An update on the Genetics, Cellular and Evolutionary Implications of X Chromosome Demasculinization
Sex chromosomes and autosomes evolve in different trajectories depending on gene expression during gametogenesis. The X chromosome gene content across animal taxa shows opposite tendencies in different species implicating the underlying genetic basis of adaptive changes. The constraints imposed on the X chromosome by meiotic sex chromosome inactivation (MSCI) and related spermatogenesis-specific regulatory processes underscore the dynamic interplay between developmental regulation and evolutionary adaptation. The ying–yang functions of MSCI results in distinctive genomic architecture in which the X chromosome is enriched for early spermatogenic genes while the late spermatogenic functions are relocated or duplicated to autosomes. In the present review using comparative evidence across taxa as a template we explore principles of sex-linked repression and germline expression constrains. Major observations include differential allele expression level of X-linked genes in males compensating for the degeneration of the Y-linked homologs. Faster evolve of ampliconic gene families on Y chromosome. Therefore, even in predominantly diploid organisms, haploid selection may shape sex chromosome evolution and reproductive strategies. This form of selection acting on genes expressed in gametes, where the absence of allele masking enables more effective purging of deleterious mutations and more efficient fixation of beneficial ones. The far ranging implications of these mechanisms include sperm biology, fish conservation genetics and susceptibility to Human Diseases. In summary, chromosome-wide regulatory mechanism act as an evolutionary filter, redistributing essential reproductive functions and reinforcing male-specific transcriptional programs.
Introduction
The X chromosome gene content is not stable across animal taxa and shows opposite tendencies in different species. Patterns of sex chromosome and autosome evolution can be used to elucidate the underlying genetic basis of adaptive change. Evolutionary theory predicts that X-linked genes will adapt more rapidly than autosomes if adaptation is limited by the availability of beneficial mutations and if such mutations are recessive. An obvious question is what could be driving unusual patterns of gene content evolution on the X? The fact that one sex possesses two copy of the X/Z chromosome while the other has one copy can result in different selective pressures. This pattern is particularly apparent in the distribution of genes with different fitness consequences in the two sexes. The X chromosome evolves differently from autosomes as a result of its sex-biased transmission, homozygosity in males, and X chromosome inactivation during spermatogenesis. In the genus Drosophila, ancestral and recently formed neo-X chromosomes show an underrepresentation of male-biased genes on the X chromosome. Comparative genomics studies have shown that depletion of male-biased genes is caused by higher rates of gene loss and lower rates of establishment of novel male biased genes. Further, existing genes with male- biased expression escape from the X to autosomes through retroposition. How exactly different genetic and evolutionary forces influence patterns of chromosome evolution, sex- biased expression and gene content and the underlying cytogenetic and cellular mechanisms is the objective of this review. The background areas of Sex chromosome differentiation and developmental roles dimorphism and effects form the introduction. The dosage differential gene and protein expression lay foundations for later sections on Evolutionary consequences. The unique trajectory of autosomes and sex chromosomes during mitosis and meiosis along with the cellular mechanism of inactivation provide details of the mechanism and nuances and impact on genetics and speciation.
Developmental Origin of Sex Chromosomes and Autosomes
Sex chromosomes are recognized by their specific features, including reduced recombination, degeneration, heteromorphic size, dosage compensation, specialization (often sex-specific) gene content, and expression [1]. The X and Y chromosomes are proposed to have evolved from an ordinary pair of autosomes that stopped recombining after acquiring a sex-determining role [2]. Sex determination is controlled by sex chromosomes in many separate-sexed organisms. In these species, a major sex-determining region determines sexual development, which triggers a cascade of sex-specific genes that regulate development of the male or females [3, 4]. While the Y chromosome degenerates due to a lack of recombination, however the X chromosome retains the majority of its ancestral genes during its evolutionary trajectory [5, 6]. The accumulation of sexually antagonistic genes associated with the sex-determining genes favours the evolution of suppression of recombination between the nascent sex chromosomes. These initially homologous chromosomes continue to differentiate in the absence of recombination [7]. Sex chromosomes (XX in females, XY in males) determine sex in humans. Although the X and Y chromosomes harbour dramatically different numbers and sets of genes (about 1,000 genes on the X and only a few dozen genes on the Y), they have originated from ordinary autosomes during the early evolution of mammals [8]. The morphological differentiation of sex chromosomes is the result of recombination restriction followed by gene loss on the Y. The Sry gene, a single master sex-determining locus on the male-limited Y chromosome, determines gender. Expression of Sry early in embryonic development initiates testis differentiation by activating male-specific developmental networks, while in its absence; ovaries develop. By the sixth week of human pregnancy, the ovary and testis show the first obvious indications of sexual differentiation [9], and sex hormones cause additional sexual differentiation in non-gonadal tissues and organs. Figure 1 summarises various events during Mitosis and Meiosis highlighting sex-based gene expression.
Sex chromosomes have emerged many times in independent plant and animal lineages. The placental mammals share a sex chromosome pair that originated ∼180 Ma [11]. In the vast majority of mammal species, the Y chromosome is sex-determining: presence of the Y-encoded protein SRY is sufficient to initiate the male developmental program. Since their divergence from the ancestral X chromosome, mammalian Y chromosomes have lost nearly all of their ancestral gene content [12]. The evolutionary diversity of Y chromosomes in mammals arises from the set of Y-acquired genes, which make up a small fraction of some Y chromosomes and a larger fraction in others—from 5% in rhesus monkey to 45% in human [13]. These genes are often present in many copies and are highly specialized for function in the male germline [14]. The Y chromosome of the house mouse (Mus musculus) stands out among mammalian Y chromosomes in terms of size and unusual gene repertoire such as size, gene-rich and euchromatic features. Copy number of genes on the repetitive long arm of both sex chromosomes is highly variable, but sequence diversity in non-repetitive regions is decreased relative to the autosomes.
![Figure 1: Trajectories of various variables during Mitosis and Meiosis highlighting sex-based gene expression [10].](/fulltextimages/14016/fig_1.png)
Not all sex chromosomes become substantially differentiated, even if certain organisms (like dragonflies and crickets) have lost the Y chromosome completely [15]. Sex-reversed female frogs (i.e., those with an XY genotype) introduce new genetic variance on the Y because their sex chromosomes recombine as much as those of XX-females. However, this only works for species with relatively undifferentiated sex chromosomes-strongly differentiated sex chromosomes cannot recombine successfully [16]. In Fish with chromosomes either (XY males) or (ZW females) systems the sex chromosome turnovers, is caused by fusions between autosomes and pre-existing sex chromosomes, the development of a novel sex-determining gene on an autosome, or due to the transposition of a sex-determining locus to an autosome [17]. Environment (temperature or social interactions) has a profound role in determination of sex [18]. In Stickleback fish the chromosomal mechanism for sex determination is based on a nascent 10 million years old Y chromosome. Further, they lack sexually dimorphic chromosomes, although closely related species do have distinguishable sex chromosomes in either males or females [19]. In flies and mammals, for instance, males are the heterogametic sex, with X and Y sex chromosomes; where as in birds and butterflies, females are heterogametic, possessing Z and W sex chromosomes, the acquisition of a major sex- determining locus by an ancestral pair of autosomes can trigger their gradual evolutionary transition into differentiated sex chromosomes [20]. Since Y chromosomes are sex-limited and non-recombining, the X and Y can evolve differences in size, morphology, gene content and modes of regulation [21]. In the genus Drosophila fusions between sex chromosomes and autosomes, resulting in neo-X and neo-Y chromosomes, and also between autosomes is observed [22]. The genes located on the dot chromosome of Drosophila are X-linked in outgroup species, whereas Drosophila X-linked genes are autosomal. Table 1 summarizes the Chromosome systems and determination mechanisms in various organisms.
| Organism | Human and other mammals | Drosophila | Insects and other organisms | Birds, reptiles, some fishes and amphibians |
|---|---|---|---|---|
| Chromosomal System | XX-XY | XX-XY (XA system) | XX-XO | ZZ-ZW |
| Chromosomal method of determination | Presence of Y chromosome triggers maleness during fetal development | Ratio of X:autosomal chromosomes determines sex | Number of X chromosomes determines sex; there is no Y chromosome. | Two Z chromosomes required for maleness |
| Mechanism | SRY gene on the Y chromosome sets off a series of events leading to development of testes. (Other genes both autosomal and sex-linked are also involved) | Expression of sex- determining genes on the X-chromosome is affected by timing (and level) of expression of autosomal genes | May vary among species: In C. elegans is related to the amount of an X-encoded protein which can inhibit maleness | DMRT1 gene on the Z chromosome is haploinsufficient; one copy of the gene can’t produce enough protein to trigger male development |
Table 1: Summary of Chromosome systems and determination mechanisms.
Evolutionary Consequences on Sex Chromosomes
The specific sets of chromosomes that individuals carry determine sex in many animals and some plants. For example, males in flies and mammals are heterogametic, with X and Y chromosomes, while females in birds and butterflies they are heterogametic with Z and W chromosomes [23]. The effects of evolution on chromosomes include differential chromosome evolution, gene expression, varied rates of protein evolution, and gene family evolution. We summarize these genetic events in the following paragraph highlighting unique observations. According to the conventional hypothesis, the gradual evolutionary transition into differentiated sex chromosomes is initiated when an ancestral pair of autosomes acquires a significant sex-determining gene through either de novo gene evolution or the relocation of an established sex determination gene [24, 25]. Because the Y chromosomes are sex-limited and do not recombine, the X and Y chromosomes evolve in different sizes, morphologies, gene content and regulatory modalities. In Drosophila, the number of X chromosomes serves as the initial signal for sex determination [26], with males being hemizygous for 20% of the genome. Once established, differentiated sex chromosomes appear to represent an evolutionary endpoint—autosomes can evolve into sex chromosomes, but the opposite is not true. The genetic differentiation and sex-biased inheritance of sex chromosomes make them vulnerable to genetic transmission conflicts, such as meiotic drive. Further, a defective Y chromosome causes a dosage discrepancy for X-linked genes promoting the evolution of dosage compensation mechanisms [27]. In Drosophila melanogaster multi-locus deletions (deficiencies) are tolerated when the size of the deletion is small, but when 0.1% of the genome is hemizygous this reduces viability [28]. However, the X chromosome in males is an exception to the rule. This has happened numerous times throughout the evolution of sex chromosomes [29].
In Drosophila melanogaster X chromosome dosage avoids male lethality by equalizing the expression level of genes on the X chromosome in males and females [30]. The Male-Specific Lethal (MSL) gene is the major determinant of dosage correction in comparison to autosomes [31]. X chromosome genes that do not show MSL connection show dosage compensation while certain X chromosome genes are dosage compensated extremely early, prior to the activation of the MSL system. Finally, removing the MSL complex components does not reduce X chromosome gene expression in relation to autosomes, implying that alternative MSL- independent processes may also contribute to dosage compensation in fly somatic cells [32]. In Drosophila S2 cells, the observed autosomal dosage compensation level is equal to approximately 1.4-fold “missing” compensation following MSL complex knockdown, indicating that MSL and general dosage compensation mechanisms could have an additive effect. Other contributing factors include unique chromatin signatures of X chromosome significantly differing from autosomes even in females [33]. And the gene traffic of the genes on X may force the dosage-sensitivity [34]. Finally, the gradual development of dosage compensation throughout sex chromosome evolution may make the X more or less susceptible to gene dosage variations, depending on which forces are dominant. In Caenorhabditis elegans, male-biased genes are underrepresented on the X chromosome, accounting for only 3% of the expected number of spermatogenic genes [35]. C. elegans exclusively underrepresents genes expressed in the male germline on the X chromosome, but not in somatic cells consistent with X inactivation playing a significant role in demasculinization. Hermaphrodite-specific genes expressed in somatic cells are enriched on the X chromosome, implying that sexually antagonistic selection may influence the genomic distribution of these genes [36]. In Anopheles gambiae, male-biased genes are uniformly dispersed throughout the genome [37]. Whereas, in mice, there is a link between the stages of spermatogenesis (and accompanying X inactivation) and the presence of male-specific genes on the X chromosome [38]. Mouse genes expressed during mitosis (before X inactivation) are overrepresented on the X, whereas genes expressed later in spermatogenesis (when the X chromosome becomes transcriptionally inactivated) are depleted, and genes expressed during meiosis are completely absent [39]. These findings lead to the conclusion that harbouring male- specific genes on the X has a selective advantage in mice—as expected for recessive male-beneficial mutations. However, inactivating the X chromosome later in spermatogenesis results in a lack of late-acting spermatogenesis genes on the X. In chickens with a ZW sex-determination system, ovary- specific genes are markedly underrepresented on the Z chromosome, but no enrichment of testis-expressed genes is observed [40]. Sex-chromosome inactivation has not been observed in heterogametic female individuals, implying that Z-inactivation is not a likely factor affecting gene distribution in chickens [41]. These examples demonstrate that X chromosome gene content patterns are not universal, and further research is needed to shed light on these variances across taxa. Furthermore, for genes expressed in the brain, the Z chromosome shows an enrichment of male-biased genes and an underrepresentation of female-biased genes [42]. This closely resembles patterns of gene content evolution in organisms with XY sex determination systems. Mammalian studies have revealed that genes that are preferentially expressed in male somatic tissues are often enriched on the X chromosome [43]. The X chromosome is enriched in genes associated to muscle function, sex, and reproduction, as well as brain-expressed genes [44]. In summary, this implicates that sexual antagonism influences the evolution of gene content on the X chromosomes in animals.
Meiotic sex chromosome inactivation (MSCI) or precocious transcriptional silencing of the sex chromosomes during male meiosis has developed separately in multiple lineages, possibly to suppress sex-linked meiotic drive elements in the male germline [45]. In Drosophila, genus approximately 12 fusions between sex chromosomes and autosomes produce neo-X and neo-Y chromosomes and there are 23 fusions between autosomes [46]. Vicoso and Bachtrog 2013 demonstrate that the ancient Dipteran X chromosome reverted to an autosome in the pathway that led to Drosophila. In Drosophila, Muller element A forms the X chromosome, and sex is determined by the relative dosage of the X-linked sex determination gene Sex fatal [47]. While the gene composition of the Muller elements is mostly consistent throughout Diptera, however the X chromosome is ‘demasculinized,’ with few male-biased genes. Further, the dot-like X appears to be demasculinized in basic dipteran species [48]. While supporting the case for the dot’s X-to- autosome transition, there is another explanation for the decreased average expression of X-linked genes in the testis. Several recent investigations have discovered that the gene content of the D. melanogaster X chromosome is not demasculinized [49]. In summary, the different sizes, morphologies, gene content and regulatory modalities of sex and autosomes are determined by specific genes, gene transfer and chromatin states on X or Z chromosomes. However, this is not a universal dictum. Dosage discrepancy for X-linked genes promotes evolution of dosage compensation mechanisms. Also, male-specific genes on the X confer selective advantages for recessive male-beneficial mutations. These factors contribute towards antagonism influencing the evolution of gene content on the X chromosomes.
Differential Gene and Protein Expression
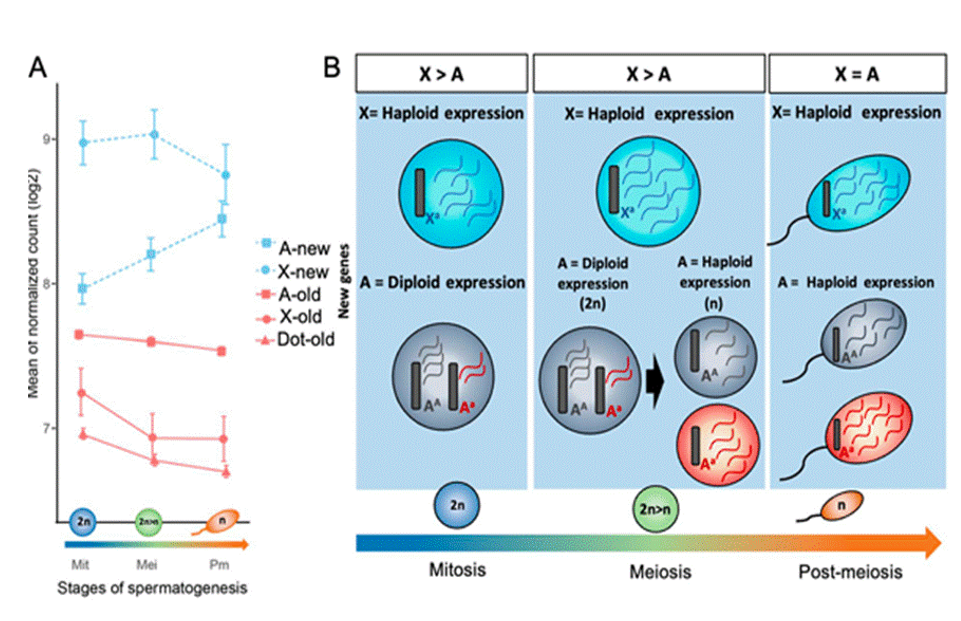
Ohno in 1970 proposed that, during the creation of the mammalian sex chromosomes the per-allele expression levels of X-linked genes in males should be quadrupled to compensate for the degeneration of the Y-linked homologs. This proposed 2-fold overexpression of X-linked genes, which is likely also present in females, explaining why one of the two X chromosomes in females must be inactivated. However, this proposal has few exceptions, a Xa:AA ratio of 0.5 was discovered by RNA sequencing (RNA-seq) [50], also in human proteomic data analysis [51]. Another study of human X-linked genes using RNA-seq expression comparison and their chicken autosomal orthologs which represent proto-X genes, showed the absence of the expected 2-fold increase in mammalian X chromosome evolution [52]. Intriguingly, X-linked genes that encode components of large protein complexes (5%) showed a 2-fold increase in human RNA-seq data [53]. It’s worth mentioning that Ohno’s hypothesis is about diploid cells. In haploid cells, the number of X chromosomes relative to the number of autosome sets is 0, 1, and 1 for Y-carrying sperm, X-carrying sperm, and eggs, respectively [54]. Apparently, X-chromosome dosage adjustment is impossible for Y-carrying sperm and unnecessary for eggs and X-carrying sperm [55]. X is repressed during male meiosis but restored in spermatids [56, 57]. Expression data suggested an Xa:A expression ratio of 1 in X-carrying spermatids and 0.9 in secondary oocytes (i.e., post-meiosis I) [58]. Further, the evolutionary fates of mutant alleles may differ between sex chromosomes and autosomes [59]. For example, in an XY system, a male- benefit mutant allele of a Y-linked gene can spread even if it is potentially lethal to females, whereas invasion of a population by a beneficial autosomal mutation requires a larger fitness benefit to males than any cost to females. The segregation of sexually antagonistic alleles at genetically related loci promotes the recruitment of chromosomal inversions, facilitating genetic links between maleness and male-beneficial [60]. Most modeling work on this aspect of sex chromosome evolution has focused on a fully sex-linked region with no recombination in the heterogametic sex, with a “genetically degenerate” Y chromosome that has lost nearly all genes present on the X, so that males are generally hemizygous for X-linked loci [61]. In the D. melanogaster lineage, X-linked genes account for an equal fraction of adaptive nonsynonymous changes as autosomal genes [62]. Unidentified random or systematic discrepancies between X-linked and autosomal genes may hide a faster-X evolution pattern. A large number of studies have investigated these hypotheses by comparing the evolutionary rates of X-linked and autosomal genes [63, 64, 65]. Thus, X linkage does not seem to increase the pace of protein adaptation in species like Drosophila.
Gene Family Evolution
Y chromosomes often consist of very few functional genes however many of these genes have undergone massive gene amplification and persist as members of multi-copy gene families. For instance, the human Y chromosome harbors nine multi-copy ampliconic gene families constituting the majority of protein-coding genes present on the Y [66]. Why these ampliconic gene families have evolved on heteromorphic sex chromosomes is a question and their phenotypic consequences form intriguing research problems in evolutionary biology. The copy number of these gene families can vary substantially, not only across closely related species but also individuals of the same species. The primate Y chromosome is distinguished by a lack of inter- chromosomal recombination along most of its length, with extensive gene loss, and a prevalence of repetitive elements. Ghenu, et al. [67] report evidences for several novel examples of gene conversion in papionin monkeys indicating that the ampliconic gene families evolve faster than autosomal gene families than single-copy genes. Also, the Y-linked singleton and autosomal gene families evolved faster in humans and chimps than they do in the other Old World Monkey lineages. Evidence for gene amplification on the duck W chromosome is weak, with only two of the 26 W-linked genes are present in multiple copies [68]. In A. pisum pea aphid genome analysis reveals an elevated incidence of both young and old gene duplications on the X chromosome in comparison to autosomes [69]. Finally, in D. melanogaster genes in the takeout gene family show male-biased expression. The phylogenetic analysis revealed conservation across insects with evidence of positive selection in some lineages and sex specific roles in male mating [70]. Figure 2 summarizes Chromosomal, Cellular and Genetic regulatory events leading to sexual differential in metazoans.
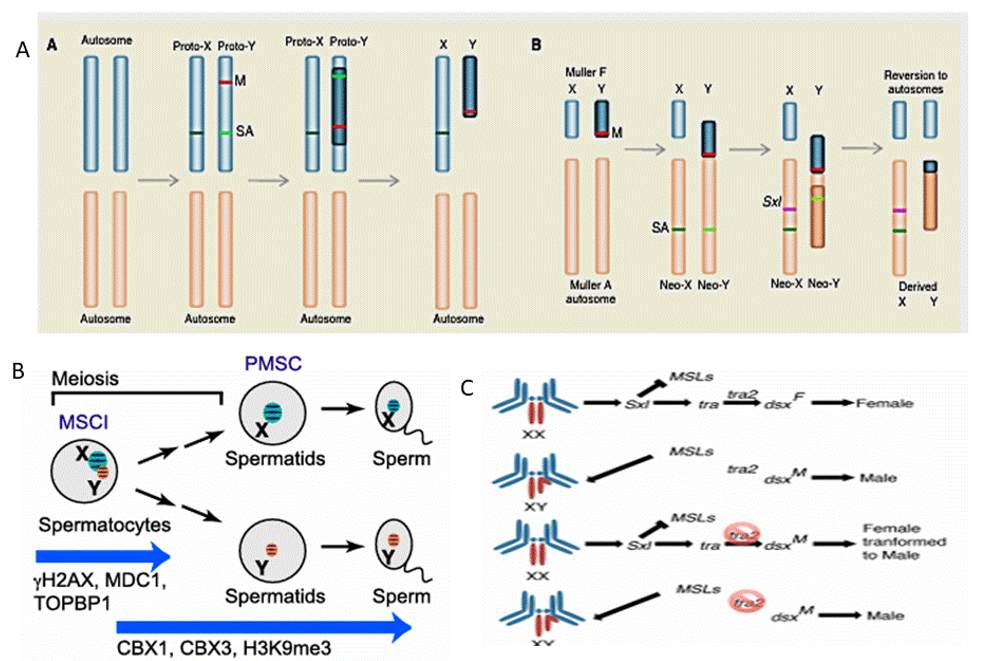
In summary, allele expression level of X-linked genes in males is compensated for the degeneration of the Y-linked homologs. Chromosomal inversions facilitate genetic links between maleness and male-beneficial alleles. The evolutionary fates of mutant alleles may differ between sex chromosomes and autosomes. Y chromosomes have undergone massive gene amplification harbouring multi- copy gene families with implication for intra-inter species evolution. Finally, the phenomenon has implications for fitness benefits of population.
Gene Movement and the Evolution of Male- Biased Gene Expression
Relocation of male-biased genes in testis tissue and their relative depletion from sex chromosomes is common among across many animal taxa [71]. This process is usually mediated by retrotransposition or DNA-based duplication, and it has consequences for gene regulation and sex biased expression [72]. The X chromosome, despite harboring many ancestral genes, is subject to specific evolutionary restrictions due to male hemizygosity, dosage compensation mechanisms, and meiotic silencing [73]. The selection factors that promote gene mobility are related to regulatory events meiotic sex chromosome inactivation (MSCI) during spermatogenesis, which silences or downregulates sex- linked [74]. Gene movement is significant because it enables critical male-biased genes to escape regulatory settings on the sex chromosomes. In species where MSCI occurs, the X is transcriptionally repressed in pachytene spermatocytes, leaving X-linked genes nonfunctional at critical stages [75]. To compensate, relocated or duplicated copies on autosomes can maintain expression, ensuring functional continuity. Furthermore, movement off the X may reduce conflicts caused by sexually antagonistic selection, where alleles beneficial to males may be detrimental to females [76]. Male-biased genes, particularly those expressed during spermatogenesis, shows rapid rate of evolution and unusual chromosomal distributions compared to unbiased or female-biased genes [77]. The migration of genes between chromosomes, particularly from sex chromosomes to autosomes, and the resulting changes in expression patterns is a notable characteristic of these dynamics. During the mitotic phase, early spermatogenic genes are preferentially expressed from the X chromosome in mammals. The hemizygous exposure of these X-linked genes in males enables more efficient selection on recessive, male-beneficial alleles—a phenomenon described by the Faster-X effect [78]. In Drosophila, young X-linked genes can establish expression in early germline stages before evolutionary pressures and regulatory constraints. Thus, the mitotic stage highlights the adaptive potential of X-linked gene expression in shaping reproductive fitness. Additionally, during meiosis, meiotic sex chromosome inactivation (MSCI) causes genome-wide repression of X-linked genes at the pachytene stage, defining a significant turning point in male germline transcriptional regulation [79]. This mechanism causes redistribution of spermatogenic activities. Genes required for mid- and late spermatogenesis are markedly underrepresented on the X, but become enriched on autosomes by mechanisms such as retrotransposition and duplication. In C. elegans, the effect of MSCI is particularly pronounced, as spermatogenesis-specific genes are almost completely absent from the X chromosome [80]. Mammalian data reinforce this stage-specific bias. The selective relocation of genes ensures continuity of expression during meiotic silencing, safeguarding the reproductive success. Following meiosis, haploid spermatids exhibit a transcriptional pathway dominated by autosomal expression. At this stage, haploid selection—the direct exposure of recessive alleles in gametes - increases the efficacy of both purging deleterious mutations and fixing advantageous alleles. As a result, autosomal genes not only compensate for the absence of X expression, but also act as hotspots for rapid evolutionary change in sperm-related features. Comparative studies show that the bulk of late spermatogenesis genes in mammals and insects are autosomal, indicating their stability under haploid selection pressures [81]. In contrast, the X chromosome is essentially silent (repressed), emphasizing its limited function in post-meiotic transcription. Together, these stage-specific dynamics highlight a recurring concept in genome evolution, and the temporal partitioning of X-linked and autosomal expression during spermatogenesis. The X chromosome provides adaptation during mitosis, but its suppression during meiosis mandates the transfer of male-biased genes to autosomes. Post-meiotically, haploid selection reinforces the autosomal bias of male-specific expression, influencing both genome architecture and reproductive choices. Thus, the cycle of enrichment, silencing and relocation are an evolutionary mechanism that connects gene mobility, male fertility and species divergence.
The movement of male-biased genes from sex chromosomes to autosomes occurs through several mechanisms that leave distinct evolutionary signatures. Retrotransposition is the most prevalent pathway, particularly in metazoans, where intron less retrocopies of X-linked genes is reinserted into autosomes and frequently acquires testis- specific expression [82]. A well-studied mammalian case is the PGK2 gene, which originated as a retroposed autosomal copy of the ubiquitously expressed PGK1 gene, providing a meiosis-specific glycolytic function in spermatogenesis [83]. Figure 3 depicts gene expression dynamics of newly expressed and older genes on x-chromosome, autosomes and dot chromosomes of Drosophila. Gene duplication and sub functionalization also contribute, duplicated autosomal paralogs often evolve spermatogenic functions, while their X-linked progenitors retain broader somatic roles, thus enabling escape from meiotic sex chromosome inactivation (MSCI) without loss of ancestral function [84]. However, chromosomal translocations and rearrangements move larger blocks of genes between chromosomes, reshaping composition and accelerating divergence between lineages less frequently [85]. Collectively, these mechanisms ensure that essential spermatogenic genes can circumvent MSCI, maintaining functional expression during meiosis and post- meiosis.
Genetic Constraints Due to MSCI and other Spermatogenesis-Specific Regulatory Effects
The regulation of sex chromosomes during spermatogenesis is a critical axis in both developmental biology and evolution. The X chromosome, in particular, confronts specific restrictions to balance dosage, supporting male fertility and navigate transcriptional silencing during meiosis. A defining feature of this regulatory landscape is meiotic sex chromosome inactivation (MSCI), a process by which the X and Y chromosomes are transcriptionally silenced at the pachytene stage of meiosis [86, 87]. Unlike somatic X chromosome inactivation (XCI), which balances gene dosage in females, MSCI appears to reflect an ancient genome defense system, now integrated into mammalian spermatogenesis [88]. The constraints imposed by MSCI extend beyond transcriptional repression. They have shaped the evolutionary fate of X-linked genes, driving gene traffic from the X to autosomes, influencing the timing of spermatogenic gene expression and also imposing selective pressures on genomic architecture.
MSCI is distinct from somatic X chromosome inactivation (XCI) in terms of initiators and molecular logic. In XCI Xist- (X-encoded long noncoding RNA) initiates somatic XCI by nucleating polycomb repressive complexes (e.g., PRC2) and histone deacetylases, which deposit H3K27me3 and stabilize long-term silence [89]. The MSCI relies on DNA repair kinases and sensors. The signaling cascade involves the recruitment of (ATR) by (BRCA1) to unsynapsed chromatids, and phosphorylation of H2AX to form γH2AX by ATR, with subsequent DNA-damage triggering of repressive modifications [90, 91]. From an evolutionary perspective, MSCI has profound consequences for X chromosome gene content. Functional genes involved in late spermatogenesis are selected on the X since they would be silenced at the critical window. The result is recurrent gene traffic from X to autosomes via retrotransposition and duplication creating an autosomal “backups” expressed when MSCI would otherwise repress their X-linked progenitors [92, 93]. Comparative
surveys reflect this pattern across taxa. C. elegans and many mammals show depletion of late spermatogenesis genes on the X, whereas genes expressed before MSCI (early spermatogenic genes) may be retained or even enriched on the X. Thus, MSCI acts as an evolutionary filter that redistributes male-fertility functions in the genome.
Sex chromosomes evolved from a pair of autosomes after acquiring a sex-determining locus, which triggered recombination suppression between proto-X and proto-Y chromosomes [94]. Over time, this led to degeneration of the Y and enrichment of functional genes on the X, along with the evolution of dosage compensation mechanisms. In Drosophila, dosage compensation occurs through hypertranscription of the male X mediated by the male-specific lethal (MSL) complex, whereas in mammals, females balance dosage by inactivating one X via the X-inactivation center (XIC) and the long noncoding RNA Xist [95, 96]. In the male germline, however, the single X does not undergo upregulation during meiosis but instead becomes transcriptionally silenced, necessitating alternative compensatory mechanisms such as the relocation of spermatogenic genes to autosomes [97]. Comparative studies reveal conserved patterns of X constraint across species. For instance, in C. elegans, spermatogenesis- specific genes are drastically underrepresented on the X. In mammals, early spermatogenic genes are enriched on the X, while late-stage genes are absent, reflecting the timing of MSCI. In birds with ZW system, female ovary-specific genes are underrepresented on the Z chromosome, highlighting a general principle of sex-linked repression shaping gene content [98].
Beyond its restrictive role in silencing late spermatogenic genes, MSCI also contributes to male germline masculinization by shaping a transcriptional program that reinforces male- specific reproductive functions. The currently prevailing hypothesis is the selective retention of recessive, male- beneficial alleles on the hemizygous X chromosome, which facilitates their exposure to selection during spermatogenesis [99, 100]. Additionally, MSCI has been implicated in establishing epigenetic imprinting patterns, providing a potential template for paternal X silencing observed in early female embryos [101]. Another hypothesis links MSCI to the suppression of meiotic drive systems, thus balancing segregation of sex chromosomes [102]. Taken together, these evidences suggest that MSCI is not merely a constraint but also a driver of male-biased transcriptional architecture, contributing to both the stability and evolutionary trajectory of male reproduction.
Evolutionary Implications
Theories suggest that sex chromosomes and their roles in determining sex and sexual traits are central to processes such as speciation, sexual selection, and the evolution of dimorphic characteristics. Further, X chromosome inactivation, mosaicism, skewing, and dimorphism in X chromosome genes play crucial role in species evolution. This could be relaxed as in certain crustacean Gammarus duebeni or labile as in fishes and reptiles, provide advantage in environments with fluctuating ecological pressures [103, 104]. Heteromorphic sex chromosomes, in particular, contribute strongly to the formation of reproductive barriers, which are keys to speciation. This is reflected in the “two rules of speciation.” as per Haldane’s rule which posits that the hybrid sterility occurring in only one sex is usually the heterogametic sex. Second, the large X/Z effect explains that hybrid dysfunction caused by genetic incompatibilities tends to map to the X or Z chromosomes in heterogametic systems [105]. Evidence from diverse species such as Drosophila, butterflies, mammals, and birds implicates that Haldane’s rule is a widespread evolutionary pattern and plays critical roles in establishing reproductive barriers necessary for speciation [106].
Sex chromosome degeneration and the consequent mechanisms of dosage compensation have potential to generate incompatibilities in hybrids when disrupted and thus be an instrument of reproductive isolation. Additionally, recessive mutations on the X or Z chromosome are more exposed to selection than in autosomes, where they can be overridden/neutralized in a heterozygous genotype [107]. This phenomenon is also linked to stronger genetic drift, which can accelerate the fixation of mildly harmful mutations on sex chromosomes. Such changes may quickly drive divergence between the sex chromosomes of newly forming species, thereby promoting reproductive isolation [108]. In two hybridizing nightingale species, stronger genetic drift on the Z chromosome suggests a case of faster-Z evolution, which likely promotes the buildup of hybrid incompatibilities and supports the large Z effect [109]. The lack of faster-Z evolution in paleognaths with uniform sex chromosomes explains the role of purifying selection. Meiotic drive—the unequal segregation of genetic elements caused by intragenomic conflict tends to impact sex chromosomes and in short timescales, alter genomes in ways that generate reproductive barrier [110]. Under particular conditions, it may also help explain the established rules of speciation. Additionally, divergences in sex differentiation pathways can produce incompatibilities that reinforce reproductive isolation. Such barriers may arise from mutations in master switch genes, changes in gene copy number, or large-scale alterations in sex chromosomes, including loss of the sex-determining gene, translocations, or even complete chromosome loss [111]. Overall, the evidence suggests that reproductive isolation—particularly postzygotic isolation—is more pronounced in species with sex chromosomes, especially those with heteromorphic pairs.
Earlier theories proposed a link between sex chromosomes and sexual dimorphism, but this relationship is much more complex than initially assumed, as it also involves significant roles for autosomal genes, hormones, and epigenetic mechanisms [112, 113]. Sexual dimorphism can manifest in traits such as coloration, body size, shape, and behavior, many of which are tied to reproduction and mate choice, making them subject to sexual selection [114]. The evolution of these dimorphic traits is likely a circular process—sex-specific selective pressures influence gene expression that produces dimorphic phenotypes, which shape species’ behaviors, and these behaviors, in turn, modify the selective pressures driving gene evolution [115]. In summary, sex chromosomes associated mechanisms— appear to play important roles in reproductive isolation and speciation for several reasons however few caveats need to be addressed.
Discussion
Sex chromosomes and autosomes evolve in different trajectories depending on gene expression during gametogenesis. When genes are active in gametes (such as sperm), selection is stronger because detrimental mutations are easier to eliminate and beneficial mutations spread more quickly. This is especially true for X-linked genes, because in males, the ‘X’ is hemizygous; hence selection acts directly - a concept known as the Faster-X theory. Autosomes on the other hand, typically expose their hidden alterations during the haploid stage (after meiosis), resulting in an altered adaptive expression. This allows new autosomal genes to reach the same stage of X-linked genes, though they seldom outperform them. Over time, older testis-expressed genes move off the X chromosome due to constraints like MSCI (X silencing during meiosis), while new X-linked genes may initially avoid these limits before relocating to autosomes. Together, these patterns underscore the dynamic relationship between sex chromosome regulation, gene movement, and the evolution of male-biased gene expression. This sustained expression across mitosis, meiosis, and post-meiosis of X-linked new genes aligns with the concept of haploid selection and supports the Faster-X hypothesis, which suggests that the hemizygous state of the X chromosome in males accelerates the evolution of X-linked genes compared to autosomes. In contrast, autosomal genes only expose recessive adaptive mutations to selection during the haploid stages. As a result, the expression boost driven by haploid selection is primarily seen at the end of meiosis and into post-meiosis, a phenomenon unique to autosomal genes. MSCI and other spermatogenesis-specific regulatory constraints continue to influence X chromosome gene content, driving the relocation of male-biased genes to autosomes, while newly emerged genes on the X may initially evade these pressures. This allows newly evolved autosomal genes, driven by positive selection, to reach expression levels comparable to new X-linked genes. Selection acts on genes expressed in gametes, where the absence of allele masking enables more effective purging of deleterious mutations and more efficient fixation of beneficial ones. Therefore, even in predominantly diploid organisms, haploid selection may shape sex chromosome evolution and reproductive strategies. Plausible questions emerging from the review are: a. How does haploid selection vary between species with strong and weak sperm competition; b. Meiotic and post-meiotic differences between X and autosomes; c. X-chromosomal evolution across taxa.
Afore discussed genetic factors could drive rapid divergence in sperm forms, function, and rates of production. Sperm are one of the most morphologically diverse cell types in animals exhibiting varying degrees of evolutionary loss, convergence, and recurrences attributed to postcopulatory sexual selection. This has implications for fertility and reproduction [116]. The dynamics discussed not only influences gene migration between chromosomes, but it also has implications for broader questions like fertility, adaptation and genome organization. Failure or defects in MSCI are directly linked to male infertility in humans and other mammals. MSCI defects can lead to a meiotic arrest at the mid-pachytene stage, a critical point in sperm development. MSCI mis-expression contributes to the loss of germ cells [117].
Species subject to environmental of sex determination face higher risks to fluctuating ecological changes. These dynamics are important in understanding evolutionary adaptations and guiding conservation efforts, in light of ongoing global and regional climate change [118]. Epigenetic mechanisms such as X chromosome inactivation, mosaicism, skewing, and dimorphism in X chromosome genes and Y chromosome regulatory genes create a sex-based variance and susceptibility in the immune response between males and females [119]. Sex chromosome complement and sex-differentiated epigenetic mechanisms to control gene expression are decisive players in the sexual dimorphism of Pulmonary Arterial Hypertension (PAH) [120]. In summary, the review highlights how haploid selection influences sex chromosome regulation, gene movement and male-biased gene expression [121, 122, 123, 124, 125, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149].
Acknowledgements
We thank the VC and Principals of GSC, and the Oxford Engineering college Bangalore for encouragement. Dedications: To the lotus feet of Bhaghavan and Gurus of Sringeri peetam Funding: Pooled by the faculty Ethics/Animal studies declaration: Since a review not required
References
-
Ubeda F, Patten MM, Wild G (2015) On the origin of sex chromosomes from meiotic drive. Proc Biol Sci 282(1798): 20141932.
-
Muyle A, Shearn R, Marais GAB (2017) The Evolution of Sex Chromosomes and Dosage Compensation in Plants. Genome Biology and Evolution 9(3): 627-645.
-
Bachtrog D, Kirkpatrick M, Mank JE, McDaniel SF, Pires JC, et al. (2011) Are all sex chromosomes created equal?. Trends Genet 27(9): 350-357.
-
Feingold KR, Ahmed SF, Anawalt B, Blackman MR, Boyce A, et al. (2000) Endotext [Internet]. South Dartmouth (MA): MDText.com, Inc.
-
Bachtrog D (2006) A dynamic view of sex chromosome evolution. Current Opinion in Genetics & Development 16(6): 578-585.
-
Gilbert SF (2014) Developmental Biology. In: 10th (Edn.), Sunderland (MA). Yale J Biol Med 87(1): 93.
-
Olito C, Abbott JK (2025) The evolution of suppressed recombination between sex chromosomes and the lengths of evolutionary strata. Evolution79(7): 1371- 1385.
-
Kubiak JZ, Kloc M, Piprek RP (2020) History of The Research on Sex Determination. Biomedical Journal of Scientific & Technical Research 25(3): 19104-19112.
-
Eggers S, Sinclair A (2012) Mammalian sex determination- insights from humans and mice. Chromosome Res 20(1): 215-238.
-
Larson CL, Kopania EEK, Good JM (2018) Spermatogenesis and the evolution of mammalian sex chromosomes. Trends Genet 34(9): 722-732.
-
Hughes JF, Page D (2015) The Biology and Evolution of Mammalian Y Chromosomes. Annual review of genetics 49(1).
-
Morgan AP, de Villena FP (2017) Sequence and Structural Diversity of Mouse Y Chromosomes. Molecular Biology and Evolution 34(12): 3186-3204.
-
van der Bijl W, Mank JE (2025) Hiding in plain sight: the Y chromosome and its reinvigorated role in evolutionary processes. Evolution Letters 9(2): 165-171.
-
Soh YQS, Alfoldi J, Pyntikova T, Brown LG, Graves T, et al. (2014) Sequencing the mouse Y chromosome reveals convergent gene acquisition and amplification on both sex chromosomes. Cell 159(4): 800-813.
-
Cirulis A, Hansson B, Abbott JK (2022) Sex-limited chromosomes and non-reproductive traits. BMC Biol 20(156).
-
Rodrigues N, Studer T, Dufresnes C, Perrin N (2018) Sex- Chromosome Recombination in Common Frogs Brings Water to the Fountain-of-Youth. Molecular Biology and Evolution 35(4): 942-948.
-
Abbott JK, Nordén AK, Hansson B (2017) Sex chromosome evolution: historical insights and future perspectives. Proc Biol Sci 284(1854): 20162806.
-
Geffroy B, Besson M, Sánchez-Baizán N, Clota F, Goikoetxea A, et al. (2021) Unraveling the genotype by environment interaction in a thermosensitive fish with a polygenic sex determination system. Proc Natl Acad Sci 118 (50): e2112660118.
-
Peichel CL, McCann SR, Ross JA (2020) Assembly of the threespine stickleback Y chromosome reveals convergent signatures of sex chromosome evolution. Genome Biol 21(177).
-
Saunders PA, Muyle A (2024) Sex Chromosome Evolution: Hallmarks and Question Marks. Molecular Biology and Evolution 41(11): 218.
-
Landeen EL, Presgraves DC (2013) Evolution: From Autosomes to Sex Chromosomes — and Back. Current Biology 23(18): 848-850.
-
Nozawa M, Onizuka K, Fujimi M (2016) Accelerated pseudogenization on the neo-X chromosome in Drosophila miranda. Nat Commun 7: 13659.
-
Strickberger MW (1976) Genetics. Pearson Education. In: 2nd (Edn.), New York. Paperback Single-Issue Magazine.
-
Wheeler BS, Anderson JA, Presgraves DC (2016) Recombination suppression and the evolution of sex chromosomes in insects. PLoS Biology 14(5): e1002524.
-
Landeen EL, Presgraves DC (2013) Evolutionary dynamics of the X chromosome: Male-driven evolution, gene traffic, and the degeneration of the Y. Genetics 194(4): 943–955.
-
Chen X, Zhang J (2015) No X-chromosome dosage compensation in human proteomes. MolBiolEvol 32: 1456-1460.
-
Meyer BJ (2022) Mechanisms of sex determination and X-chromosome dosage compensation. Genetics 220(2): iyab197.
-
Lee H, Cho D, Whitworth C, Eisman R, Phelps M, et al. (2016) Effects of Gene Dose, Chromatin, and Network Topology on Expression in Drosophila melanogaster. PLoS Genet 12(9): e1006295.
-
Palmer DH, Rogers TF, Dean R, Wright AE (2019) How to identify sex chromosomes and their turnover, Mol Ecol 28(21): 4709-4724.
-
Duan J, Larschan EN (2019) Dosage Compensation: How to Be Compensated…Or Not?. Current Biology 29(23): R1229-R1231.
-
Ercan S, Dick LL, Lieb JD (2015) The role of the dosage compensation complex in balancing X-linked gene expression in Drosophila and Caenorhabditis elegans. Chromosome Research 23(3): 303-322.
-
Bhadra U, Gandhi SG, Palaparthi R, Balyan MK, Pal- Bhadra M (2016) Drosophila maleless gene counteracts X global aneuploid effects in males. FEBS J 283(18): 3457-3470.
-
Yuri S, Sergey U, Mikhail G, Stepan B, Sergey R (2022) Dosage Compensation in Drosophila: Its Canonical and Non-Canonical Mechanisms. International Journal of Molecular Sciences 23: 10976.
-
Meisel RP, Hart MLI, Vu BL, Bolden Q, Chen KT, et al. (2022) Genes relocated between Drosophila chromosomes evolve under selection pressures different from canonical dispersed duplicates. bioRxiv.
-
Strome S, Powers J, Dunn M, Korf I (2014) Sex chromosome regulation in C. elegans: Absence of spermatogenesis genes on the X chromosome. Developmental Cell 28(6): 674-684.
-
Meyer BJ (2022) The X chromosome in C. elegans sex determination and dosage compensation. Curr Opin Genet Dev 74: 101912.
-
Magnusson K, Lycett GJ, Mendes AM, Lynd A, Papathanos PA, et al. (2012) Demasculinization of the Anopheles gambiae X chromosome. BMC Evolutionary Biology 12: (69).
-
Ernst C, Eling N, Martinez-Jimenez CP (2019) Staged developmental mapping and X chromosome transcriptional dynamics during mouse spermatogenesis. Nat Commun 10: 1251.
-
Wei C, Kesner B, Yin H, Lee JT (2024) Imprinted X chromosome inactivation at the gamete-to-embryo transition. Molecular Cell 84(8): 1442-1459.
-
Luo Q, Zhang X, Zhou Q (2024) Sex chromosome regulation of gametogenesis: Comparative insights from ZW and XY systems. Trends in Genetics, Advance online publication.
-
Wang Q, Mank JE, Li J, Yang N, Qu L (2017) Allele-Specific Expression Analysis Does Not Support Sex Chromosome Inactivation on the Chicken Z Chromosome. Genome Biology and Evolution 9(3): 619-626.
-
Wapeesittipan P, Joshi A (2023) Integrated analysis of robust sex-biased gene signatures in human brain. Biol Sex Differ 14(1): 36.
-
Alfeghaly C (2025) X chromosome inactivation in mammals: general principles and species-specific considerations. EMBO Rep 26(14): 3478-3490.
-
Tenorio M, Cortez D (2022) X Chromosome Genomics, Reference Module in Life Sciences. Elsevier.
-
Abe H, Alavattam KG, Kato Y, Castrillon DH, Pang Q, et al. (2020) The initiation of meiotic sex chromosome inactivation sequesters DNA damage signaling from autosomes in Mouse spermatogenesis. Current Biology 30(21): 408-420.
-
Bracewell R, Bachtrog D (2020) Complex Evolutionary History of the Y Chromosome in Flies of the Drosophila obscura Species Group. Genome Biology and Evolution 12(5): 494-505.
-
Lucchesi JC, Kuroda MI (2015) Dosage Compensation in Drosophila. Cold Spring Harb Perspect Biol 7(5): a019398.
-
Vicoso B, Bachtrog D (2015) Numerous Transitions of Sex Chromosomes in Diptera. PLoS Biol 13(4): e1002078.
-
Meiklejohn CD, Presgraves DC (2012) Little evidence for demasculinization of the Drosophila X chromosome among genes expressed in the male germline. Genome Biology and Evolution 4(10): 1007-1016.
-
Xiong Y, Chen X, Chen Z, Wang X, Shi S, Wang X, et al. (2010) RNA sequencing shows no dosage compensation of the active X chromosome. Nat Genet 42(12): 1043- 1047.
-
Chen X, Oliver B (2015) X Chromosome and Autosome Dosage Responses in Drosophila melanogaster Heads.G3 (Bethesda) 5(6): 1057-1063.
-
Julien P, Brawand D, Soumillon M, Necsulea A, Liechti A, et al. (2012) Mechanisms and evolutionary patterns of mammalian and avian dosage compensation. PLoS Biol 10(5): e1001328.
-
Pessia E, Makino T, Bailly-Bechet M, McLysaght A, Marais GA (2012) Mammalian X chromosome inactivation evolved as a dosage compensation mechanism for dosage-sensitive genes on the X chromosome. Proc Natl Acad Sci USA, 109(14): 5346-5351.
-
Rahman MS, Pang MG (2020) New Biological Insights on X and Y Chromosome-Bearing Spermatozoa. Front Cell Dev Biol 7: 388.
-
Arlt MF, Kruger AN, Swanepoel CM, Mueller JL (2025) Reenacting a mouse genetic evolutionary arms race in yeast reveals that SLXL1/SLX compete with SLY1/2 for binding to Spindlins. PNAS 122(7): e2421446122.
-
Viera A, Parra MT, Arévalo S, de la Vega CG, Santos JL, et al. (2021) X Chromosome Inactivation during Grasshopper Spermatogenesis. Genes (Basel) 12(12): 1844.
-
Murat F, Mbengue N, Winge SB, Trefzer T, Leushkin E, et al. (2023) The molecular evolution of spermatogenesis across mammals. Nature 613(7943): 308-316.
-
Lin F, Xing K, Zhang J, He X (2012) Expression reduction in mammalian X chromosome evolution refutes Ohno’s hypothesis of dosage compensation. Proc Natl Acad Sci USA 109(29): 11752-11757.
-
McDonough Y, Ruzicka F, Connallon T (2024) Reconciling theories of dominance with the relative rates of adaptive substitution on sex chromosomes and autosomes. Proc Natl Acad Sci USA 121 (44): e2406335121.
-
Eyer PA, Blumenfeld AJ, Vargo EL (2019) Sexually antagonistic selection promotes genetic divergence between males and females in an ant. Proc Natl Acad Sci USA 116(48): 24157-24163.
-
Filatov DA (2022) Recent expansion of the non‐ recombining sex‐linked region on Silene latifolia sex chromosomes. J Evol Biol 35(12): 1696-1708.
-
Griffin RM, Le Gall D, Schielzeth H, Friberg U, Arnqvist G (2016) Within-population variation in sperm length is driven by haploid selection. Biology Letters 12(2): 20150913.
-
Parker DJ, Jaron KS, Dumas Z, Robinson‐Rechavi M, Schwander T (2022) X chromosomes show relaxed selection and complete somatic dosage compensation across Timema stick insect species. Journal of Evolutionary Biology 35(12): 1734-1750.
-
Krasovec M, Nevado B, Filatov DA (2018) A Comparison of Selective Pressures in Plant X-Linked and Autosomal Genes. Genes 9(5): 234.
-
Arbiza L, Gottipati S, Siepel A, Keinan A (2014) Contrasting X-Linked and Autosomal Diversity across 14 Human Populations. The American Journal of Human Genetics 94(6): 827-844.
-
Vegesna R, Tomaszkiewicz M, Medvedev P, Makova KD (2019) Dosage regulation, and variation in gene expression and copy number of human Y chromosome ampliconic genes. PLoS Genet 15(9): e1008369.
-
Ghenu AH, Bolker BM, Melnick DJ, Evans BJ (2016) Multicopy gene family evolution on primate Y chromosomes. BMC Genomics 17: 157.
-
Rogers TF, Pizzari T, Wright AE (2021) Multi-Copy Gene Family Evolution on the Avian W Chromosome. Journal of Heredity 112(3): 250-259.
-
Li Y, Park H, Smith TE, Moran NA (2019) Gene Family Evolution in the Pea Aphid Based on Chromosome-Level Genome Assembly. Mol Biol Evol 36(10): 2143-2156.
-
Vanaphan N, Dauwalder B, Rebecca A (2012) Zufall Diversification of takeout, a male-biased gene family in Drosophila. Gene 491(2): 142-148.
-
Vibranovski MD, Lopes HF, Karr TL, Long M (2009) Stage-specific expression profiling of Drosophila spermatogenesis suggests that meiotic sex chromosome inactivation drives genomic relocation of testis- expressed genes. PLoS Genetics 5(11): e1000731.
-
Meisel RP, Han MV, Liebeskind BJ (2012) Evolutionary dynamics of gene duplication in the Drosophila melanogaster genome. Genome Biology and Evolution 4(10): 986-1006.
-
Djordjevic J, Van PT, Toubiana W, Labédan M, Dumas Z, et al. (2025) Dynamics of X chromosome hyper-expression and inactivation in male tissues during stick insect development. PLoS Genet 21(3): e1011615.
-
Turner JMA (2007) Meiotic sex chromosome inactivation. Development 134(10): 1823-1831.
-
Jesus P, de la Fuente R, Marcia M, María P, Alberto V, et al. (2012) Inactivation or non-reactivation: What accounts better for the silence of sex chromosomes during mammalian male meiosis?. Chromosoma 121: 307-326.
-
Mrnjavac A, Khudiakova KA, Barton NH, Vicoso B (2023) Slower-X: reduced efficiency of selection in the early stages of X chromosome evolution. Evol Lett 7(1): 4-12.
-
Wright AE, Fumagalli M, Cooney CR, Bloch NI, Vieira FG, et al. (2018) Male-biased gene expression resolves sexual conflict through the evolution of sex-specific genetic architecture. Evolution Letters 2(2): 52-61.
-
Baird RB, Hitchcock TJ, Ševčík J, Monteith KM, Gardner A, et al. (2025) Faster adaptation but slower divergence of X chromosomes under paternal genome elimination. Nat Commun 16(1): 5288.
-
Alavattam KG, Maezawa S, Andreassen PR, Namekawa SH (2021) Meiotic sex chromosome inactivation and the XY body: a phase separation hypothesis. Cell Mol Life Sci 79(1): 18.
-
Strome S, Kelly WG, Ercan S, Lieb JD (2014) Regulation of the X chromosomes in the germ line and soma of Caenorhabditis elegans. Cold Spring Harbor Perspectives in Biology 6(3): a018366.
-
Griffin RM, Schielzeth H, Friberg U (2016) Autosomal and X-Linked Additive Genetic Variation for Lifespan and Aging: Comparisons Within and Between the Sexes in Drosophila melanogaster. G3 (Bethesda), 6(12): 3903- 3911.
-
Richardson SR, Doucet AJ, Kopera HC, Moldovan JB, Garcia-Perez JL, et al. (2015) The Influence of LINE-1 and SINE Retrotransposons on Mammalian Genomes. Microbiol Spectr 3(2): MDNA3-0061-2014.
-
Yoshioka H, Geyer CB, Hornecker JL, Patel KT, McCarrey JR (2007) In vivo analysis of developmentally and evolutionarily dynamic protein-DNA interactions regulating transcription of the Pgk2 gene during mammalian spermatogenesis. Mol Cell Biol 27(22): 7871-7885.
-
Meisel RP, Malone JH, Clark AG (2022) Disentangling the mechanisms of gene traffic from the X chromosome. Genome Biology and Evolution 14(6): evac072.
-
Vicoso B, Bachtrog D (2013) Reversal of an ancient sex chromosome to an autosome in Drosophila. Nature 499(7458): 332-335.
-
Turner JMA (2007) Meiotic sex chromosome inactivation. Nature Reviews Genetics 8(11): 822–832.
-
Avelino LS, Martins FR, Silva RJ (2025) Meiotic sex chromosome inactivation and the regulation of mammalian spermatogenesis. Developmental Biology, Advance online publication.
-
Lee JT (2005) Regulation of X-chromosome counting by Tsix and Xist. Science 309(5735): 768-771.
-
Heard E, Disteche CM (2006) Dosage compensation in mammals: Fine-tuning the expression of the X chromosome. Genes & Development 20(14): 1848-1867.
-
Turner JMA, Aprelikova O, Xu X, Wang R, Kim S, et al. (2004) BRCA1, histone H2AX phosphorylation, and male meiotic sex chromosome inactivation. Current Biology 14(23): 2135-2142.
-
Fernandez-Capetillo O, Mahadevaiah SK, Celeste A, Romanienko PJ, Camerini-Otero RD, et al. (2003) H2AX is required for chromatin remodeling and inactivation of sex chromosomes in male mouse meiosis. Developmental Cell 4(4): 497-508.
-
Rohozinski J, Bishop CE (2004) The mouse X-linked retrogene Utp14b is expressed in spermatids and is required for male fertility. Biology of Reproduction 70(3): 770-776.
-
Meisel RP, Malone JH, Clark AG (2022) Patterns of X chromosome gene traffic shaped by sex-biased gene expression during spermatogenesis. Evolution 76(2): 432-448.
-
Wheeler BS, Anderson E, Frokjer Jensen C, Bian Q, Jorgensen E, et al. (2016) Chromosome-wide mechanisms to decouple gene expression from gene dose during sex-chromosome evolution. ELife 5: e17365.
-
Ercan S (2015) Mechanisms of X Chromosome Dosage Compensation. J Genomics 3: 1-19.
-
Lyon MF (1999) X-chromosome inactivation. Current Biology 9(7): R235-R237.
-
Meisel RP, Asgari D, Schlamp F, Unckless RL (2022) Induction and inhibition of Drosophila X chromosome gene expression are both impeded by the dosage compensation. G3 Genes|Genomes|Genetics 12(9): 165.
-
Luo R, Zhang H, Xu J (2024) Sex-linked repression and gene distribution in avian genomes: Evidence from ovary-specific transcriptomes. Genome Biology and Evolution 16(5): 101.
-
Rice WR (1984) Sex chromosomes and the evolution of sexual dimorphism. Evolution 38(4): 735-742.
-
Khil PP, Camerini-Otero RD (2005) Genetic cross- talk between X and Y chromosomes in mammalian meiosis. PLoS Genetics 1(3): e54.
-
Huynh KD, LeeJ T (2003) Inheritance of a preinactivated paternal X chromosome in early mouse embryos. Nature 426(6968) 857-862.
-
Abe H Alavioon, G Li C, Dean R (2020) Meiotic drive and transmission ratio distortion in the context of spermatogenesis Philosophical Transactions of the Royal Society B Biological Sciences 375(1795): 20200067.
-
ArizmendiMejía R, Ledoux JB, Civit S Antunes, A Thanopoulou Z, Garrabou J, et al. (2015) Demographic responses to warming reproductive maturity and sex influence vulnerability in an octocoral. Coral Reefs 34(4): 1207-1216.
-
Holleley CE, Sarre SD, OMeally D, Georges A (2016) Sex reversal in reptiles reproductive oddity or powerful driver of evolutionary change Sex. Dev 10(56): 279-287.
-
Coyne JA (2018) Two rules of speciation revisited. Mol Ecol 27: 3749-3752.
-
Cowell F (2023) 100 years of Haldanes rule. J Evol Biol 36(2) 337-346.
-
Meisel RP, Connallon T (2013) The fasterX effect integrating theory and data Trends. Genet 29(9): 537- 544.
-
Mank JE, Vicoso B, Berlin S, Charlesworth B (2010) Effective population size and the FasterX effect Empirical results and their interpretation Evolution 64(3): 663- 674.
-
Janoušek V, Fischerová J, Mořkovský L, Reif J Antczak M, Albrecht T, et al. (2019) Postcopulatory sexual selection reduces Zlinked genetic variation and might contribute to the large Z effect in passerine birds. Heredity (Edinb) 122(5): 622-635.
-
Searl JB, de Villena FPM (2024) Meiotic Drive and Speciation. Annu Rev Genet 58(1): 341-363.
-
Jaenike J (2008) X chromosome drive Curr Biol 18(12): R508-511.
-
Kasimatis KR, SánchezRamírez S, Stevenson ZC (2021) Sexual Dimorphism through the Lens of Genome Manipulation Forward Genetics and Spatiotemporal Sequencing. Genome Biol Evol 13(2): 243.
-
Deegan DF, Engel N (2019) Sexual Dimorphism in the Age of Genomics How When Where. Front Cell Dev Biol 7: 186.
-
Pokorný Š, Pavlovič O, Kleisner K (2024) Sexual Dimorphism The Interrelation of Shape and Color. Arch Sex Behav 53(8): 3255-3265.
-
Mank JE (2009) Sex chromosomes and the evolution of sexual dimorphism lessons from the genome. Am Nat 173(2): 141-150.
-
Lüpold S, Pitnick S (2018) Sperm form and function what do we know about the role of sexual selection Reproduction 155(5): R229-R243.
-
Hann MC, Lau PE, Tempest HG (2011) Meiotic recombination and male infertility from basic science to clinical reality. Asian J Androl 13(2): 212-218.
-
Lockley EC, Eizaguirre C (2021) Effects of global warming on species with temperaturedependent sex determination Bridging the gap between empirical research and management. Evol Appl 14(10): 2361- 2377
-
Bhattacharya S, Sadhukhan D, Saraswathy R (2024) Role of sex in immune response and epigenetic mechanisms. Epigenetics & Chromatin 17(1).
-
Predescu DN, Mokhlesi M, Predescu SA (2022) The Impact of Sex Chromosomes in the Sexual Dimorphism of Pulmonary Arterial Hypertension. The American Journal of Pathology 192(4):582-594.
-
Wright AE, Fumagalli M, Cooney CR, Bloch NI, Vieira FG, et al. (2018) Malebiased gene expression resolves sexual conflict through the evolution of sexspecific genetic architecture. Evolution Letters 2(2): 52-61.
-
Alavattam KG, Maezawa S, Andreassen PR, Namekawa SH (2021) Meiotic sex chromosome inactivation and the XY body a phase separation hypothesis. Cell Mol Life Sci 79(1): 18.
-
Andjel L, Kratochvil L, Rovatsos M (2024) Comparison of dNdS ratios shows little evidence for fasterZ effect in Furcifer chameleons after controlling for genespecific evolutionary rates. The Journal of Heredity 116(5): 582-590.
-
Bachtrog D, Mank JE, Peichel CL, Kirkpatrick M, Otto SP, et al. (2014) Sex determination why so many ways of doing it?. PLoS Biol 12(7): e1001899.
-
Baird RB, Hitchcock TJ, Ševčík J, Monteith KM, Gardner A, et al. (2025) Faster adaptation but slower divergence of X chromosomes under paternal genome elimination. Nat Commun 16(1): 5288.
-
Betrán E, Thornton K, Long M (2002) Retroposed new genes out of the X in Drosophila. Genome Research 12(12): 1854-1859.
-
Betrán E, Thornton K, Long M (2002) Retroposed new genes out of the X in Drosophila Proceedings of the National Academy of Sciences of the United States of America 99(23): 14257-14262
-
Beukeboom LW, Perrin N (2014) The evolution of sex determination Oxford UK Oxford University Press
-
BravoEstupiñan DM, AguilarGuerrero K, Quirós S, Acón MS, MarínMüller C, et al. (2023) Gene dosage compensation Origins criteria to identify compensated genes and mechanisms including sensor loops as an emerging systemslevel property in cancer. Cancer Med 12(24): 22130-22155.
-
Courret C, ChingHo C, Wei K HC, MontchampMoreau C, Larracuente AM (2019) Meiotic drive mechanisms lessons from Drosophila Proc R Soc B 286(1913): 20191430.
-
Djordjevic J, Tran Van P, Toubiana W, Labédan M, Dumas Z, et al. (2025) Dynamics of X chromosome hyper- expression and inactivation in male tissues during stick insect development. PLoS Genet 21(3): e1011615.
-
Khil PP, Camerini Otero RD (2005) X inactivation in the mouse meiotic sex body is correlated with the absence of sex-biased genes. Genetics 171(1): 173-183.
-
Kousathanas A, Halligan DL, Keightley PD (2014) Faster-X adaptive protein evolution in House Mice. Genetics 196(4): 1131-1143.
-
Luo X, Guo J, Zhang J, Ma Z, Li H (2024) Overview of chicken embryo genes related to sex differentiation. Peer J 12: e17072.
-
Mank JE, Nam K, Ellegren H (2010) Faster-Z evolution is predominantly due to genetic drift. Mol Biol Evol 27(3): 661-70.
-
McCarrey JR, Watson C, Atencio J, Ostermeier GC, Marahrens Y, et al. (2002) X-chromosome inactivation during spermatogenesis is regulated by an Xist/Tsix- independent mechanism in the mouse. Genesis 34(4): 257-266.
-
Mrnjavac A, Khudiakova KA, Barton NH, Vicoso B (2023) Slower-X: reduced efficiency of selection in the early stages of X chromosome evolution. Evol Lett 7(1): 4-12.
-
Ohno S (1970) Evolution by Gene Duplication. London: Allen and Unwin.
-
O’Neill JJ, O’Neill RJ (2018) Sex chromosome repeats tip the balance towards speciation. Mol Ecol 27(19): 3783-3798.
-
Ono M, Harley VR (2013) Disorders of sex development: new genes, new concepts. Nat Rev Endocrinol 9(2): 79-91.
-
Page J, de la Fuente R, Manterola M, Parra M, Viera A, et al. (2012) Inactivation or non-reactivation: What accounts better for the silence of sex chromosomes during mammalian male meiosis? Chromosoma 121(3): 307-326.
-
Penny GD, Kay GF, Sheardown SA, Rastan S, Brockdorff N (1996) Requirement for Xist in X chromosome inactivation. Nature 379(6561): 131-137.
-
Richardson SR, Doucet AJ, Kopera HC, Moldovan JB, Garcia Perez JL, (2014) The Influence of LINE-1 and SINE Retrotransposons on Mammalian Genomes. Microbiol Spectr 3(2): MDNA3-0061-2014.
-
Riddle NC, Elgin SCR (2018) The Drosophila Dot Chromosome: Where Genes Flourish Amidst Repeats. Genetics 210(3): 757-772.
-
Rohozinski J, Bishop CE (2004) The mouse juvenile spermatogonial depletion (jsd) phenotype is due to a mutation in the X-linked gene Utp14b. Proc Natl Acad Sci USA 101(32): 11695-11700.
-
Vibranovski MD, Zhang YE, Long M (2009) General gene movement off the X chromosome in the Drosophila lineage. Genome Res 19(5): 897-903.
-
Wei Q, Yu X, Shi H, Chen Z, Luo M (2024) Editorial: Mammalian spermatogenesis: Genetic and environmental factors. Front Cell Dev Biol 12: 1372799.
-
Wei Z, Liu J, Wang J (2024) Evolutionary redistribution of spermatogenesis genes across the mammalian genome. Nat Commun 15: 4521.
-
Yoshioka H, Geyer CB, Hornecker JL, Patel KT, McCarrey JR (2007) In vivo analysis of developmentally and evolutionarily dynamic protein-DNA interactions regulating transcription of the Pgk2 gene during mammalian spermatogenesis. Mol Cell Biol 27(22): 7871-85.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind