Outfitting COVID-19: An Effective Therapeutic Approach
Use of antisense oligonucleotides of the type 3′-(N)x-AAAUUUG-(N)x-5′ against slippery sequence and polynucleotides against pseudoknots forming sequences of SARS-CoV-2 RNA would block the first translation of ORF1a and ORF1b and hence dwindle the virus replication. It is easy to synthesize and deliver the antisense oligonucleotides to the target by directly injecting the nano formulation into the blood.
Introduction
COVID-19 public-health emergency is caused by the outbreak of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2). While it is important to investing time to search the origins of the pandemic it is the prime need of the moment to find out remedy against the outbreak and it is the molecular and biochemical mechanisms of multiplication of the virus would help us better than any other ways [1, 2, 3].
Current Therapeutic Approaches
Angiotensin-converting enzyme II (ACE2) expression is enhanced by SARS-CoV-2 infection [4]. Repression of ACE2 gene by EZH2-mediated H3K27me3 modifications of ACE2 promoter could be a targeted for prevention and adjuvant therapy of COVID-19. ACE inhibitors or angiotensin receptor blockers would be helpful for patients to fight COVD-19 [4, 5]. Non-specific medicines, including antimalarial and broad spectrum antibiotics are being used in many clinics. That the convalescent plasma (CP) would benefit COVID-19 patients along with antivirals is emerging. However, without control subject it is not clear yet although within three days of CP therapy patients exhibited improved clinical symptoms CP was given within two weeks of symptom onset [6, 7, 8].
Mechanism of Synthesis of Virus Subparticles
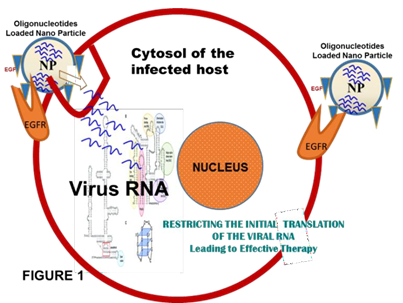
Prevailing knowledge based on studies with viral replication processes is; after the entry into the host cell, translation of ORF 1a and 1b into polyproteins Pp1a (4382 aas) and Pp1ab (7073 aas) is the primary function of the guest genome. Both the polyproteins then cleaved into fifteen non-structural proteins (nsps), which assemble and form the replication-transcription complex (RTC). ORF1b is translated by ribosome shifting one nucleotide in the −1 direction, from the ORF1a reading frame into ORF1b reading frame (reviewed in Tan, et al. [9]. This mechanism of repositioning is facilitated by two RNA elements; (i) a 5′-UUUAAAC-3′ heptanucleotide slippery sequence [10], and (ii) RNA pseudoknot structure (Figure 1) [11]. After the formation of RTC the full-length positive strand of genomic RNA transcribes to form a full-length negative- strand template for the synthesis of new genomic RNAs and overlapping subgenomic negative-strand templates. These subgenomic mRNAs are then transcribed and translated to produce the structural and accessory proteins. Several heterologous nuclear ribonucleoproteins (hnRNA) family members (hnRNPA1, PTB, SYN-CRYP) have been found to be essential for efficient RNA replication [9, 11, 12].
Formation of the cap structure of eukaryotic host and virus mRNAs basically requires three successive enzymatic reactions [13]. Removal of the the γ-phosphate group from the 5′-triphosphate end (pppN) of the newly transcribed mRNA chain to generate the diphosphate 5′-ppN by RNA 5′-triphosphatase (RTP) is initial reaction. Then, RNA- guanylyltransferase transfers a GMP to the 5′-diphosphate end to produce the cap core structure (GpppN). Finally, guanine-N7 methyltransferase (GNMT) methylates the attached GMP (capping) at the N7 position to produce a cap- 0 structure (me7GpppN). The 2′-O of ribose of the first and second nucleotides of the mRNA in higher eukaryotes and their viruses are additionally methylated. The responsible enzyme is ribose 2′-O-MT to form cap-1 and cap-2 structures, respectively Furuichi Y, et al. [14], Ferron F, at al [15]; (Figure 1). Ribose 2′-O-methylation of viral RNA cap provides a mechanism for viruses to escape host immune recognition [16, 17, 18].
Cap-0 reaction is mediated by the C-terminal domain of CoV nsp14 using S-Adenosylmethionine (SAM) as the methyl group donor (Figure 1) [19]. Conversion of cap-0 to cap-1 structures involves nsp16 that acts as a 2′-O-MT and forms a complex with nsp10 that appears to be required for efficient binding to SAM and the RNA substrate. Interestingly, SARS-CoV nsp10 plays an essential role in the specific binding of nsp16 to m7GpppA-capped RNA (first nucleotide is adenine). The crystal structure of the heterodimer of nsp16/nsp10 with bound methyl donor SAM showed that nsp10 may stabilize the SAM-binding pocket and extend the RNA-binding groove of nsp16 [20, 21]. Thus, blocking the translation of the virus SARS-CoV-2 mRNA definitely serves the purpose. Use of non-specific drugs would complicate the disease keeping the virus in dormant stage with a chance to relapse by re-activation of the pathogen and outbreak of the disease. Efficacy of Chloroquine (CQ) and a less toxic derivative of CQ, Hydroxychloroquine (HCQ) against malaria provoked some research groups around the globe and in many clinics, it has been in use against COVID-19.
Recommendation for Target Specific Convenient Therapy
In view of this, and to obtain toxicity free therapy of COVID-19 patients my recommendation is to conjointly use: (i) A oligonucleotide against the 5′-UUUAAAC-3′ heptanucleotide slippery sequence, and (ii) dismantling the pseudoknot preventing the pairing of matching sequences/RNA dimerization [22]. Oligonucleotide of the antisense-type 3′-(N)x-AAAUUUG-(N)x-5′ [where N is any one of the A, U, G, C and x = 8 to 10] will block the slippery sequence and antisense oligonucleotides against pseudoknots forming sequences may stop the translation of the viral RNA by ribosome shifting [22, 23, 24]. The polynucleotides can easily be design synthesize and delivered by well-tailored nanoparticle carriers. Ligands like EGF/folate attached with nanoparticles surface (oligonucleotides loaded) would easily be delivered to the affected tissues by ligand-receptor complex formation where there is overexpression of the cognate receptors, including EGFR/CD44 and subsequent endocytosis will bring them into the sites of action [25, 26]. Figure 1 for the overall procedure in brief.
It could be tested within seven days against lung cancer cell line or animal model.
Conflict of Interests
I declare that there is no conflict of interest.
References
-
Forster P, Forster L, Renfrew C, Forster M (2020) Phylogenetic network analysis of SARS-CoV-2 genomes. Proc Natl Acad Sci U S A 117(17): 9241-9243.
-
Zhou P, Yang XL, Wang XG, Hu B, Zhang L, et al. (2020) A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 579(7798): 270-273.
-
Andersen KG, Rambaut A, Lipkin WI, Holmes EC, Garry RF (2020) The proximal origin of SARS-CoV-2. Nat Med 26(4): 450-452.
-
Li Y, Li H, Zhou L (2020) EZH2-mediated H3K27me3 inhibits ACE2 expression. Biochem Biophys Res Commun 526(4): 947-952.
-
Talreja H, Tan J, Dawes M, Supershad S, Rabindranath K, et al. (2020) A consensus statement on the use of angiotensin receptor blockers and angiotensin converting enzyme inhibitors in relation to COVID-19 (corona virus disease 2019). N Z Med J 133(1512): 85- 87.
-
Abbasi J (2020) The promise and peril of antibody testing for COVID-19. JAMA 323(19): 1881-1883.
-
Shen C, Wang Z, Zhao F, Yang Y, Li J, et al. (2020) Treatment of 5 critically ill patients with COVID-19 with convalescent plasma. JAMA 323(16): 1582-1589.
-
Duan K, Liu B, Li C, Zhang H, Yu T, et al. (2020) Effectiveness of convalescent plasma therapy in severe COVID-19 patients. Proc Natl Acad Sci USA 117(17): 9490-9496.
-
Tan YW, Hong W, Liu DX (2012) Binding of the 5′-untranslated region of coronavirus RNA to zinc finger CCHC-type and RNA-binding motif 1 enhances viral replication and transcription. Nucleic Acids Res 40(11): 5065-5077.
-
Brierley I, Meredith MR, Bloys AJ, Hagervall TG (1997) Expression of a coronavirus ribosomal frameshift signal in Escherichia coli: influence of tRNA anticodon modification on frameshifting. J Mol Biol 270(3): 360- 373.
-
Brierley I, Pennell S, Gilbert RJC (2007) Viral RNA pseudoknots: versatile motifs in gene expression and replication. Nature Reviews Microbiology 5(8): 598-610.
-
Wu CH, Chen PJ, Yeh SH (2014) Nucleocapsid phosphorylation and RNA helicase DDX1 recruitment enables coronavirus transition from discontinuous to continuous transcription. Cell Host Microb 16(4): 462- 472.
-
Furuichi Y, Muthukrishnan S, Tomasz J, Shatkin AJ (1976) Mechanism of formation of reovirus mRNA 5′-terminal blocked and methylated sequence, m7GpppGmpC. J Biol Chem 251(16): 5043-5053.
-
Furuichi Y, Shatkin AJ (2000) Viral and cellular mRNA capping: past and prospects. Adv Virus Res 55: 135-184.
-
Ferron F, Subissi L, Silveira De Morais AT, Sevajol M, Gluais L, et al. (2018) Structural and molecular basis of mismatch correction and ribavirin excision from coronavirus RNA. Proc Natl Acad Sci U S A 115(2): E162-E171.
-
Daffis S, Szretter KJ, Schriewer J, Li J, Youn S, et al. (2010) 2′-O methylation of the viral mRNA cap evades host restriction by IFIT family members. Nature 468(7322): 452-456.
-
Zust R, Cervantes-Barragan L, Habjan M, Maier R, Neuman BW, et al. (2011) Ribose 2′-O-methylation provides a molecular signature for the distinction of self and non-self mRNA dependent on the RNA sensor Mda5. Nat Immunol 12: 137-143.
-
Nakagawa K, Lokugamage KG, Makino S (2016) Viral and cellular mRNA translation in coronavirus-infected cells. Adv Virus Res 96: 165-192.
-
Chen Y, Cai H, Pan J, Xiang N, Tien P, Ahola T (2009) Functional screen reveals SARS coronavirus nonstructural protein nsp14 as a novel cap N7 methyltransferase. Proc Natl Acad Sci USA 106(9): 3484-3489.
-
Chen SC, Olsthoorn RC (2010) Group-specific structural features of the 5′-proximal sequences of coronavirus genomic RNAs. Virology 401(1): 29-41.
-
Chen Y, Su C, Ke M, Jin X, Xu L, et al. (2011) Biochemical and structural insights into the mechanisms of SARS coronavirus RNA ribose 2′-O-methylation by nsp16/ nsp10 protein complex. PLoS Pathog 7: e1002294.
-
Ishimaru D, Plant EP, Sims AC, Yount BL, Roth BM, et al. (2013) RNA dimerization plays a role in ribosomal frameshifting of the SARS coronavirus. Nucleic Acids Res 41(4): 2594-2608.
-
Staple DW, Butcher SE (2005) Pseudoknots: RNA Structures with Diverse Functions. PLoS Biol 3(6): e213.
-
Dawson WK, Fujiwara K, Kawai G (2007) Prediction of RNA pseudoknots using heuristic modeling with mapping and sequential folding. PLoS One 2(9): e905.
-
Kim YD, Park TE, Singh B, Maharjan S, Choi YJ, et al. (2015) Nanoparticle-mediated delivery of siRNA for effective lung cancer therapy. Nanomedicine 10(7): 1165-1188.
-
Draz MS, Fang BA, Zhang P, Hu Z, Gu S, et al. (2014) Nanoparticle-mediated systemic delivery of siRNA for treatment of cancers and viral infections. Theranostics 4: 872-892.
- Solution-Processed Chiral Perovskites for Biomedical Applications
- Nanotechnology in Health Chemistry and Medicine: Current Challenges and Future Directions
- Human Exposure to Micro- and Nanoplastics: Pathways, Toxicity, and Intervention Strategies
- Exosome Nanomedicine for Cancer Therapy
- Micro and Nanoplastics–Plastisphere, Biotoxicity, Impact on Human Health, and Mitigation Strategies
- Process Validation of Cefixime Powder for Suspension Dosage Form, 50 mL