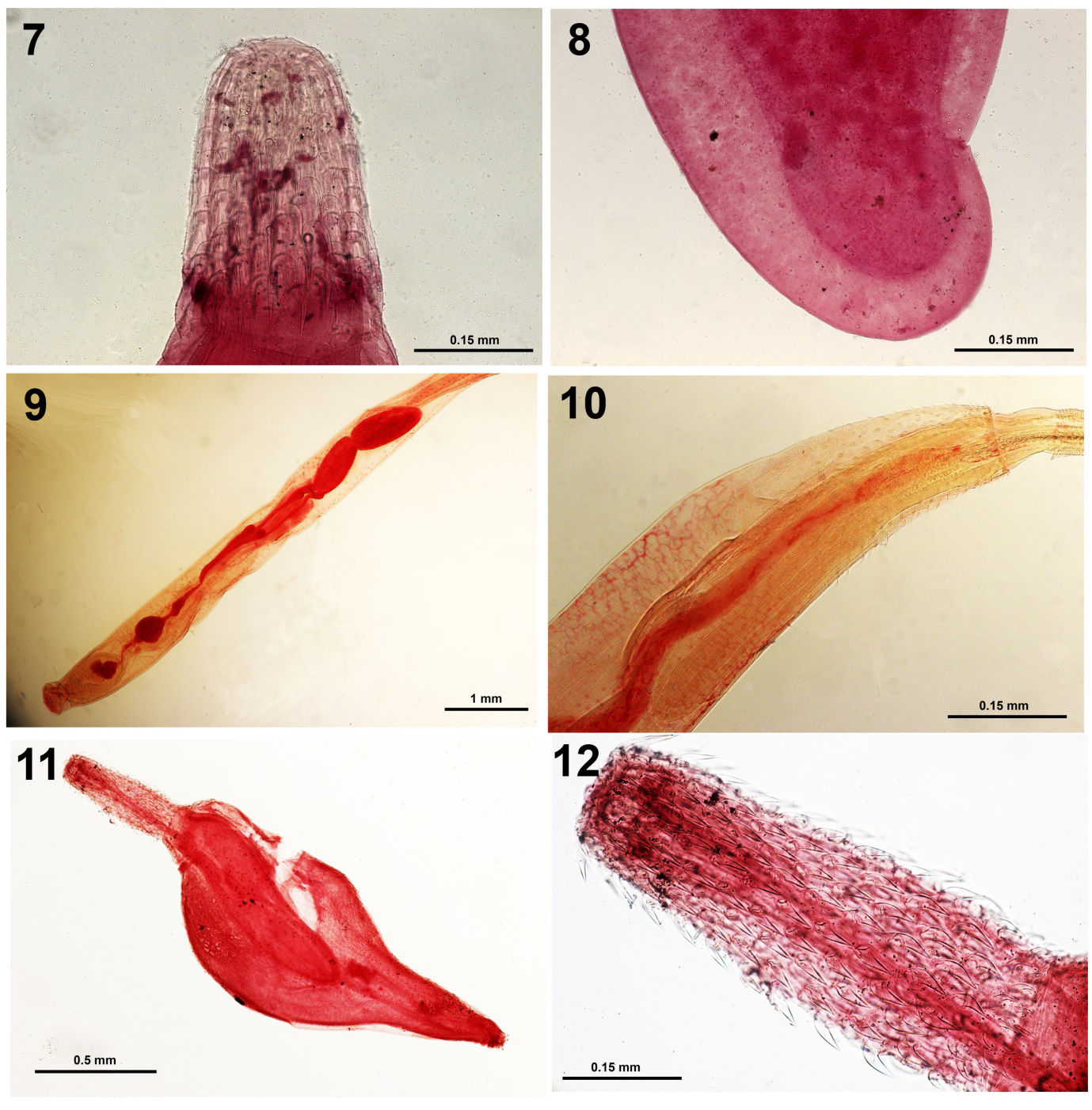
Figure 7-12: Pseudoacanthocephalus lutzi from Chaunus limensis. 7. A proboscis showing the shape and arrangement of hooks. 8. The subterminal gonopore of a female specimen. Figures 9, 10. Illiosentis furcatus from Sciaena deliciosa. 9. The reproductive system of a male specimen marked by the red-stained structures. Note the relative proportions of each. 10. The anterior part of the same specimen in Fig. 9 showing part of the long lemnisci and the unequal distribution of dorsal vs. ventral trunk spines. Figures 11, 12. A juvenile Corynosoma australe from Paralabrax humeralis. 11. A whole organism. 12. An everted proboscis. Note the shape and distribution of hooks.
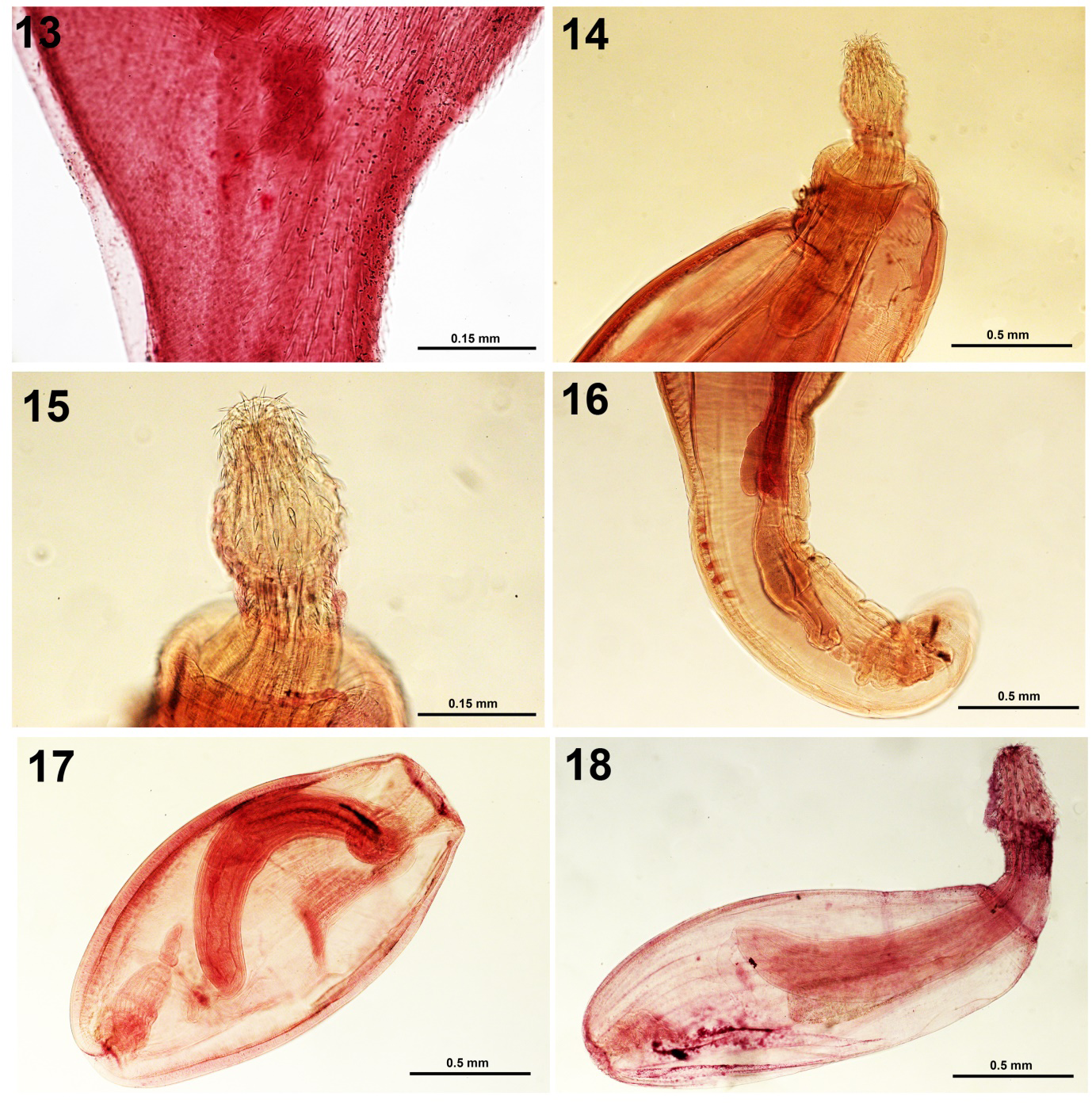
Figures 13-18: 13. A juvenile Corynosoma australe from Paralabrax humeralis. The middle section of a juvenile showing the distribution of the ventral trunk spines. Figures 14-16. Adult Polymorphus spindlatus from Nycticorax nycticorax. 14. Anterior trunk showing the receptacle, strong neck, and characteristic spindle-shaped proboscis. 15. A higher magnification of the proboscis of the same specimen in Fig. 14. 16. The posterior portion of the same male specimen showing the cement glands and body wall nuclei. Figures 17, 18. Polymorphus spindlatus cystacan_ths from _Orestias agassii. 17. A cystacanth with invaginated large receptacle. 18. A cystacanth with a normally evaginated spindle-shaped proboscis comparable to that of the adult.
Illiosentis furcatus Van Cleave & Lincicome 1939
(Family Illiosentidae) (Figs. 20)
Amin, et al. [12] briefly referred to the unusual finding of I. furcatus Van Cleave and Lincicome 1939 from the Lorna drum Sciaena deliciosa Tschudi in Peru for the first time. Illiosentis furcatus was originally described by Van Cleave and Lincicome [13] from the southern kingfish Menticirrhus americanus (Linn.) in the Grand Isle, Louisiana at which time they noted its distribution in the West Atlantic from Cape Cod in Massachusetts, USA to northern Argentina making our finding from Peru a unique extension of its range of distribution. The distribution of the closely related Illiosentis cetratus Van Cleave, 1945 is limited to the Pacific coast of California. The light microscope images of our two specimens of I. furcatus from Peru provided an additional insight into the internal anatomy of these acanthocephalans (Figs. 9,10) that appeared compatible with the original description including the proboscis armature formula that has been corrected in Amin, et al. [13] to 12-15 rows each with 26-33 hooks per row with the presence of genital spines. The closely related I. cetratus has 14 rows of hooks each with 18-23 hooks and genital spines are absent.
Corynosoma australe Johnston, 1937 juveniles
(Polymorphidae) (Figs. 11-13)
Iannacone provided 4 juvenile specimens of Corynosoma sp. collected from the Peruvian rock bass Paralabrax humeralis (Valenciennes) from the Fishing Terminal Chorrillos, Lima in 2009. Later on, Chero, et al. [14] reported it from the corvina drum Cilus gilberti (Abbot) from the same locality as Corynosoma obtuscens Lincicome, 1943 which is a junior synonym of C. australe. Tantalean, et al. [11] reported juvenile C. obtuscens from 18 paratenic hosts in Peru not including P. humeralis and C. gilberti. The most comprehensive study of adult C. australe from sea lions Zalophus californianus (Lesson) by Amin, et al. [15] included morphology including specimens from California, Mexico, Argentina, Antarctica, South Australia, and Brazil, with 32 SEM images and 4 lines drawings, Energy Dispersive X-ray Analysis, and molecular analysis using Cox 1 gene and amplified 18 S ribosomal DNA. The Cox 1 analysis revealed two haplotype networks, one in the Northern Hemisphere (USA and Mexico) and the other in the Southern Hemisphere (Argentina, Brazil, and Peru).
Light microscopy of our 4 peruvian specimens show a whole body (Fig. 11), details of proboscis and hooks (Fig. 12) the distribution of ventral trunk spines (Fig. 13).
Polymorphus spindlatus Amin & Heckmann, 1991 adults & cystacanths
(Polymorphidae) (Figs. 14-16 & 17-18)
Adults of P. spindlatus were collected from two black- crowned night herons Nycticorax Nycticorax (Linn.) shot at Puno Bay, Lake Titicaca in the summer of 1988 [1]. The two birds yielded hundreds of small adult specimens reaching up to 6 mm in length. The specimens were described from 52 whole-mounted worms and distinguished from closely related species especially with their unique spindle-shaped proboscis. The description included 7-line drawings (male, female, proboscis, row of hooks and spines with roots, female reproductive system, eggs), and 4 images of histopathological sections. The proboscis with receptacle, enlarged proboscis, and male reproductive system in adults are depicted in figures 14-16.
We have also studied cystacanths of P. spindlatus from paratenic hosts with the proboscis retracted and everted (Figs. 17, 18) on two separate occasions. Firstly, cystacanths were collected from the liver and intestinal serosa of 64 of 304 individuals (21%) of 4 species of killifish of the genus Orestias collected by gill nets from Lake Titicaca in two locations between 1991 and 1993 (Table 1) [2]. The description was based on 33 whole-mounted specimens with 2-line drawings of a male and female specimens and others sectioned for histopathology with 2 figures. Worms infected liver capsules in hyaline envelopes more frequently than intestinal sites. The pattern of proboscis armature and trunk spines was similar to that of the adults and was distinguished from that of related species. Cystacanths were observed to be not precocious. On another occasion, developing larval stages were obtained from the intestine of the Nile Tilapia Oreochromis niloticus Linn. In Villa Swamps and documented photographically in 8 Figures [4]. The morphology of the fully developed juveniles was comparable to that observed in the Lake Titicaca material, above. The attached light microscopy images of adults and cystacanths provided new perspectives to the line drawings and images in the original descriptions of this species.
Profilicollis altmani (Perry, 1942) Van Cleave, 1947 Adults & Cystacanths
(Polymorphidae) (Figs. 20)
Cystacanths and adults of P. altmani were comprehensively studied from the Pacific mole crab Emerita analoga (Stimpson) and the Belcher’s gull Larus belcheri (Vigors) by Amin, et al. [3]. The energy Dispersive
X-ray analysis of hooks showed unusually high sulfur and diminished phosphorous and calcium levels at hook tips, but higher levels of calcium in middle hooks. Histopathology in L. belcheri showed disruption and destruction of villi, necrotic tissue, and hemorrhaging.
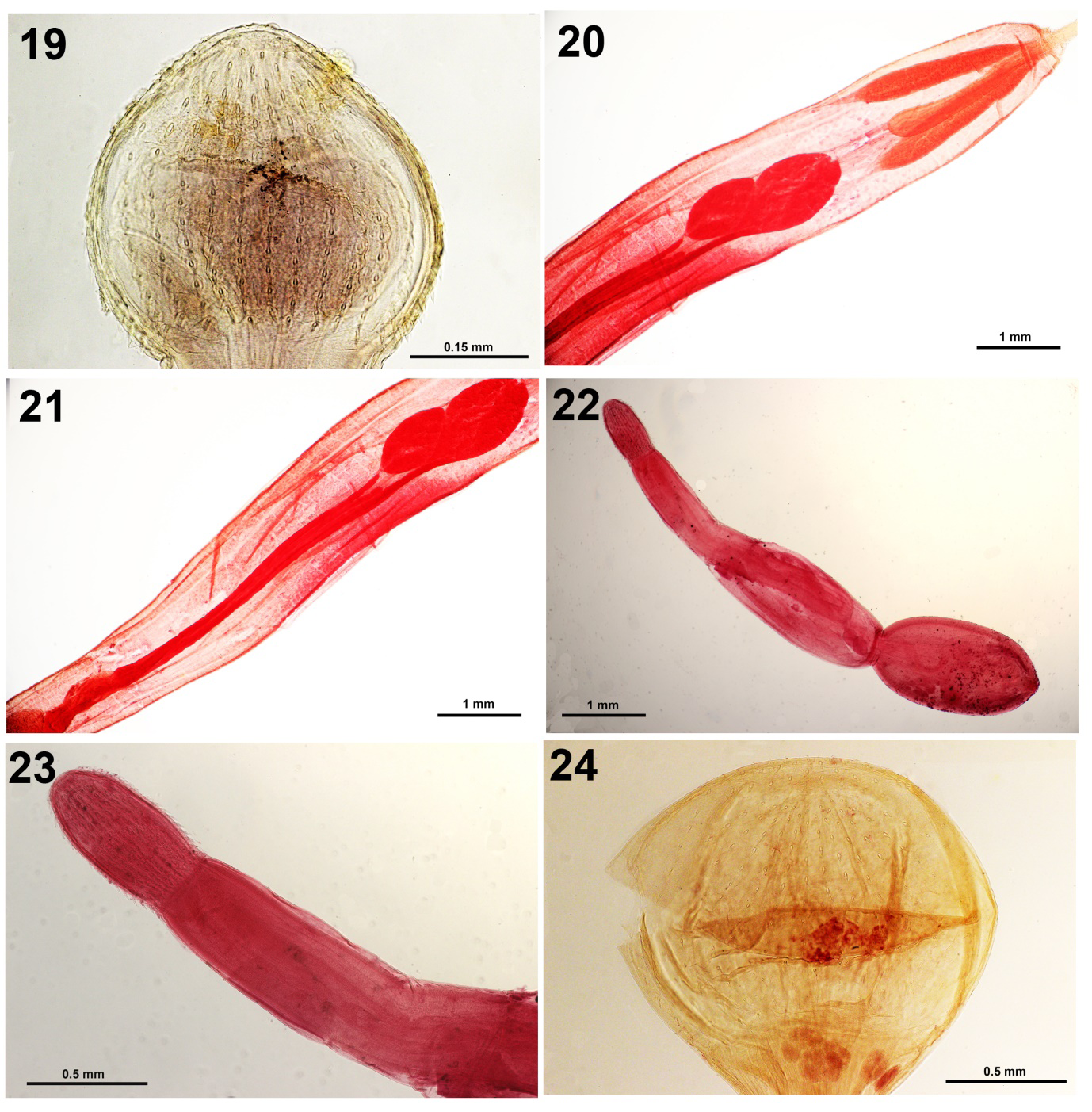
Figure 19-21: Adult Profilicollis altmani from Larus belcheri. 19. A typical proboscis with characteristic shape and armature. 20. Anterior trunk with comparative sizes of receptacle vs. lemnisci, testes interphase and unequal insertion of anterior ends of cement glands. 21. Male reproductive system of the same specimen in Fig. 20 showing the extent of the length of cement glands. Figures 22, 23. Profilicollis altmani cystacanths. 22. A male cystacanth showing the division of trunk regions, the extent of the size of the receptacle and two small testes in the middle section directly posterior to the receptacle. Testes can sometimes be in the posterior trunk. 23. The slightly ovoid proboscis and the elongated neck of the cystacanth in Fig. 22. Figure 24. A typical oblately spheroid proboscis of “Polymorphus (Profilicollis) pacificus” with fewer hooks per row from Larus pipixcan identified by our Peruvian colleagues.
Our recent light microscopy study revealed additional features of the internal anatomy of the proboscis armature (Fig. 19) and male reproductive system (Figs. 20, 21). The morphology of cystacanths (Figs. 22, 23) from various locations in Peru adds to the 34 SEM images produced by Amin, et al. [3] that were compared with those from other populations from the Atlantic and Pacific coasts of North and South America in California, North Carolina, Chile, and Uruguay.
“Polymorphus (= Profilicollis) pacificus Karl, 1967”
(Polymorphidae) (Fig. 24)
Karl [9] described a new species of Profilicollis from California as Polymorphus pacificus Karl, 1967 from two California shore birds (ducks), the rhinoceros auklet Cerorhinca monocerata (Pallas) and the diving duck (lesser scaup) Aythya (= Marila) affinis (Eyton), unlike the gull hosts of P. altmani. Karl [8] found it in mixed infections with P. altmani in C. monocerata and with P. botulus (Van Cleave, 1916) Van Cleave 1939 [13]. Karl [8] distinguished it from populations of P. altmani and its synonyms by the large size of its trunk, oblately spheroidal proboscis (Fig. 24) with 25-30 (28) longitudinal rows of 11-12 hooks each, much- reduced trunk spines, lemnisci longer than receptacle, a pair of cement duct pouches, two independent vaginal sphincters, and larger eggs. The hosts of Karl’s [8] P. pacificus are marine ducks, unlike those of P. altmani and its synonyms.
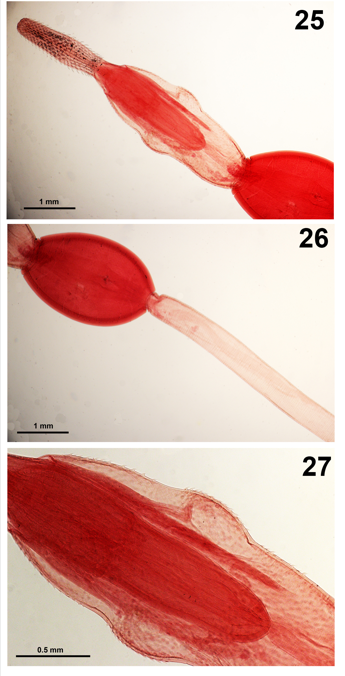
Figures 25-27: Neoandracantha peruensis cystacanths from Ocypod gaudichaudii. 25. The proboscis and anterior trunk section showing its shape with two bulges. Note the comparative size and length of the lemnisci vs. the receptacle. 26. The ovoid posterior trunk and the beginning of the long tail. 27. A higher magnification of the anterior trunk showing the 3 zones of trunk spines in profile.
The trunk of Karl’s [8] P. pacificus is clearly markedly larger than that of P. altmani and its synonyms but the size and oblate proboscis is similar to that of the oblately spheroidal proboscis of the P. bullocki of Mateo, et al. [17] (Table 2). Measurements of the proboscis in the first description by Perry [16] are the smallest. The comparative proboscis hook and spine formulas decidedly overlap. The duct pouches and the distinction in the vaginal sphincters have not been observed by other observers so comparisons cannot be made, and the relatively larger egg size is similar to that of P. bullocki as per Mateo, et al. [17].
| United States | Peru | |||||
|---|---|---|---|---|---|---|
| Study site | Carmel Bay, California | San Francisco, California | Galveston Island, Texas | San Francisco, California | Ancón, Lima | Chancay, Lima |
| Original names | Filicollis altmani | Polymorphus altmani | Polymorphus (Falsificollis) texensis 2 | Polymorphus pacificus | Profilicollis altmani | Polymorphus (Profilicollis) bullocki |
| Authors | Perry (1942) | Perry (1942) emended by Karl (1967)3 | Webster (1948) | Karl (1967) | Amin et al. (2022) | Mateo et al., 1982 |
| Hosts | Melanitta perspicillata & M. deglandi | Catoptrophorus semipalmatus | Crocethia alba | Cerorhinca monocerata & Aythya affinis | Larus belcheri | Larus belcheri |
| Sample size | 1,482 | 10 MM, 10 FF | 2MM, 2FF | 35 of 175 collected | 5 MM, 11 FF | 34 MM, FF |
| Males | ||||||
| Total body L (mm) | ---- | 8.50-16.60 (12.70) | 11.0-15.0 (16.2 in Karl, 1967) | 20.0-25.0 (21.7)3 | 12.75-16.00 (14.50) | 15.60-16.37 (15.86) |
| Trunk L (mm) | 8.5-12.0 (10.5)4 | 9.80-14.97 (11.66) | ---- | 20.88-23.37 (22.52) | 9.37-12.00 (10.47) | ---- |
| Trunk W (mm) | 1.2-2.2 (1.67) | 1.51-3.00 (2.20) | 1.96 in Karl, 1967) | 2.3-3.2 (2.8) | 1.25-2.45 (1.77) | 1.51-1.87 (1.66) |
| Trunk spines no. & L (μm) | Minute, anterior | 25-30 rows each with10-16 spines | 18-27 long (Karl, 1967: in 28-30 rows, 10 spines each) | Reduced, surficial | 30-36 (32) rows of 21-23 (22) spines each; 25-30 (28) μm long | ---- |
| Proboscis shape | Ovoid to oblately spheroidal | Spheroid | Spherical | Oblately spheroidal | Spheroid | Oblately spheroidal |
| Prob. L (mm) | 0.62-0.80 (0.70) | 0.63-1.22 (0.78) | 0.93-0.95 (1.01 in Karl, 1967) | 1.29-1.45 (1.36) | 1.02-1.12 (1.05) | ---- |
| Prob. W (mm) | 0.50-0.96 (0.80) | 0.71-1.23 (0.80) | 1.01-1.14 | 1.5-1.62 (1.59) | 1.20-1.57 (1.41) | 1.50-1.87 (1.66) |
| Hook rows | 25-30 (28) | 20-30 (27) | 22 (24-25 in Karl,1967) | 25-30 (28) | 25-36 (30) | 27-33 |
| Hooks per row | 9-12 (11) | 10-15 (13) | 8-10 (10-11 in Karl, 1967) | 11-12 | 11-13 (12) | 13-15 |
| Apical hook L (μm) | 30-50 (38) | 30-50 (40) | 38 (30-50 in Karl, 1967) | 30-40 (36) | 38-42 (39) | 44 |
| Middle hook L(μm) | 30-60 (47) | 30-60 (50) | 42-51 (40-70 in Karl, 1967) | 30-70 (54) | 37-43 (39) | 32 |
| Basal hook L (μm) | 50-60 (56) | 40-60 (50) | 38 (40-60 in Karl, 1967) | 40-50 (44) | 50-58 (53) | 54 |
| Recept. L (mm) | 1.7-2.75 (2.2) | 1.91-3.51 (2.61) | 3.12-3.56 | 4.55-5.76 (4.81) | 4.12-5.75 (5.28) | 5.74-7.32 (6.17) |
| Recept. W (mm) | ---- | 0.31-0.74 (0.53) | 0.61 (in Karl, 1967) | 0.45-0.57 (0.51) | 0.29-0.50 (0.42) | 0.46-0.53 (0.49) |
| Lemnisci L (mm) | 0.05 shorter than recept. | 1.43-2.40 (1.61); variable | 1.76-2.42 | 2.89-4.00 (3.01); equal | 2.02-2.92 (2.67) | 2.78-3.60 (3.10) |
| Ant. testis L (mm) | Spherical-ovoid | 0.41-1.11 (0.67) | 0.72-1.14 | 0.85-1.01 (0.89); ovoid | 0.85-1.20 (1.05) | 0.96-1.39 (1.20) |
| Ant. testis W (mm) | Spherical-ovoid | 0.39-1.01 (0.56) | 0.84-1.14 | 0.50-0.61 (0.56); ovoid | 0.60-0.80 (0.67) | 0.65-0.96 (0.79) |
| Post. testis L (mm) | Spherical-ovoid | 0.41-1.11 (0.67) | 0.72-1.14 | 0.85-1.01 (0.89); ovoid | 0.87-1.20 (1.05) | 1.03-1.39 (1.25) |
| Post. testis W (mm) | Spherical-ovoid | 0.39-1.01 (0.56) | 0.84-1.14 | 0.50-0.61 (0.56); ovoid | 0.49-0.70 (0.63) | 0.67-0.96 (0.80) |
| Cement gl. No., shape & L (mm) | 4, tubular | 4, tubular 4.00- 5.40 | 4, tubular (4.0-5.4 in Karl, 1967) | 4, tubular 5.01- 5.85 (5.41) | 4, tubular 3.37-5.25 (4.13) | 4, tubular 5.52- 6.50 (5.95) |
| Cement duct pouches | Not observed | Not observed | Not observed | 1 pair | Not observed | Not observed |
| Females | ||||||
| Body L (mm) | ---- | 10.70-18.40 (15.40) | 18 | 22.1-27.1 (24.4) | 10.00-23.00 (15.05) | 16.51-21.60 (19.44) |
| Trunk L (mm) | 10.5-14.0 (12.25) | 9.80-14.17 (11.66) | ---- | 20.88-23.37 (22.52) | 6.87-18.87 (11.27) | ---- |
| Trunk W (mm) | 1.2-2.1 (1.67) | 1.92-2.63 (2.31) | 2.6 (in Karl, 1967) | 2.3-3.2 (2.9) | 1.17-2.20 (1.60) | 1.56-1.99 (1.79) |
| Trunk spines no. & L (μm) | Minute, anterior | 25-30 rows each with 10-16 spines | 18-27 long (Karl, 1967: in 26-28 rows, 10 spines each) | Reduced, surficial | 38-54 (47) rows of 23-34 (28) spines each; 22-30 (26) μm long | ---- |
| Proboscis shape | Ovoid to oblately spheroidal | Spheroid | Spherical | Oblately spheroidal | Spheroid | Oblately spheroidal |
| Prob. L (mm) | 0.62-0.80 (0.70) | 0.63-1.22 (0.78) | 1.20-1.23 | 1.29-1.45 (1.36) | 0.67-1.37 (0.96) | ---- |
| Prob. W (mm) | 0.50-0.96 (0.80) | 0.71-1.23 (0.80) | 1.23 | 1.5-1.62 (1.59) | 0.75-1.57 (1.13) | 1.54-1.91 (1.71) |
| Hook rows | 25-30 (28) | 23-30 (27) | 22-24 (24-25 in Karl, 1967) | 25-30 (28) | 30-34 (32) | 27-33 |
| Hooks per row | 9-12 (11) | 10-15 (13) | 8-10 (10-11 in Karl, 1967) | 11-12 | 11-16 (13) | 13-15 (usually 15) |
| Apical hook L (μm) | 30-50 (38) | 30-50 (40) | 33 (30-50 in Karl, 1967) | 30-40 (36) | 34-45 (38) | 45 |
| Middle hook L (μm) | 30-60 (47) | 30-60 (50) | 51 (40-60 in Karl, 1967) | 30-70 (54) | 38-42 (39) | 35 |
| Basal hook L (μm) | 50-70 (56) | 40-60 (50) | 33 (40-60 in Karl, 1967) | 40-50 (44) | 48-60 (55) | 56 |
| Recept. L (mm) | 1.70-2.75 (2.20) | 1.91-3.51 (2.61) | 2.95-3.12 (3.22 in Karl, 1967) | 4.55-5.76 (4.81) | 4.00-6.50 (5.54) | 6.48-6.72 (6.60) |
| Recept. W (mm) | ---- | 0.31-0.74 (0.53) | (0.62 in Karl, 1967) | 0.45-0.57 (0.51) | 0.20-0.55 (0.34) | 0.48-0.55 (0.52) |
| Lemnisci L (mm) | 0.05 shorter than recept. | 1.43-2.40 (1.61); variable | 1.65-1.98 (1.98 in Karl, 1967) | 2.89-4.00 (3.01); equal | 1.75-3.75 (2.60) | 3.00-3.89 (3.46) |
| Vaginal sphincters | ---- | ---- | ---- | Separate | ---- | ---- |
| Egg length (μm) | 60-70 (75) | 60-70 (65) | 58-73 | 65-81 (74) | 52-60 (57) | 66-76 (71) |
| Egg width (μm) | 22-30 (26) | 22-30 (26) | 18-22 | 25-33 (29) | 16-23 (19) | 24-38 (28) |
Table 2: Comparative morphometrics between populations and synonymies of _Profilicollis altmani_ (Perry, 1942) Van Cleave, 1947 (
Table 2: Comparative morphometrics between populations and synonymies of Profilicollis altmani (Perry, 1942) Van Cleave, 1947 (=P. texensis & P. bullocki)1 and Polymorphus (=Profilicollis) pacificus of Karl, 1967. 1. Other synonyms include P. kenti Van Cleave, 1947. 2. Polymorphus texensis has a wide range of measurements and is included to give a good measure of intraspecific variability within Profilicollis altmani. Results of Karl’s (1967) re-examination of Webster’s type material are added between parentheses. 3. Karl (1967) reported 8 species of California shore birds naturally infected with P. altmani, with Catoptrophorus semipalmatus being the most heavily infected species. His morphometrics (in this table) were based on emended description of specimens obtained from his heavily infected experimental hosts, the duckling Anas platyrhynchus Linn. Given common measurements of proboscis, hooks, receptacle, and lemnisci were the same for both sexes. 4. Range (mean). 5. Bolded figures are exceptional extremes.
Karl’s [8] P. pacificus has not been recognized since. We are concerned about this species because certain of our Peruvian collaborators labeled some of their slides of adults as Polymorphus (Profilicollis) pacificum from Larus pipixcan Wangler, from Lima in 1976. Some of the slides of cystacanths collected from Emerita analoga by the same workers were also labeled Polymorphus (Profilicollis) pacificum. We have critically examined these whole-mounted smaller adults from Peru and light microscopical images of some of their features were compared with those described by Karl [8]. We have observed some comparability especially in the proboscis shape, size and armature (fewer hooks per row) to support Karl’s [8] description that appears, however, to depict an extreme variant of P. altmani as measurements and counts of critical structures appear to merge or overlap (Table 2) but perhaps not sufficient to confer a new species status.
Neoandracantha peruensis Amin & Heckmann, 2017 cystacanths
(Polymorphidae) (Figs. 25-27)
The cystacanths of Neoandracantha peruensis Amin & Heckmann [7] were described from the ghost crab Ocypod gaudichaudii Milne-Edwards & Lucas from the Pacific coast off Lima, Peru. The new genus Neoandracantha Amin and Heckmann [7] has two anterior swellings (Figs. 25,26) with 3 separate fields of spines on the fore-trunk swelling (Fig. 27) which distinguished it from Andracantha Schmidt, 1975 that has only two fields of anterior trunk spines. Our specimens are slender with long tubular hind trunk (Fig. 26) void of genital spines. Our original description was supplemented with 6-line drawings, 17 SEM images, and an Energy Dispersive X-ray analysis (EDXA). The EDXA showed very low sulfur levels and high phosphorous and calcium levels in proboscis hooks and the opposite pattern was observed in trunk spines [7].
Acknowledgements
We gratefully appreciate the contributions of all colleagues and associates who have helped with providing the research materials acknowledged in the noted and referenced texts especially in relation to field collections and sharing specimens. This work could not have been completed without their support.
Conflict of Interest
The authors declare no conflicts of interest.
Ethical Approval
The authors declare that they complied with all relevant standards.
References
-
Amin OM, Heckmann HA (1991) Description and host relationships of _Polymorphus splindlatus_ n. sp. (Acanthocephala: Polymorphidae) from the heron, _Nycticorax nycticorax_ in Peru. J Parasitol 77(2): 201-205.
-
Amin OM, Heckmann RA, Mesa R, Mesa E (1995) Description and host relationships of cystacanths of _Polymorphus_ _spindlatus_ (Acanthocephala: Polymorphidae) from their paratenic fish hosts in Peru. J Helminthol Soc Wash 62(2): 249-253.
-
Amin OM, Rodríguez SM, Rubtsova N, Heckmann RA, Peña C, et al. (2022) A comparative assessment of the morphology of _Profilicollis_ _altmani_ (Acanthocephala, Polymorphidae) from crustaceans and shore birds in Peru, with special notes on hook elemental analysis (EDXA), SEM imaging, histopathology, and molecular profile. Parasite 29: 9.
-
Amin, OM, Heckmann RA, Peña C, Castro T (2010) On the larval stages of _Polymorphus spindlatus_ (Acanthocephala: Polymorphidae) from a new fish host, _Oreochromis_ _niloticus,_ in Peru. Neotrop Helminthol 4: 81-85.
-
Amin OM, Heckmann RA (2014) First description of _Pseudoacanthocephalus lutzi_ from Peru using SEM. Sci Parasitol 15: 19-26.
-
Amin OM, Peña C, Castro T (2021) An unusual type of _Neoechinorhynchus cylindratus_ (Acanthocephala: Neoechinorhynchidae) from _Gambusia affinis_ in Peru, with notes on introductions. Acta Parasitol.
-
Amin OM, Heckmann RA (2017) _Neoandracantha_ _peruensis_ n. gen., n. sp. (Acanthocephala: Polymorphidae) described from cystacanths infecting the ghost crab _ocypode guadichaudi_ in the peruvian coast. Parasite 24: 40.
-
Karl JE (1967) Studies on the systematics and life history of _Polymorphus altmani_ (Perry). Ph.D. Dissertation. Baton Rouge, Louisiana: Louisiana State University pp: 132.
-
Amin OM, Heckmann RA, Halajian A, El-Naggar A, Takavol S (2014) Description of _Moniliformis kalahariensis_ (Acanthocephala: Moniliformidae) from the South African hedgehog _Atelerix frontalis_ (Erinaceidae) in South Africa. Comp Parasitol 81: 33-43.
-
Amin OM (2002) Revision of _Neoechinorhynchus_ Stiles and Hassall, 1905 (Acanthocephala: Neoechinorhynchidae) with keys to 88 species in two subgenera. Syst Parasitol 53: 1-18.
-
Tantaleán M, Sánchez L, Gómez L, Huiza A (2005) Acanthocephalan from Peru. Rev Peru Biol 12(1): 83-92.
-
Amin OM, Chaudhary A, Singh HS (2023a) Redescription of _Illiosentis cetratus_ Van Cleave, 1945 (Acanthocephala: Illiosentidae) from _Menticirrhus undulatus_ (Girard) in California, with notes on _Illiosentis furcatus_ from Peru. Folia Parasitol 70: 018.
-
Van Cleave HJ, Lincicome DR (1939) On a new genus and species of Rhadinorhynchidae (Acanthocephala). Parasitology 31: 413-416.
-
Chero J, Cruces C, Sáez G, Alvariño L (2014) Drum _Cilus_ _gilberti_ (Abbott, 1899) (Perciformes: Sciaenidae) in the coastal zone of Chorrillos, Lima, Peru. Neotropical Helminthology 8: 163-182.
-
Amin OM, Chaudhary A, Singh HS, Kuzmina T (2023b) Revision of _Corynosoma australe_ Johnston, 1937 (Acanthocephala: Polymorphidae) from a North American population using novel SEM images, Energy Dispersive X-ray analysis and molecular analysis. Helminthologia 60(1): 1-27.
-
Perry ML (1942) A new species of the acanthocephalan _Filicollis_. Journal of Parasitology 28: 385-388.
-
Mateo E, Cordova R, Guzmán E (1982) _Polymorphus_ (_Profilicollis_) _bullocki._ Nueva especie de acantocéfalo hallado en la gaviota _Larus belcheri_ en el Perú. Boletin de Lima 4: 73-78.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind