Morphogenetic Characteristics of the European Roe Deer (Capreolus capreolus l.) From Ukraine
A study of the exterior and craniometric characteristics of European roe deer (n = 244) from Ukraine showed that animals from southern populations outperform their counterparts from northern areas in most indicators. This is observed in all age groups regardless of sex. Certain geographical differences were found between the south–eastern, south–western and Crimean populations. They were most noticeable in the forests of the Crimean mountains, where an isolated aboriginal centre of European roe deer has been preserved. Two well–differentiated haplogroups were identified in Ukraine, with a separate subgroup of closely related haplotypes in Crimea, which is a consequence of the long–term geographical isolation of the roe deer population on the Crimean peninsula.
Introduction
All wild animals are capable of living for long periods in their natural environment only if their biological needs are met by the ecological conditions. This principle of unity between the organism and the environment is supported by morphological and physiological adaptations, which are relatively constant over a certain period of time. Their formation in different organisms is ensured by variability, which is hereditary in nature. Since any population consists of territorial and social groups, interbreeding occurs more frequently within them than on the periphery. In isolated pockets, unique gene pools emerge, which are the product of random changes [1]. At the same time, the number of heterozygotes is decreasing everywhere [3], and therefore the reserve of genetic variability that could ensure adaptation to dynamic situations in the future becomes very limited over time [4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19].
In Ukraine, most mammal populations are somewhat isolated, despite the absence of significant geographical barriers. This is particularly true of the European roe deer, whose habitats have been quite distant from each other for a long time. Together with low numbers, this has significantly complicated genetic exchange between their groups. Other species also suffer from partial isolation, which is exacerbated by intensive agricultural use of most territories. This is manifested in the variability of craniological and external characteristics, the study of which reveals the degree of similarity and/or difference between intraspecific groups.
In recent years, climate warming has become noticeable in many places, manifested in an increase in the average annual temperature, a reduction in the duration of ice cover, the disappearance of blizzards, heavy snowfalls, etc. Of course, this has changed the living conditions of wild animals, which could not but affect their corresponding morphological response. Therefore, our goal was to study the exterior, craniological, and genetic characteristics of European roe deer in Ukraine.
Materials and Methods
Body and skull measurements (n = 244) were performed in accordance with the methods approved by the Coordination Council of Roe Deer Specialists of the Member Countries of the Council for Mutual Economic Assistance [6]. Age was determined based on tooth cement and tooth wear [17]. The study used collections from the National Museum of Natural History of the National Academy of Sciences of Ukraine (Kyiv), the Zoological Museum of Odessa National University (Odessa), our own trophies, and materials from colleagues. The data obtained were calculated using regression and cluster analysis. Clustering of similarity matrices was performed using the weighted pair group method [14]. To date, the craniology of European roe deer in Ukraine remains an understudied topic. With this in mind, we tried to collect a sufficiently large array of materials on this issue, which was divided into the following age groups: calves animals aged 5–10 months, yearlings 15–21 months, and adults 29-126 months. This division is related to the duration of the roe deer hunting season in Ukraine.
Molecular genetic analysis was performed on muscle tissue samples of roe deer preserved in alcohol, collected in eight regions of Ukraine. The control region and the cytochrome b gene of mitochondrial DNA were used as molecular genetic markers. A combined sequence of corresponding fragments of Capreolus pygargus mitochondrial DNA from the Krasnoyarsk region of Russia was used as an outgroup [20]. Median haplotype networks were constructed using the Network program [2]. The main approaches to processing and interpreting the material correspond to the ideas of E. Mayr [8].
In recent years, climate warming has become noticeable in many places, as evidenced by an increase in the average annual temperature, a reduction in the periods of ice cover, the disappearance of blizzards, heavy snowfalls, etc. Of course, this has changed the habitat of wild animals, which could not but affect their morphological response. Therefore, our goal was to study the external, craniological, and genetic characteristics of the European roe deer in Ukraine [20, 21].
Results
The Exterior of the European Roe Deer
In Ukraine, which is characterised by diverse natural conditions, the European roe deer has a number of interesting genetic and morphological features. Despite the sexual dimorphism in size and body weight characteristic of most ungulates, we were unable to detect such dimorphism in roe deer in the steppe zone (Table 1). Although males surpassed females in almost all parameters, these differences were not statistically significant, and in some cases completely coincided. Interestingly, in terms of most external indicators, roe deer from the steppe zone of Ukraine were similar to roe deer from Bulgaria [12], Germany [9] and the field population of Poland [8]. At the same time, when comparing our data with the results of a study of several animals (5♂ and 4♀) from the forest-steppe zone of Ukraine, it turned out that the latter surpassed our roe deer (males/females) in body length by 7.96/9.24, in height at the withers — by 0.62/10.33, in body circumference - by 7.70/7.35, and in limb length - by 8.82/10.56%. The body weight of the animals was also greater: males weighed 29.6±1.6 (24-33) kg, and females weighed 29.0±1.3 (26- 32) kg. This may be due to the peculiarities of the genotype structure of these animals, which have survived in the southern forest-steppe zone since ancient times, as well as the influence of larger Siberian roe deer. Judging by karyotypes and morphological indicators, four roe deer studied on the right bank of the Dnieper in the Black Forest (Kirovograd region) belong to the European species.
On the left bank, in a group from the Samara Forest (Dnipropetrovsk region), where Siberian roe deer lived at the beginning of the 20th century, two of the nine animals studied did not have additional chromosomes, which allows them to be considered European. However, three had one and four had two B chromosomes, which is characteristic of the Siberian species [6]. The influence of the small sample size, which consisted of adult and fairly large individuals, on the differences found cannot be ruled out. For comparison, in the Crimean Reserve, the body length of a male roe deer that died in August reached 117, the head length was 23.0, and the ear length was 12.4 cm. The body length of three females that died in February-March 1928 was 115.3±4.41 (109.8–124.0), head length — 22.4±0.78 (20.9–23.4), ear length — 13.0±0.42 (12.5–13.8), tail length — 2.2±0.15 (1.0–2.5) cm, height at the withers — 69.8±0.44 (69.0–75.7) cm, height at the croup — 77.6±1.27 (75.7–80.0) cm. Their average body weight was 16 kg, and the maximum-30 kg [5]. These indicators do not differ significantly from those of roe deer in continental Europe.
| Measurements | Sex | n | M±m | Min | Max | Std. Dev. | t |
|---|---|---|---|---|---|---|---|
| Body weight, kg | ♂ | 12 | 27.3±1.71 | 20 | 35.3 | 5.38 | 0.9 |
| ♀ | 14 | 25.1±2.36 | 18 | 34.1 | 6.38 | ||
| Body length, cm | ♂ | 11 | 115.6±4.66 | 90 | 144 | 16.31 | 0.1 |
| ♀ | 13 | 113.7±4.78 | 89 | 148 | 17.23 | ||
| Oblique length, cm | ♂ | 10 | 69.2±1.36 | 60 | 74 | 4.31 | 1.3 |
| ♀ | 10 | 64.3±3.34 | 47 | 78.1 | 10.57 | ||
| Head length, cm | ♂ | 11 | 25.1±0.51 | 21 | 26.8 | 1.7 | |
| ♀ | 10 | 25.1±0.56 | 22 | 28.5 | 1.94 | 0.3 | |
| Body circumference, cm | ♂ | 12 | 70.1±1.66 | 62 | 78 | 5.75 | 0.4 |
| ♀ | 12 | 70.8±3.29 | 58 | 79.9 | 11.14 | ||
| Height at withers, cm | ♂ | 10 | 80.1±2.05 | 69 | 91.5 | 6.14 | 1.3 |
| ♀ | 13 | 75.5±2.18 | 61 | 88.9 | 7.87 | ||
| Height at sacrum, cm | ♂ | 10 | 87.2±1.44 | 80 | 92 | 4.55 | 1.9 |
| ♀ | 12 | 79.9±2.55 | 64 | 91.4 | 8.83 | ||
| Paw length, cm | ♂ | 11 | 29.4±0.32 | 28 | 31.2 | 1.05 | 1.9 |
| ♀ | 6 | 28.0±0.65 | 26 | 29.9 | 1.59 | ||
| Foot length, cm | ♂ | 12 | 36.3±1.54 | 28 | 44.5 | 5.33 | 0.6 |
| ♀ | 10 | 36.0±1.56 | 25 | 43.5 | 4.93 | ||
| Tail length, cm | ♂ | 9 | 3.3±0.37 | 2.2 | 5.1 | 1.1 | 0.3 |
| ♀ | 12 | 2.9±0.31 | 2 | 6 | 1.07 | ||
| Ear length, cm | ♂ | 11 | 13.4±0.28 | 12 | 15.5 | 0.92 | 0.4 |
| ♀ | 13 | 13.2±0.37 | 11 | 15.1 | 1.33 |
Table 1: Body size and weight of European roe deer from the steppe zone (14 November–13 February).
In western Ukraine, from the Carpathians to the Black Sea, roe deer with a very similar phenotype live: many individuals have a transverse light brown stripe on their necks (Figure 1a- c). Representatives of the Crimean population do not have it. The special morphology of animals from the mountain forests of Crimea was noted as early as the beginning of the 20th century by hunters [5], who reported that two forms of European roe deer inhabit the peninsula: a dark one, called ‘black’, and an animal with a white neck. It is believed that the Crimean population was formed as a result of the mixing of white–necked animals, which came to Crimea via a land bridge in the Pliocene from the Caucasus and Asia Minor, with later dark immigrants who penetrated from continental regions in the Quaternary period.
However, we have never encountered black roe deer in the mountain forests of Crimea — only occasionally have we seen individuals with dark heads and muzzles (Figure 1d).

Craniometric Characteristics of the European Roe Deer
In the first year of life, sexual dimorphism in the size of the skull of the European roe deer is weakly expressed, which is also observed in other populations [4, 8, 19]. Despite this, male calves outperform females in all indicators, although reliable differences between them have been recorded in only four of them. These are: interorbital width (t=2.16 at P = 0.03), facial length (t=2.09 at P=0.05), nasal bone length (t=3.10) and lower jaw diastema length (t=2.35 at P=0.02). Moreover, most craniometric indicators have low individual variability, which is clearly evident from the small coefficient of variation. Among them, the zygomatic width, cerebral capsule width and lower jaw length are particularly stable in both sexes. The greatest variability was found in females in the length of the lower row of molars and the diastema of the lower jaw (Table 2).
| Measurements | Sex | Age groups of animals | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Calves (20 ♂; 32 ♀) | Yearlings (18 ♂; 28 ♀) | Adults (71 ♂; 58 ♀) | ||||||||
| M±m | Min | Max | M±m | Min | Max | M±m | Min | Max | ||
| Total cranial length | ♂ | 18.5±0.25 | 16 | 20 | 19.9±0.24 | 19 | 21 | 20.9±0.10 | 19 | 23 |
| ♀ | 18.2±0.17 | 17 | 20 | 20.0±0.15 | 20 | 20.4 | 20.6±0.11 | 19 | 22 | |
| Cоndylobasal length | ♂ | 17.4±0.28 | 15 | 19 | 18.8±0.24 | 17 | 20 | 19.6±0.11 | 18 | 21 |
| ♀ | 17.0±0.19 | 15 | 18 | 19.0±0.20 | 18 | 19.5 | 19.5±0.12 | 15 | 21 | |
| Basal length | ♂ | 16.3±0.30 | 14 | 17 | 17.5±0.23 | 16 | 18.7 | 18.4±0.10 | 16 | 20 |
| ♀ | 15.9±0.16 | 15 | 18 | 17.7±0.17 | 17 | 17.9 | 18.2±0.10 | 16 | 20 | |
| Maximum width of the skull | ♂ | 8.2±0.06 | 7.6 | 9.1 | 8.9±0.09 | 8.4 | 9.6 | 9.4±0.05 | 8.5 | 10 |
| ♀ | 8.0±0.10 | 5.7 | 8.7 | 8.8±0.09 | 8.4 | 9.3 | 9.0±0.05 | 8.4 | 9.8 | |
| Zygomatic breadth | ♂ | 7.9±0.04 | 7.4 | 8.4 | 8.4±0.05 | 8 | 8.9 | 8.8±0.04 | 8.1 | 9.5 |
| ♀ | 7.9±0.05 | 7.4 | 8.4 | 8.4±0.04 | 8.2 | 8.6 | 8.6±0.04 | 8 | 9.1 | |
| Interorbital constriction | ♂ | 4.8±0.05 | 4.4 | 5.7 | 5.3±0.07 | 4.8 | 5.6 | 5.5±0.05 | 4.5 | 6.6 |
| ♀ | 4.7±0.05 | 4.3 | 5.2 | 5.1±0.06 | 4.8 | 5.4 | 5.2±0.04 | 4.7 | 5.9 | |
| Length of the frontal part of the skull | ♂ | 9.4±0.16 | 8.4 | 10 | 10.3±0.21 | 9.3 | 11.5 | 10.8±0.09 | 9.4 | 14 |
| ♀ | 8.9±0.13 | 8.1 | 10 | 10.4±0.06 | 10 | 10.6 | 10.9±0.07 | 9.7 | 12 | |
| Maximum length of the nasal | ♂ | 5.4±0.08 | 4.6 | 6.1 | 6.1±0.13 | 4.9 | 6.8 | 6.4±0.06 | 5.2 | 7.4 |
| ♀ | 5.1±0.07 | 4.2 | 5.8 | 6.2±0.12 | 5.9 | 6.9 | 6.4±0.08 | 5.4 | 7.7 | |
| Length of the top row of molars | ♂ | 5.3±0.08 | 4.6 | 6.5 | 5.9±0.08 | 5.2 | 6.4 | 6.0±0.03 | 5.4 | 6.8 |
| ♀ | 5.3±0.06 | 4.6 | 6.1 | 5.8±0.11 | 5.1 | 6.3 | 5.9±0.05 | 5.1 | 6.8 | |
| Width of the cerebral capsule | ♂ | 5.7±0.05 | 5.1 | 6.2 | 6.1±0.07 | 5.6 | 6.9 | 6.3±0.03 | 5.8 | 6.8 |
| ♀ | 5.7±0.04 | 5.3 | 6.3 | 6.0±0.05 | 5.8 | 6.3 | 6.1±0.03 | 5.7 | 6.7 | |
| Occipital condyle breadth | ♂ | 3.7±0.08 | 3.2 | 4.2 | 3.9±0.08 | 3.5 | 4.3 | 3.9±0.04 | 3.2 | 4.4 |
| ♀ | 3.6±0.05 | 3.2 | 3.9 | 3.7±0.04 | 3.4 | 3.7 | 3.7±0.04 | 3 | 4.3 | |
| Mandibule length | ♂ | 14.5±0.14 | 13 | 16 | 15.8±0.15 | 15 | 17 | 16.7±0.08 | 15 | 18 |
| ♀ | 14.3±0.13 | 13 | 16 | 16.2±0.16 | 16 | 17.3 | 16.4±0.10 | 15 | 19 | |
| Length of the lower row of molars | ♂ | 5.6±0.10 | 4.9 | 6.7 | 6.7±0.09 | 5.5 | 7.2 | 6.7±0.04 | 6 | 7.6 |
| ♀ | 5.5±0.09 | 4.6 | 6.9 | 6.5±0.11 | 5.6 | 6.9 | 6.5±0.06 | 4.5 | 7.2 | |
| Diastema of the mandible length | ♂ | 4.0±0.06 | 3.5 | 4.5 | 4.3±0.06 | 3.6 | 4.6 | 4.6±0.05 | 3.3 | 5.5 |
| ♀ | 3.7±0.09 | 2.8 | 4.9 | 4.5±0.08 | 4.1 | 4.8 | 4.6±0.06 | 3.8 | 5.8 |
Table 2: Age-related variability of craniometric indicators (cm) in European roe deer.
Naturally, we are aware that the group of yearlings included animals that differed in age by almost two times. Therefore, the indicated superiority of males over females only indicates faster growth in the former of the specified sections of the skull, which, in this particular case, is not perceived by us as an evolutionarily formed difference. This may be due to the appearance of special protrusions on the frontal bones of males, which are the stumps of future horns, since during their formation, the entire adjacent part of the skull undergoes changes. In calves, the internal distance between them was 1.9±0.07 (1.11–2.72) cm, and the external distance was 4.3±1.10 (3.20–5.89) cm.
In the second year, the situation changes dramatically, and females surpass males in half of the craniological indicators (Table 2). However, a significant difference is still observed in males, but only in the interorbital width (t=2.29 at P=0.03), which was also the case in calves. In one- year-old males, compared to calves, there is a decrease in the internal distance between the horn stumps (1.7±0.14; limit: 0.90–2.48) and an increase in the external distance (4.6±0.19; limit: 3.21–6.02) cm. In general, in this age group, the earlier superiority of males over females in terms of the size of individual parts of the skull is completely levelled out. However, in many cases, a decrease in the coefficient of variation is observed in one-year-old males. The exceptions are: the length of the facial section, the length of the nasal bones, the width of the cerebral capsule, and the length of the diastema of the lower jaw.
At the same time, in one-year-old females, compared to calves, the coefficient of variation decreases across all craniometric indicators. Taken together, this indicates a change in the growth rate of various parts of the roe deer skull and an increase in secondary sexual dimorphism. This is accompanied by a change in craniometric proportions in animals of different sexes and the superiority of males over females in many respects. For example, in adult roe deer, not only is there a noticeable increase in craniometric indicators in skull size compared to one-year-old individuals (Table 2), but there is also a significant increase in males. Starting at the age of 2.5 years, males significantly exceed females in 8 out of 14 measurements. Particularly large differences are observed in the maximum, interorbital and zygomatic widths. In addition, adult male European roe deer significantly exceed females in maximum length, width of the occipital condyles, as well as in the width of the cerebral capsule (at P = 0.03), length of the lower jaw (at P = 0.05) and length of the lower row of molars (at P < 0.01). No statistically significant differences in skull size between adult males and females were found in terms of such indicators as condylar–basal and main length, as well as the length of the upper row of teeth, nasal bones, facial part and diastema of the lower jaw. Moreover, although the latter two indicators did not reach statistically significant values in females, they were even slightly larger than in males. It should be noted that in two roe deer of different sexes, obtained in 1996 in the north of the Odessa region, the size of the lower jaw diastema was very significant. In a male aged ~80 months, it was 60.4 mm, and in a female aged ~103 months, it was 64.4 mm, which is characteristic of Siberian roe deer, not European roe deer. These indicators differed so much from the others that we were forced to exclude them from further calculations.
After reaching sexual maturity and further maturation, the formation of antler stubs ends in male roe deer. In the steppe region of Ukraine, the internal distance between them in adult individuals was 1.63±0.04 (1.0-2.7), and the external distance was 5.6±0.08 (4.0-6.8) cm. This is close to roe deer from France and the Republic of Belarus [6]. In general, it should be noted that the variability of craniometric characteristics in roe deer in the steppe zone of Ukraine is insignificant. It is highest in calves, as evidenced by the coefficient of variation (6.10±0.40, limit = 3.19-13.37); it is slightly lower in adults (5.35±0.34, limit = 3.25-8.87), and lowest in yearlings (4.32±0.34, limit = 1.42-8.37). At the same time, judging by the extreme values, many craniometric indicators are capable of a significant dynamic response to changes in the ecological situation.
Given the wide distribution of the European roe deer and its importance for European hunting, the main features of its ecology and morphology, in particular the variability of craniometric indicators, have been well studied. An in-depth analysis of the latter issue revealed significant similarities between representatives of the Polish, Moravian and Czech populations and the predominance of individuals from Germany and Hungary [11]. The reasons for this phenomenon are considered to be differences in soil characteristics, nutrition, climate, parasitofauna, population density, protection conditions and even the organisation of hunting. However, they are all inferior to roe deer living in Sweden [10] and Lithuania [4]. Therefore, we decided to compare the results of our studies conducted in the southern part of the European roe deer range with the results of studies in the Baltic countries, where this species lives in the unique conditions of the northern taiga (Table 3).
The analysis showed that in their first year of life, animals from the steppe zone of Ukraine surpass the largest roe deer in Europe. Particularly large differences were found in animals of both sexes in terms of maximum skull length, face length and cerebral capsule width. The significant advantage of young males from the steppe zone of Ukraine over their northern counterparts is also noticeable in the width of the zygomatic bone, the width of the occipital protuberance, and the length of the protuberance base (at P = 0.03). At the same time, young females from the north have a slightly, though not significantly, greater width of the zygomatic bone than roe deer from the south of the range. However, we do not attach great importance to all the differences found, as they may be due to the different ages of the animals being compared.
Although Table 3 analyses data on 6-month-old roe deer, it is unrealistic to distinguish between hunted young animals by age with an accuracy of 1 month. Therefore, roe deer samples from both Lithuania and Ukraine may include individuals that are close in age but still differ by 1–2 months. Given the different birth dates and the characteristics mentioned above, they naturally differ in both appearance and skull size. However, the skull features of young roe deer from southern populations, which exceed those of their northern counterparts, are of particular interest. Therefore, we tried to compare animals from these groups at mature age, after the growth rate of all skull elements had slowed down. Among adult roe deer, females and males from Lithuania reliably and significantly exceed representatives from southern Ukraine only in terms of zygomatic bone width. Surprisingly, in the female group, Ukrainian representatives significantly exceed representatives from the north in terms of maximum and condyle–basal skull length (at P = 0.03), length of the facial part and upper row of molars, as well as width of the cerebral capsule.
| Measurements | Part of the range | M±m | CV, % | t | M±m | CV, % | t |
|---|---|---|---|---|---|---|---|
| Males (6 months old) | Females (6 months) | ||||||
| Total cranial length | North | 175.3±1.03 | 3.5 | 175.9±1.12 | 3.2 | ||
| South | 181.2±0.14 | 3.7 | 5.68 | 182.8±0.26 | 5.1 | 6 | |
| Cоndylobasal length | North | 164.6±1.05 | 3.8 | 166.3±1.18 | 3.6 | ||
| South | 167.5±0.19 | 5.2 | 2.72 | 170.3±0.30 | 5.2 | 3.3 | |
| Maximum width of the skull | North | 76.9±0.51 | 3.9 | 79.9±0.52 | 3.2 | 2.1 | |
| South | 78.5±0.05 | 3.3 | 3.12 | 78.8±0.05 | 3 | ||
| Interorbital constriction | North | 45.9±0.32 | 4.1 | 47.2±0.40 | 4.2 | ||
| South | 46.7±0.04 | 4.7 | 2.48 | 47.7±0.05 | 4.9 | 1.2 | |
| Length of front part | North | 85.8±0.73 | 5 | 86.2±0.72 | 4.2 | ||
| South | 89.1±0.12 | 5.8 | 4.46 | 92.2±0.17 | 5.5 | 8.1 | |
| Width of the cerebral capsule | North | 55.1±0.22 | 2.3 | 54.9±0.36 | 3.3 | ||
| South | 57.1±0.04 | 3.5 | 8.94 | 57.4±0.05 | 4.7 | 6.9 | |
| Width of the occipital condyles | North | 34.9±0.29 | 5 | 36.0±0.44 | 5.9 | ||
| South | 36.2±0.05 | 5.7 | 4.42 | 36.3±0.06 | 5.8 | 0.7 | |
| Males (6.5 years and older) | Females (6.5 years and older) | ||||||
| Total cranial length | North | 204.3±1.13 | 3.2 | 206.5±1.78 | 2.9 | ||
| South | 207.7±0.15 | 3.5 | 2.98 | 207.7±0.15 | 3.5 | 0.7 | |
| Cоndylobasal length | North | 191.7±1.30 | 2.9 | 194.7±1.51 | 2.6 | ||
| South | 194.9±0.22 | 5.7 | 2.43 | 194.8±0.22 | 5.7 | 0.1 | |
| Maximum width of the skull | North | 90.4±0.85 | 4 | 5.05 | 95.6±0.81 | 2.8 | 12 |
| South | 86.1±0.06 | 3.5 | 86.1±0.06 | 3.5 | |||
| Interorbital constriction | North | 52.1±0.66 | 5.4 | 56.5±0.75 | 4.4 | 4.7 | |
| South | 52.9±0.05 | 5.1 | 1.21 | 53.0±0.04 | 5.1 | ||
| Length of front part | North | 106.2±0.73 | 2.9 | 105.7±1.39 | 4.3 | ||
| South | 109.5±0.10 | 4.7 | 4.48 | 109.5±0.10 | 4.7 | 2.7 | |
| Length of the upper row of molars | North | 55.1±0.54 | 3.3 | 55.4±0.40 | 3.1 | ||
| South | 57.6±0.07 | 6.8 | 4.59 | 57.6±0.07 | 6.8 | 5.4 | |
| Width of the cerebral capsule | North | 60.1±0.45 | 3.2 | 62.6±0.48 | 2.5 | 2.1 | |
| South | 61.6±0.04 | 3.6 | 3.32 | 61.6±0.04 | 3.6 | ||
| Width of the occipital condyles | North | 36.5±0.33 | 3.9 | 39.2±0.33 | 2.8 | 6.9 | |
| South | 36.9±0.05 | 6.1 | 1.2 | 36.9±0.05 | 6.1 |
Table 3: Craniometric measurements (mm) of roe deer calves from the north (Lithuania; n = 99) [4] and south (Ukraine; n = 110) of
In addition, among males, Lithuanian roe deer significantly exceed Ukrainian roe deer in terms of interorbital width of the skull (t = 4.7) and width of the occipital condyles (t = 6.9), but are inferior to them in terms of the length of the upper row of molars (t = 4.7) and the length of the facial part (P = 0.03). The difference in the values of other craniometric characteristics is insignificant. Judging by the coefficient of variation, both in Lithuania and Ukraine, despite age-related characteristics, craniometric indicators in young and adult roe deer are characterised by low variability, 4.3±0.19 and 4.1±0.23, respectively (t = 0.76 at P = 0.45). This is due to the insignificant spread of values of craniological indicators in each of the variation series, which is biologically determined by the fairly complete realisation of hereditary qualities, despite the uneven growth of all elements of the skull. A similar pattern is observed in southern groups of roe deer, whose natural habitats have been preserved in the mountainous part of Crimea, northern regions of Odessa region, and Moldova. Despite the geographical remoteness of the lowland populations, no significant differences in craniological characteristics have been found between them in the steppe zone (Figure 2). The absence of differences may be not only a consequence of their genetic relatedness, but also the result of the interaction of the genotype with homogeneous ecological conditions in the steppe zone, as well as the mixing of animals during the last surge in population size. In general, the dynamics of craniometric characteristics of European roe deer in the southern marginal groups indicate that they have reached the state of their ancestral populations. However, in the mountain-forest areas of Crimea, a special form according to Wright’s principle [17] is observed, which is characteristic of island fauna and geographical isolates.
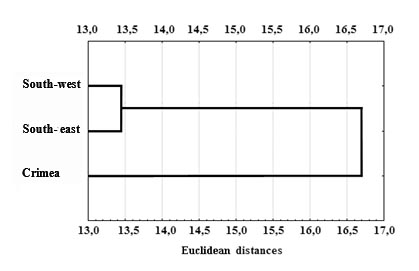
Our comparison of craniometric indicators between adult individuals from continental and mountain roe deer groups also revealed a number of interesting differences (Table 4). In particular, males from the lowland populations of Ukraine significantly exceed those from the mountainous Crimea in the following skull characteristics: maximum length (at P = 0.01), condylo–basal length (at P = 0.02), width of the cerebral capsule (at P = 0.01) and occipital condyles (at P = 0.04), length of the lower jaw (at P = 0.01) and its diastema between the first premolar (Pm1) and canine (C) of the lower jaw. In females from continental Ukraine, a significant superiority was found in such indicators as: main length (at P = 0.01) and length of the diastema of the lower jaw (at P = 0.01). In contrast, adult females of Crimean roe deer were significantly larger than those from the continental south only in terms of the length of the lower row of molars (at P = 0.01). Judging by our limited data, in the Crimean roe deer population, unlike in continental Ukraine, adult males significantly exceed females only in two craniometric indicators. These are the interorbital width (t = 2.61 at P = 0.01) and the length of the nasal bones (t = 2.24 at P = 0.03). This indicates a difference in the proportions of the skulls of roe deer in this group compared to representatives of continental populations [15].
| Measurements, cm | Sex | Continental South (71♂; 58♀) | Crimean Peninsula (21♂; 13♀)* | t | ||||
|---|---|---|---|---|---|---|---|---|
| M±m | Min | Max | M±m | Min | Max | |||
| Maximum length | ♂ | 20.92±0.10 | 19 | 23 | 20.32±0.18 | 19 | 22 | 2.8 |
| ♀ | 20.60±0.11 | 19 | 22 | 20.26±0.20 | 20 | 22 | 1.4 | |
| Condylo-basal length | ♂ | 19.60±0.11 | 18 | 21 | 19.05±0.19 | 17 | 20 | 2.4 |
| ♀ | 19.46±0.12 | 15 | 21 | 18.97±0.09 | 19 | 20 | 1.9 | |
| Main length | ♂ | 18.35±0.10 | 16 | 20 | 17.98±0.17 | 19 | 22 | 1.7 |
| ♀ | 18.22±0.10 | 16 | 20 | 17.58±0.12 | 17 | 19 | 3.1 | |
| Maximum width | ♂ | 9.41±0.05 | 8.5 | 10 | 9.29±0.08 | 8.6 | 9.2 | 1.2 |
| ♀ | 9.01±0.05 | 8.4 | 9.8 | 8.93±0.08 | 8.5 | 9.4 | 0.8 | |
| Zygomatic width | ♂ | 8.76±0.04 | 8.1 | 9.5 | 8.94±0.05 | 8.6 | 9.2 | 2 |
| ♀ | 8.56±0.04 | 8 | 9.1 | 8.50±0.25 | 6.4 | 9.1 | 0.4 | |
| Interorbital width | ♂ | 5.53±0.04 | 4.5 | 6.6 | 5.43±0.08 | 5 | 6.4 | 1.1 |
| ♀ | 5.22±0.04 | 4.7 | 5.9 | 5.16±0.04 | 4.9 | 5.5 | 0.8 | |
| Length of the facial part | ♂ | 10.82±0.09 | 9.4 | 14 | 10.54±0.17 | 9.9 | 12 | 1.2 |
| ♀ | 10.86±0.07 | 9.7 | 12 | 10.58±0.08 | 10 | 11 | 1.7 | |
| Length of the nasal bones | ♂ | 6.36±0.06 | 5.2 | 7.4 | 6.25±0.10 | 5.8 | 6.9 | 0.8 |
| ♀ | 6.42±0.08 | 5.4 | 7.7 | 6.26±0.10 | 5.7 | 6.9 | 1 | |
| Length of the upper row of molars | ♂ | 5.95±0.03 | 5.4 | 6.8 | 5.92±0.06 | 5.4 | 6.4 | 0.4 |
| ♀ | 5.88±0.05 | 5.1 | 6.8 | 5.86±0.11 | 5.3 | 6.8 | 0.2 | |
| Width of the cerebral capsule | ♂ | 6.25±0.03 | 5.8 | 6.8 | 6.07±0.08 | 5.3 | 6.6 | 2.7 |
| ♀ | 6.13±0.03 | 5.7 | 6.7 | 6.00±0.08 | 5.3 | 6.4 | 1.9 | |
| Width of the occipital condyles | ♂ | 3.87±0.04 | 3.2 | 4.4 | 3.68±0.05 | 3.5 | 3.9 | 2.2 |
| ♀ | 3.73±0.04 | 3 | 4.3 | 3.58±0.03 | 3.4 | 3.7 | 1.8 | |
| Length of the lower jaw | ♂ | 16.66±0.08 | 15 | 18 | 16.16±0.15 | 15 | 17 | 2.5 |
| ♀ | 16.41±0.10 | 15 | 19 | 16.04±0.13 | 15 | 17 | 1.8 | |
| Length of the lower row of teeth | ♂ | 6.68±0.04 | 6 | 7.6 | 6.77±0.11 | 6 | 7.3 | 1 |
| ♀ | 6.49±0.06 | 4.5 | 7.2 | 6.88±0.12 | 5.6 | 7.6 | 2.9 | |
| Length of the diastema of the lower jaw | ♂ | 4.59±0.06 | 3.3 | 6 | 4.17±0.10 | 3.5 | 4.9 | 3.2 |
| ♀ | 4.65±0.08 | 3.8 | 6.7 | 4.19±0.07 | 3.8 | 4.7 | 3 |
Table 4: Comparative characteristics of skull sizes in adult roe deer (29–126 months).
Genotypic Characterisation of the European Roe Deer
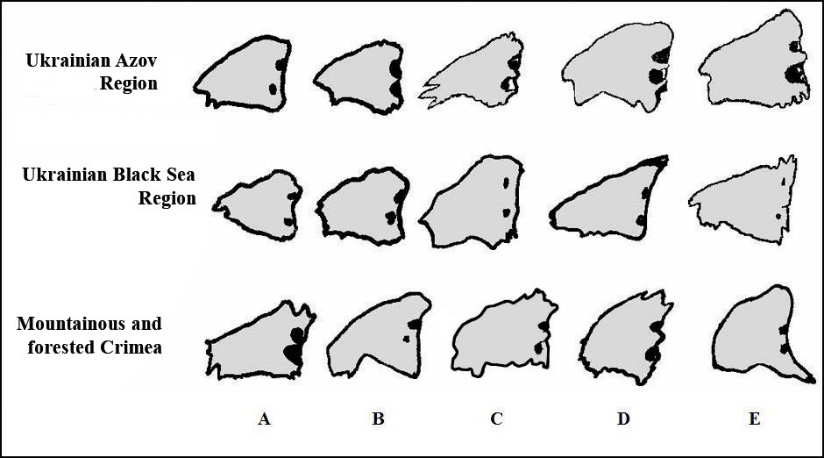
When comparing roe deer from different populations, we used a genotype marker such as the shape of the os lacrimale (Figure 3). Despite significant individual polymorphism, individuals from the mountain forests of Crimea are the most unique [15]. No roe deer with a similar shape of the lacrimal bone scale were found in any region of southern Ukraine. If we consider only animals from Crimea, it should be noted that roe deer with phenotype A (62.50%) are most common in the forests near Sevastopol, individuals with phenotype B are significantly less common (18.75%), and those with phenotypes C, D and E are very rare (6.25% each).
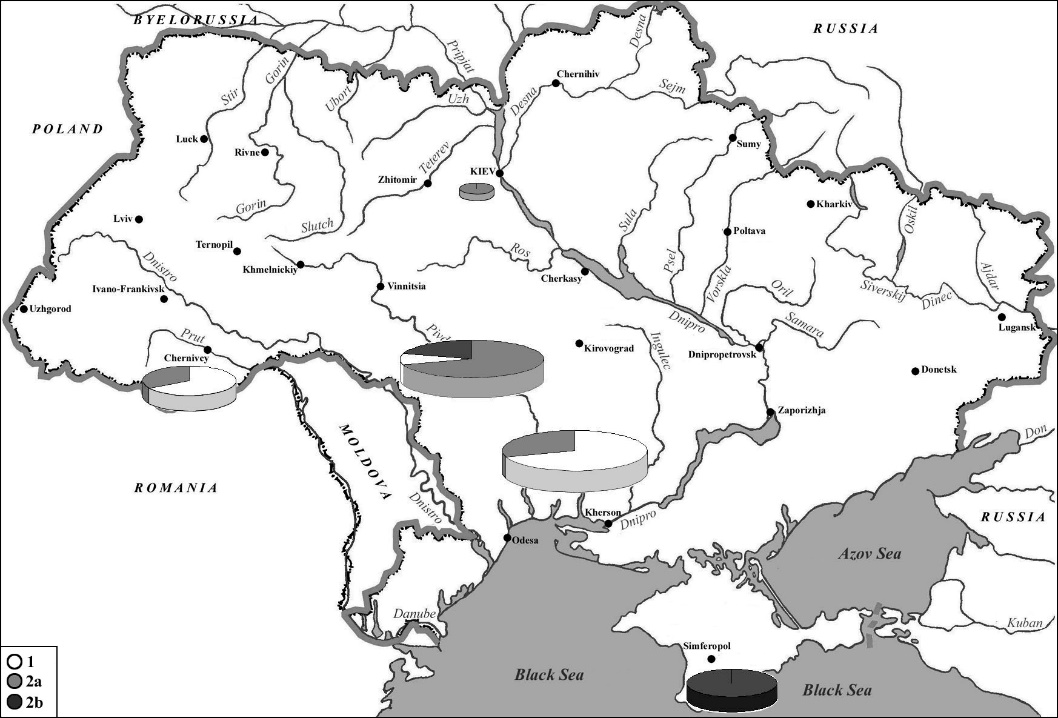
The prolonged geographical isolation of the Crimean roe deer population has influenced its genotype. Our studies of their genetic polymorphism in different parts of Ukraine, based on a median haplotype network, have shown that the animals belong to two well-separated haplogroups (Figure 4). Haplogroup 1 was found in the Lviv, Odessa, Nikolaev, Chernivtsi and Ternopil regions; haplogroup 2a — in the Chernivtsi, Odessa, Kiev, Vinnitsa and Ternopil regions; haplogroup 2b was found in the Zaporizhzhia, Chernivtsi, and Vinnytsia regions and in Crimea [21].
It should be noted that the Crimean roe deer population consisted exclusively of individuals belonging to haplogroup 2b, which indicates its taxonomic uniqueness. In other words, there are significant morphological and genetic differences between the Crimean and continental populations of European roe deer in Ukraine, which have been isolated from each other for a long time. For a long time, this species was completely absent from the steppe regions of the Azov and Black Sea coasts, as their territory did not provide suitable ecological conditions for roe deer to live.
After the formation of southern populations of European roe deer in Ukraine, which took place only in 1972–1975 [14, 15], the lack of natural forests, low density of artificial forest plantations, significant impact of wolves and poaching became a major obstacle to the normal exchange of migrants between continental and Crimean populations.
Conclusion
The inability to regularly exchange genes has led to the formation of a noticeable divergence between the steppe and mountain-forest populations of the European roe deer on the southern border of its range in Ukraine. The significant morphological and genetic characteristics of the Crimean population of the European roe deer, taking into account phylogeny, are sufficient to assign it the status of a subspecies, for example, Capreolus capreolus tauricus.
Acknowledgements
The following individuals provided significant assistance in researching European roe deer skulls: Yevhen Pysanets, Doctor of Biological Sciences, Professor (National Museum of Natural History of the National Academy of Sciences of Ukraine, Kyiv) and Volodymyr Lobkov, Doctor of Biological Sciences, Professor (Zoological Museum of Odessa National University, Odessa). I would like to thank all the hunters, gamekeepers and hunting experts who assisted in collecting tissue samples for analysis, but especially: Gennadiy Boiko, Sergey Samokhin and Yuriy Tkachuk.
References
-
Ayala FH (1981) Mechanisms of Evolution. Evolution. World pp: 35-65.
-
Bandelt HJ, Forster P, Rohl A (1999) Median-Joining networks for inferring intraspecific phylogenies Mol Biol 16(1): 37-48.
-
Berry RJ (1977) Variability in mammals. Basic concepts and problems. Advances in modern teriology Nauka pp: 5-25.
-
Bluzma PP (1974) Morphology of the skull of the Lithuanian roe deer (Capreolus capreolus). Zoological Journal 53(2): 263-271.
-
Dal SK (1930) Materials on the systematics and biology of the Crimean roe deer. Proceedings of the Crimean Society of Naturalists 12: 64-127.
-
Danilkin AA (1999) Deer (Mammals of Russia and neighbouring countries). GEOS pp: 1-552.
-
Mayr E (1971) Principles of Zoological Systematics. World pp: 1-454.
-
Frunziński B, Kaluziński J, Baksalary J (1982) Weight and body measurements of forest and field roe deer. Acta theriol 27(33): 479-488.
-
Stubbe G, Bruchholz Z (1979) Experiments on the hybridisation of European and Siberian roe deer. Zoological Journal 58(9): 1398-1403.
-
Essen L (1966) Das Rehwild in Sweden. Beiträge zur Jagd & Wild forschung 5(90): 143-147.
-
Kratochvil Z, Kux Z (1984) Kraniometrische Untersuchungen an Rehgeissen. Prirodověd pr Ustavu ČSAV 18(12): 1-625.
-
Petrov P, Dragoev P, Kolev I (1968) The Roe Deer in Bulgaria. Zemizdat pp: 1-236.
-
Sokal RR, Sneath PHA (1963) Principles of numerical taxonomy. WH Freeman & Co, San Francisco, USA, pp: 1-359.
-
Volokh AM (2014) Hunting Animals of the Ukrainian Steppe. Book 1. FLP Grin DS Kherson, pp: 1-412.
-
Volokh AM (2016) Hunting Animals of the Steppe Ukraine. Book 2. FLP Grin DS Kherson, pp: 1-573.
-
Volokh AM (2022) Determining the age of game animals. Oldi+, India, pp: 1-374.
-
Wright S (1943) Isolation by distance. Genetics 28: 114- 138.
-
Wright S (1977) Evolution and the genetics of poрulations. Experimental results and evolutionary deductions. Univ Chicago press 3: 1-613.
-
Zeida J, Zdeňka B (1985) Home ranges of field roe deer. Priridovĕd pr Ústavú ČSAV Brnĕ 19(1): 1-48.
-
Zvychayna EYu, Danilkin AA, Kholodova MV, Sipko TP, Berber AP (2011) Analysis of variability in the control region and cytochrome b mtDNA gene of the Siberian roe deer Capreolus pygargus Pall. Izvestiya RAN. Biology Series 1a No 5: 511-517.
-
Zvychayna EYu, Volokh AM, Kholodova MV, Danilkin AA (2013) Mitochondrial DNA Polymorphism of the European Roe Deer, Capreolus capreolus (Artiodactyla, Cervidae) from the South-West of Ukraine. Vestnyk zoologii Т 47(5): 415-420.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind