Redescription of Pseudobenedenia lauriei Szidat 1965 (Monogenea: Capsalidae) from Notothenia neglecta in Laurie, Peterman, and South Orkney Islands, South Antarctica
While reviewing museum collections of Pseudobenedenia Johnston, 1931, we found five deposited slides marked by Dr. Lothar Carlos Guillermo Szidat as Pseudobenedenia lauriei Szidat, 1965 from Notothenia neglecta Nybelin, 1951. We have removed the synonymization of Pseudobenedenia lauriei Szidat, 1965 as a junior synonym of Pseudobenedenia nototheniae Johnston, 1931 and restored its identity as a valid species that we redescribe herein. We added never-published data on the ovary, testes, bursa, haptor diameter, and egg of P. lauriei. As well, we added data on body size, measurements of sclerotized structures of haptor (accessory sclerites, anterior and posterior hamuli), oral suckers, pharynx and anterior hamulus to accessory sclerite ratio. A comparison of the measurements the presently recognized species of P. lauriei shows that it differs from all known species of Pseudobenedenia Johnston, 1931 and corresponds to the data of its original description. We provide photomicrographs of P. lauriei for the first time in addition to more informative line drawings of this species.
Nataliya Y Rubtsova1* and Paula Marcotegui2
Keywords: Pseudobenedenia lauriei; Redescription; Notothenia neglecta; South Antarctica
Introduction
The first species of Pseudobenedeniae – Pseudobenedenia nototheniae Johnston, 1931 was described in 1931 and 1937 [1, 2] from the body surface of Notothenia microlepidota Hutton, 1875 and Paranotothenia magellanica (Forster, 1801). In 1965, Szidat described Pseudobenedenia lauriei Szidat, 1965 from the pectoral fins and body surface of Notothenia neglecta Nybelin, 1951 [3]. No type material was designated by Szidat [3] and information on the number of specimens used for the description was missing [3]. In 1968 Hargis, et al. [4] described Pseudobenedenia shorti Hargis et Dillon, 1968 from nototheniid fish of the genus Trematomus [4] and Yamaguti [5] described Pseudobenedenia elongate Yamaguti, 1968, Pseudobenedenia merinthe Yamaguti, 1968 and Pseudobenedenia ovalis Yamaguti, 1968 from Hawaiian fish [5]. In 1976, Gibson [6] revised the genus Pseudobenedenia and transferred P. shorti to the genus Pseudobenebenoides Szidat, 1969 [6, 7]. He established the new genus Parabenedenia Gibson, 1976 and transferred to it Pseudobenedenia nototheniae, Parabenedenia noblei [8], Pseudobenedenia elongate, Pseudobenedenia merinthe and Pseudobenedenia ovalis. The same year Gibson corrected himself by replacing the name of the genus Parabenedenia with Menziesia Gibson, 1976, and transferring to this genus P. noblei, P. elongate, P. merinthe, and P. ovalis. He also transferred Pseudobenedenia nototheniae back to the genus Pseudobenedenia Johnston [9]. Gibson [6] also synonymized Pseudobenedenia lauriei with Pseudobenedenia nototheniae without studying any collections of this species. Later on, Timofeeva, et al. [10] described Pseudobenedenia gibberifrons Timofeeva, Gaevskaja, Kovaleva, 1987 from Gobinotothen gibberifrons (Lönnberg, 1905) and Pseudobenedenia dissostichii Timofeeva, Gaevskaja, Kovaleva, 1987 from Dissostichus eleginoides Smitt, 1898. More recently, a new species Pseudobenedenia coriicepsi Rubtsova, Chaudhary, Salganskiy, Kuzmina, 2023 was described from Notothenia coriiceps [11].
This work aimed to redescribe P. lauriei using museum collections of P. lauriei from the Argentine Museum of Natural Sciences Bernardino Rivadavia, Buenos Aires, Argentina, collected by Szidat, Bellizio in 1963 and Szidat and Groussac in 1968. We intended to provide additional information on this species, designate lectotype and paralectotypes, and establish that P. lauriei is a valid species.
Materials and Methods
Our study based on five whole-mounted museum specimens, fixed in Canada balsam. Measurements and microphotographs were made in the Ichthyoparasitology Laboratory, Institute of Marine and Coastal Research (IIMyC), Faculty of Exact and Natural Sciences, National University of Mar del Plata, Argentina, with microscope Leica LED2500, x10, x40 magnification). Microphotographs made by Leica DFC 295 camera. Line drawing and analysis of data made in the Institute of Parasitic Diseases (IPD) Scottsdale, AZ, USA.
Results
The Argentine collection at hand included two specimens collected in 1963 by Szidat and Bellizio near Laurie Island. These two specimens possibly were used for the original description of P. lauriei published two years later. Szidat and Groussac collected three other specimens of P. lauriei in 1968 in other locations – Peterson and South Orkney Islands. All this information we revealed by studying hand-written inscriptions on labels of the museum specimens.
In Table 1, we present the results of our measurements of five specimens of P. lauriei from N. neglecta – two collected by Dr. Lothar Szidat together with Dr. N. Bellizio (full name of researcher unknown) near Lauriei Island in 1963 and four collected by Szidat and Groussac (full name unknown) in 1968 at Peterman and South Orkney Islands (Table 1). For comparison, we added existing measurements of the type species of genus Pseudobenedenia (P. nototheniae) and a few other species described from other nototheniid fish from Antarctica. We excluded from this table records that we consider to represent mixed data from a few hosts that were measured together or misidentifications (except for P. nototheniae, that we consider a species complex [11].
| Host | Paranotothenia magellanica (Forster 1801), Notothenia microlepidota Hutton 1875 | Notothenia neglecta Nybelin 1951 | Notothenia neglecta Nybelin 1951 | Gobionotothen gibberifrons (Lönnberg 1905) | Dissostichus eleginoides Smitt 1898 | Notothenia coriiceps Richardson 1844 |
|---|---|---|---|---|---|---|
| Parasite | Pseudobenedenia nototheniae* | Pseudobenedenia lauriei | Pseudobenedenia lauriei | Pseudobenedenia gibberifrons | Pseudobenedenia dissostichii | Pseudobenedenia coriicepsi |
| Authority | Johnston, 1931 Johnston, 1937 | Szidat, 1965 | Our measurements from museum whole mounts deposited by Szidat, Bellizio 1963 and Szidat, Groussac 1968 | Timofeeva et al. 1987 | Timofeeva et al. 1987 | Rubtsova et al. 2023 |
| Locality | Sub-Antarctic islands of New Zealand, Macquarie Isl. | Scotia Bay, Laurie Isl., South Orkney Isl. | Laurie Isl, Peterman Isl, South Orkney Isl. | Georgiy Isl., Mordvinov Isl. | Georgiy Isl., Mordvinov Isl. | Galindez Isl. |
| Site of infection | Body surface | Pectoral fin, body surface | Pectoral fin, body surface | Pectoral fins | Body surface | Body surface |
| Body length | 4.7–7.0 | 4.5 | 4.06 (3.56-4.26) | 5.6 (4.7-6.2) | 8.7 (7.1-10.8) | 6.03 (4.75–7.0) |
| Body width | 2.3–4.0 | 2.5 | 2.31(2.19-2.46) | 2.6 (2.1-3.7) | 3.9 (3.3-4.8) | 2.73 (2.0–3.70) |
| Oral suckers diameter | 0.8 | 0.4 | 0.57 (0.45-0.71) × 0.53 (0.35- 0.67) | 0.76 (0.67-0.88) | 1.04 (0.81-1.24) | 0.45 (0.33–0.52) × 0.48 (0.44–0.52) |
| Pharynx | 0.5 × 0.6 | 0.3 | 0.31 (0.27-0.34) × 0.45 (0.40- 0.53) | 0.59(0.46-0.62) × 0.70(0.48-0.80) | 0.88 (0.67-1.12) ×1.07 (0.75-1.28) | 0.46 (0.33–0.6) × 0.48 (0.35–0.7) |
| Ovary | 0.3 × 0.5–0.7 | n/a | 0.58 (0.48-0.73) × 0.43 (0.34- 0.51) | 0.48(0.37-0.55) ×0.67(0.43-0.75) | 0.76 (0.67-0.80) ×1.05 (0.91-1.15) | 0.47 (0.38–0.55) × 0.50 (0.44–0.6) |
| Testes | 1.2–1.6 × 0.8–1.0 | n/a | 0.34 (0.29-0.45) × 0.47 (0.41- 0.51) | 0.68(0.58-0.75) ×0.61(0.44-0.75) | 1.62 (1.38-1.95) × 0.91 (0.86-1.38) | 0.91 (0.73–1.04) × 0.60 (0.49–0.78) |
| Bursa | 0.8 × 0.3 | n/a | 0.52 (0.40-0.67) | 0.53(0.48-0.64) ×0.22(0.19-0.24) | 1.16 (0.98-1.39) × 0.34 (0.26-0.37) | 0.55 (0.50–0.59) |
| Haptor diameter | 2.0–2.4 | n/a | 1.56 (1.41-1.77) | 2.04(1.85-2.38) | 2.78 (2.46-3.40) | 1.44 (1.14–1.57) × 1.57 (1.40–1.75) |
| Anterior hamulus length | 0.38–0.53 | 0.4 | 0.46 (0.38-0.55) | 0.58(0.55-0.64) | 0.80 (0.72-0.86) | 0.51 (0.41–0.60) |
| Posterior hamulus | 0.13–0.17 | 0.1 | 0.22 (0.17-0.25) | 0.27(0.22-0.32) | 0.26 (0.21-0.32) | 0.16 (0.13–0.23) |
| Accessory sclerite | 0.23 | 0.28 | 0.27 (0.20-0.36) | 0.39(0.33-0.45) | 0.53 (0.44-0.59) | 0.38 (0.24–0.46) |
| Egg | 0.2 × 0.1 | n/a | 0.17 (0.21-0.22) × 0.13 (0.11- 0.15) | 0.24(0.22-0.27) ×0.14(0.10-0.15) | 0.22 (0.19-0.24) × 0.14 (0.10-0.16) | 0.31 (0.31–0.31) × 0.13 (0.13–0.14) |
Table 1: Comparative morphometric data on Pseudobenedenia species from its different specific hosts.
*Type species of the genus; we consider it a species complex [11]. Table 1: Comparative morphometric data on Pseudobenedenia species from its different specific hosts.
Redescription of Pseudobenedenia lauriei Szidat, 1965 (Figures 1 & 2, Table 1).
Type host: Notothenia neglecta Nybelin, 1951. Site of infection: pectoral fin, body surface. Type locality: coastal waters of Lauriei (60028’S, 43084’W) and South Orkney Islands (60047’S, 45055’W); other locality: Peterman Island 64093’S, 65005’W (West Antarctica). Material studied for redescription: five Canada balsam mounted specimens.
Designated specimens: lectotype (27.977 d) and paralectotypes (27.977 a, b, c, e), Argentine Museum of Natural Sciences Bernardino Rivadavia, Buenos Aires, Argentina.
Based on five mature egg-bearing specimens and the original description information. Information below in mm, average (min-max). In square brackets we provided data from the original description of P. lauriei, N/A – data in the original description were not available: body length 4.06 (3.56-4.26) [4.5], body width at the widest part 2.31 (2.19-
2.46) [2.5] (Figures 1 & 2, Table 1). Prohaptor with two lobes and slight [to deep] cleft (Figures 1 & 2). Oral suckers well developed, occupying most prohaptor and part of trunk 0.57 (0.45-0.71) x 0.53(0.35-067) [0.4]. Pharynx 0.31 (0.27- 0.34) x 0.45(0.40-0.53) [0.3]. Intestine branches obscured. Bursa 0.52 (0.40-0.67) [N/A] opens on left ventral side of the body, under middle part of the left oral sucker. Ovary central 0.58(0.48-0.73) x 0.43 (0.34-0.51) [N/A]. Genital opening not visible [less to the left in comparison with P. nototheniae]. Two testes 0.34 (029-0.45) x 0.47 (0.47 (0.41-
0.51) [N/A] posterior to ovary, longitudinal oval, side-by- side, not lobulated, occasionally with pointed upper part adjacent to ovary, with moderate perforations. Single egg 0.17 (0.21-0.22) x 0.13 (0.11-0.15) [N/A], with long coiled filament. Haptor 1.56 (1.41-1.77) [N/A] divided by six septa. Sclerotized parts of haptor in three pairs of structures: accessory sclerites 0.27 90.20-0.36 [0.28] closer to central part of haptor, and anterior hamuli 0.46 (0.38-0.55) [0.4] at distal part of haptor with adjacent posterior hamuli 0.22 (0.17-0.25) [0.1]. Marginal hooks inconspicuous.
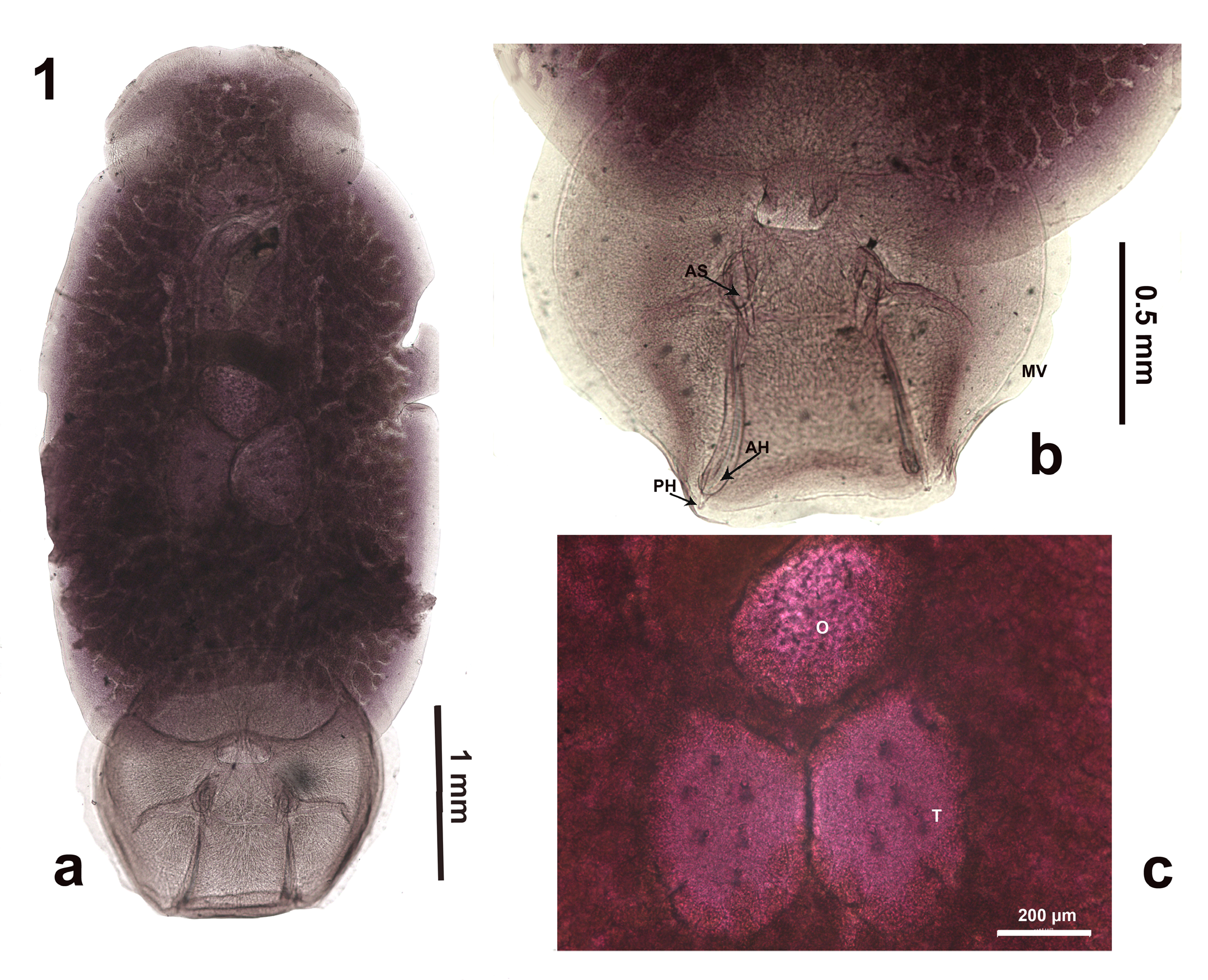
Figure 1: Total view microphotography of Pseudobenedenia lauriei Szidat 1965. Note slightly pointed shape of ovaries and well-seen septae and commissures of haptor; 1b: enlarged haptor of other specimen of P. lauriei, MV – marginal valve, AS – accessory sclerite, AH – anterior hamulus, PH – posterior hamulus; 1c: close view ovary (O) and testes (T) complex of the other specimen of P. lauriei.
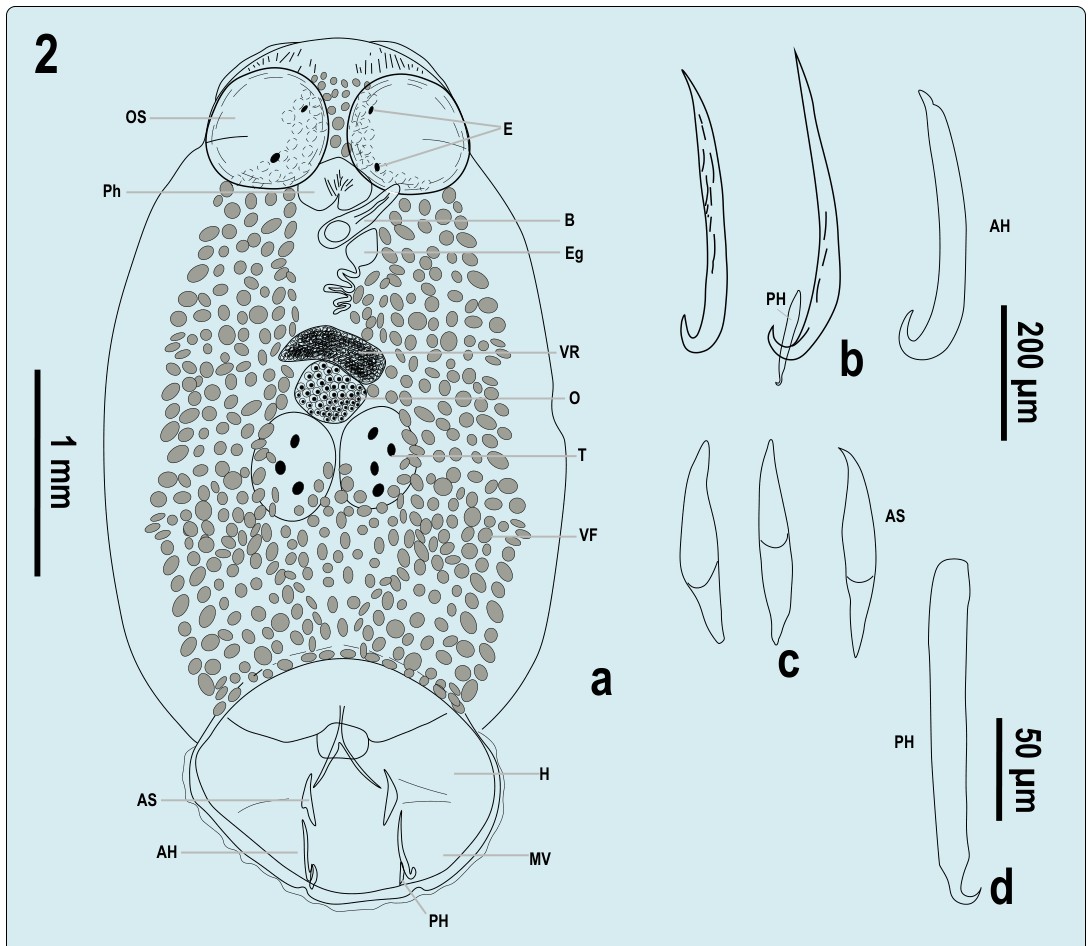
Figure 2: Total view of Pseudobenedenia lauriei Szidat 1965: E – eyes, OS – oral sucker, Ph– pharynx, B – bursa, Eg – egg, VR – vitelline reservoir, O – ovary, T – testes, VF –vitellaria filed, H – haptor, MV – marginal valve, AS – accessory sclerite, AH – anterior hamulus, PH – posterior hamulus. 2b: AH – anterior hamuli (not in profile, from three different specimens, PH – posterior hamulus in its natural adjacent position to anterior hamulus; 2c: accessory sclerites; 2d posterior hamulus, in profile.
Remarks
Because the description of P. lauriei lacked detailed graphic information, we provided additional information extracted from the museum collection. Fig. 1 shows a light microscopy photograph of P. lauriei. Fig 2 shows a line drawing based on all available whole mounts (Fig. 2a). We made drawings of sclerotized parts of haptor (Fig. 2 B-C). As for anterior hamuli (2B), none of them was perfectly positioned in profile, which was impossible to correct in Canada balsam, we found informative accessory sclerites (2 C) and posterior hamulus (2 D).
Differential Diagnosis
Pseudobenedenia lauriei differs from P. nototheniae by smaller oral suckers diameter, pharynx, testes, and haptor diameter, by shape of the anterior hamulus (Table 1), anterior hamulus to accessory sclerite ratio 1.7 (versus 2.1 in P. nototheniae) and by the host species. We consider P.
nototheniae a species complex, so this comparison is relative.
Pseudobenedenia lauriei differs from P. coriicepsi by smaller body length and width, larger diameter of oral suckers and ovary, smaller diameter of pharynx, anterior hamulus, and accessory sclerite (Table 1), anterior hamulus to accessory sclerite ratio 1.7 (versus 1.4 in P. coriicepsi) [11] and by type of host.
Pseudobenedenia lauriei differs from P. gibberifrons and from P. dissostichii by all smaller metric parameters, and by different hosts (Table 1).
Discussion
Some specimens from the museum that we studied were in a very poor state, but we tried to measure them as precisely as possible. There was a sixth specimen marked as P. lauriei from N. neglecta in the museum collection that was completely unsuitable for research. If we took into account only two specimens collected in 1963 by Szidat and Bellizio, they still differed from the published material by Szidat in 1965 in the original description. The diameter of pharynx differs (0.5 and 0.46 from 0.3 in the original description as well as accessory sclerite and anterior hamuli are slightly longer than in the description of P. lauriei [3], while other measurements correspond to those published in the description). This indicates that the original description was based on more than these two specimens collected in 1963 or (less likely) that the author did not use these specimens for the description at all. What matters is that they were collected by the author in the same location and the same host and were never designated for this species deposited in the museum until the present.
According to our data, we can see that the original measurements of P. lauriei lay in the range of our measurements of the P. lauriei collected by the author [3]. The original description was missing data on the ovary, testes, bursa, haptor diameter, and egg. The only parameter that was out of the range is the length of the tiny sclerotized structure - the posterior hamulus that was 0.1 in Szidat`s description [3] and 0.22 in ours. Therefore, by the metric parameters and by host specificity our data and Szidat’s description form the harmonic unity. Despite the somewhat poor quality of these 60-year-old specimens, we tried to create a more detailed drawing of the whole view of P. lauriei together with more detailed images of sclerotized structures of haptor (Fig. 2, b-d). In the studied whole mounts, sclerotized structures never appeared perfectly in profile, as needed for taxonomical studies, and extremely difficult to achieve while working with such big monogeneans as Capsalidae are [11].
Our results suggest that synonymizing P. lauriei with P. nototheniae, [6] was in error. Intuitively Gibson [6] may have detected that something with his synonymy was not right. He wrote about the surprisingly low degree of host-specificity of P. nototheniae, which markedly contrasts with the related Entobdella solae Kearn, 1967, which has strict host specificity. He also had the right notion that there is something wrong with such a wide range of hosts, emphasizing that P. nototheniae being a skin parasite should be susceptible to any immune factors present in the mucus of its host [6].
The host of P. lauriei, Notothenia neglecta Nybelin, 1951 is a valid fish species, the closest sister species to Notothenia coriiceps Richardson, 1844 [12, 13]. Notothenia neglecta is spread along the coast of the Antarctic continent, Antarctic Peninsula, South Georgia, South Shetland, South Orkney Islands, and Peter I Island, according to Fish Base [14].
While studying Capsalids from twenty-three species of notothenoid fish, including fourteen representatives of family Nototheniidae, over few years in Atlantic part of Antarctica, Timofeeva, et al. [10] have formulated few patterns that are common for these monogeneans. All Pseudobenedeniae specimens collected from one species of host were different from ones from other fish species by body size, size of haptor, sizes of its sclerotized attachment structures of haptor and by their shape, sizes and relative position of sexual organs [10]. The localization on the body of the individual hosts was often different, and there was a difference in parameters of infection [10]. That corresponds with our published data [11] and our data that are in press. Thus, different nototheniid fish harbor close but different species of Pseudobenedenia, as well as two other genera of Pseudobenedeniella Timofeeva, Gaevskaja, Kovaleva, 1987 and Pseudobenedenoides Szidat, 1969 [1, 7, 10]
Acknowledgments
Our gratitude goes to Alejandro Tablado, Argentine Museum of Natural Sciences Bernardino Rivadavia, Buenos Aires, Argentina, for his help with museum collections.
References
-
Johnston TH (1931) New Trematodes from the Subantarctic and Antarctic. Austr J Experim Biol Medical Sci 8(2): 91-98.
-
Johnston TH (1937) Trematoda. Australasian Antarctic Expedition 1911-1914. Scient Rep Ser C Zool Bot 10(1): 1-29.
-
Szidat L (1965) Estudios sobre la fauna de parasitos de peces antarticos. I. Los parasitos de Notothenia neglecta Nybelin. Servic Hidrogr Naval Secr Mar de la Rep Argent 910: 1-84.
-
Hargis WJJr, Dillon WA (1968) Helminth parasites of Antarctic fishes: the superfamily Capsaloidea Price, 1931. Proc Biol Soc Wash 81: 403-412.
-
Yamaguti S (1968) Monogenetic trematodes of Hawaiian fishes. University of Hawaii Press, Honolulu, pp: 287.
-
Gibson DI (1976a) Monogenea and Digenea from fishes. Discov Rep 36: 179-266.
-
Szidat L (1969) Estudios sobre la faune de parasitos de peces antarticos III Pseudobenedenoides branchicola gen nov, sp nov (Trematoda, Monogenea, Capsalidae). Un nuevo parasito branquial del Pez Antartico Trematomus bernacchii. Neotropica 15: 25-32.
-
Menzies R J (1946) Benedenia noblei, a new Monogenetic trematode of the Pacific coast rock fish. The Journal of Parasitology 32(4): 428-430.
-
Gibson DI (1976b) Menziesia (Monogenea: Capsalidae), a new name for Parabenedenia Gibson, 1976, nec Johnston, 1929. J Helminthol 50(2): 98.
-
Timofeeva TA, Gaevskaja AV, Kovaleva AA (1987) Capsalids (Monogenea) of the notothenioid fishes from the Atlantic region of Antarctica and Subantarctica. Tr Zool Inst 161: 78-93.
-
Rubtsova NY, Chaudhary A, Salganskiy OO, Kuzmina TA (2023) Description of Pseudobenedenia coriicepsi sp. n. (Monogenea: Capsalidae) from the Antarctic Black Rockcod, Notothenia coriiceps Richardson in Coastal Waters of West Antarctica Using Novel SEM Images, Energy Dispersive X-Ray Analysis and Molecular Analysis. Int J Zoo Animal Biol 6(5): 000512.
-
Dettai A, Berkani M, Lautredou AC, Couloux AG, Lecointre G, et al. (2012) Tracking the elusive monophyly of nototheniid fishes (Teleostei) with multiple mitochondrial and nuclear markers Marine Genomics 8(2012): 49-58.
-
(2024) Elchmeyer Catalog of fishes.
-
Froese R, Pauly D (2023) FishBase. World Wide Web electronic publication.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind