Microbiome and Neurotrauma: Emerging Innovations
The gut-brain axis plays an important role in bidirectional communication that exists and can be altered by injury. Neurotrauma provides acute alteration in the GI tract and alters autonomic function. In this focused review, we highlight what is known about GI disruption following neurotrauma. We then delve into how this affects recovery. Areas of innovation and emerging pre-clinical results are addressed. Finally, we address the link between neurotrauma induced GI dysfunction and progression to neurodegenerative disease states.
Introduction
The established neuroscience paradigm called the “gut-brain axis” is a method of describing and interpreting the bidirectional communication that exists between the brain and the microbiota of the digestive system [1, 2]. The physical connection between the two entities exists as the gastrointestinal (GI) organs are highly innervated by the central nervous system (CNS) and much enteric signaling originates from the gut [3, 4]. Emerging research suggests that the health of the CNS is modulated by these interactions between gut microbiota and neurotransmitters [1, 5, 6]. The gut-brain axis is becoming more relevant to investigations regarding the biological basis of neurodevelopmental and neurodegenerative disorders [6, 7]. Also, it has been studied extensively for its potential utility in regard to understanding the physiology of mental illness [1]. As traumatic head injuries have become an increasingly relevant public health issue, there have been new investigations into how the gut-brain axis may be altered because of neurotrauma and subsequent recovery. In fact, according to pre-clinical research, structural and functional damage to the GI tract is seen following head injuries, such as traumatic brain injury (TBI) [3, 8]. Potential mechanisms mediating this phenomenon are beginning to be elucidated, including reduction in short-chain fatty acid (SCFA)- producing microbiota in the gut post-TBI [9]. In addition, evidence that post-TBI alterations in gut microbiota may be sustained long-term has also begun to emerge [10]. The gut-brain axis may provide insight into a microbiota- based model to support recovery following neurotrauma and, in doing so, provide insight into innovative treatment modalities to target dysfunction in the gut-brain axis following neurotrauma.
Materials and Methodology
The electronic databases PubMed/MEDLINE, ScienceDirect, and Google Scholar were searched for original articles spanning from 1982 to 2022, with an emphasis on recent articles published between 2012 and 2022. All articles were in English language and the searches were performed in June and July of 2022. Original articles searched and ultimately included are comprised of comparative studies, retrospective cohort studies, case reports, systematic reviews, other related review articles, observational studies, randomized control trials, and clinical trials.
Neurotrauma and the Enteric Nervous System
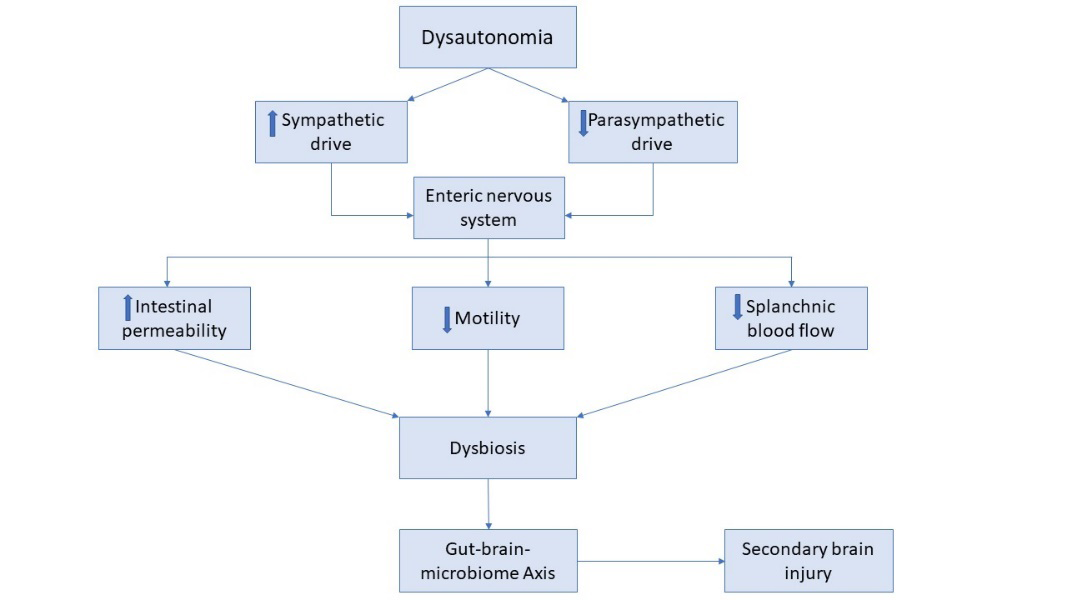
The CNS exists in close communication with the enteric nervous system (ENS) via a growing appreciation for the bidirectional gut-brain axis. Following neurotrauma, a systemic stress and inflammatory response occurs which has been proposed to disrupt this dynamically orchestrated axis and contribute to dysbiosis [11, 12]. Perturbations within the autonomic nervous system (ANS), which directly communicates with the ENS through the gut-brain axis, is thought to play a key role in TBI-associated intestinal dysfunction [11, 13]. The ANS is comprised of a sympathetic and parasympathetic branch and an imbalance between these two divisions may lead to the autonomic dysfunction commonly seen in TBI patients [14]. Although the ENS can function independently, its activity is extensively modulated by the ANS and, thus, a dysregulated ANS has potential to significantly alter ENS activity. The vagus nerve, a key neural pathway of the parasympathetic nervous system, innervates the gut from the esophagus to mid-colon and is a key modulator of secretomotor GI function [15]. Originating in the nucleus ambiguus and the dorsal motor nucleus in the brainstem, vagal fibers ultimately synapse directly onto the ENS plexuses to promote secretions and motor activity [11, 15]. On the other hand, post-ganglionic sympathetic fibers originating from prevertebral ganglia, synapse onto ENS neurons and generally serve to inhibit GI secretomotor function [11, 15].
Acetylcholine and norepinephrine are two key neurotransmitters that modulate parasympathetic and sympathetic pathways, respectively [8, 16]. Acetylcholine serves as both the pre- and post-ganglionic neurotransmitter in the parasympathetic nervous system and is essential to vagal nerve activity. Currently, there is substantial evidence for chronic changes in central cholinergic function following TBI [17, 18]. Cholinergic activity may likewise be disrupted in the peripheral nervous system following TBI, contributing to ENS dysfunction [8]. Increased intestinal permeability and decreased GI motility are two common secondary GI findings in TBI patients and have often been linked to perturbations in vagal activity [3, 11]. For example, in experimental studies, prophylactic vagal nerve stimulation has potential to preserve gut integrity and increase expression of tight junction proteins [3, 19]. Moreover, parasympathetic input onto the ENS via the vagus nerve is known to promote GI motility and, thus, a decrease in cholinergic activity following TBI may explain slowed GI motility seen in these patients [20]. Finally, cholinergic activity within the ENS appears to play an important role in regulating anti-inflammatory responses within the gut, potentially via interactions with α7 receptors on macrophages [21, 22]. In effect, a disrupted cholinergic system may not only alter intestinal structure and function, but may also serve to promote intestinal inflammation, further disrupting the intra-intestinal homeostasis.
An increase in sympathetic activity is similarly observed following TBI and is associated with a rise in systemic catecholamine levels, such as norepinephrine and epinephrine [12, 23]. Norepinephrine is the postganglionic neurotransmitter of the sympathetic nervous system and is one of the catecholamines largely responsible for end-organ sympathetic input to the ENS [24]. Although not yet directly studied in the setting of TBI, norepinephrine has been shown to slow gut motility and delay gastric emptying in animal studies and may likewise contribute to GI dysmotility seen in TBI patients [11, 16, 25]. In addition, catecholamines may also indirectly impact the gut-brain axis through a decrease in splanchnic blood flow [16, 20]. In particular, the gut relies heavily on blood flow to regulate GI secretions and nutrient absorptions and, thus, such alterations can significantly perturb intra-intestinal homeostasis [20, 26].
Altogether, disruptions in the ANS may lead to important changes in intestinal structure and function, such as disrupted intestinal barrier integrity, dysmotility and decreased blood flow. Such changes may disrupt the delicate homeostasis within the gut which maintains the microbiotic environment [11]. As a result, gut microbiota may intrinsically adapt, leading to dysbiosis. In a positive feedback loop, a dysregulated microbiotic environment may utilize the gut-brain-axis to then feedback onto the brain, enhancing neuroinflammation and secondary brain injury (Figure 1) [3, 27].
Innovation
Several innovative treatment approaches for optimization of the GI interface with the CNS post-TBI are either currently being studied or have been hypothesized as beneficial based on previously demonstrated research. As previously outlined, gut dysbiosis is triggered by neurotrauma. There are numerous subsequent consequences of dysbiosis, including compromised mucosal barrier function, intraluminal translocation of bacteria, and dysfunctional immune responses in the gut, which may play a multifactorial role in several disease processes [28] (Figure 2).

The dysbiosis of the small intestine altering the immune homeostasis causes an increase in regulatory T (Treg) cells and a decrease in interleukin 17 due to decreasing the response of γδ T cells through changes in the activity of dendritic cells [29]. The presence of gut microbiota has been demonstrated in studies of germ-free mice to help maintain gut motility, protect the integrity of the intestinal mucosa, and facilitate the differentiation of Treg cells [30]. It follows, then, that the restoration of gut microbiota to a baseline microbial profile following TBI may improve outcomes. Thus, fecal microbiota transplantation (FMT) (Figure 3) has been proposed as a method to restore the microbiota in this manner [31].
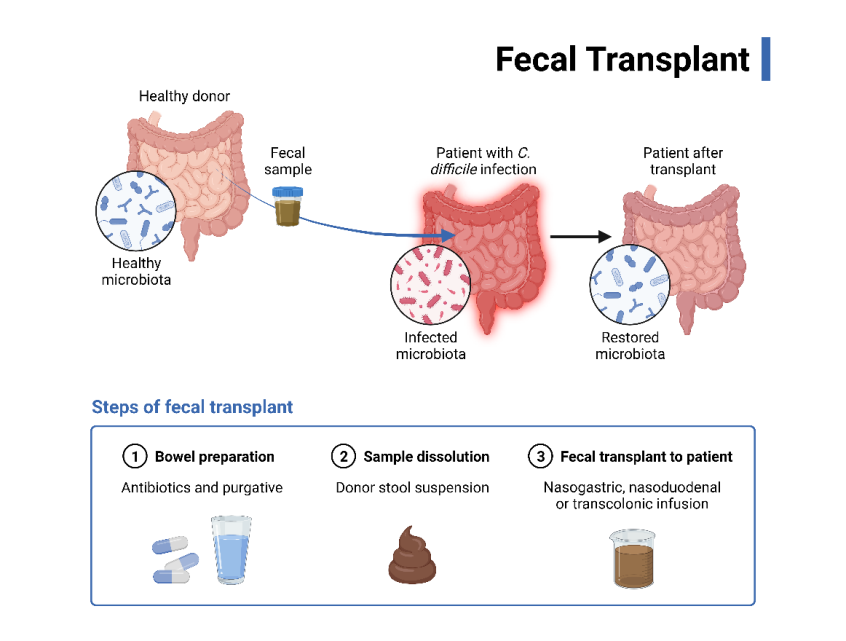
The use of FMT in treating Clostridium difficile infection has already been well described. It has also shown promise in treating other noninfectious diseases, such as inflammatory bowel disease [32]. It has been demonstrated that FMT may contribute to the reduction in the size of brain lesions and may also improve health outcomes following ischemic stroke, potentially by way of neuroprotection offered by FoxP3+ Treg cells in the gut after stroke in mice [33]. Another mechanism of neuroprotection in this manner has been proposed to be mediated by increased microglial activity [34]. Although in the primitive stages of research, FMT post- TBI is a leading innovative approach related to this topic as it currently stands, due to the demonstrated observation of neuroprotection.
Another innovation that could theoretically contribute to reduction in gut dysbiosis post-TBI is the administration of probiotics. There is evidence that probiotics causes an increase in interleukin-10 levels, thereby decreasing pro-inflammatory cytokines in the intestinal epithelium [35]. Specifically, probiotic formulations often consist of Lactobacillus and Bifidobacterium species of bacteria. These species are more likely to enhance Treg activity in the CNS and peripheral nervous system [28]. Bacterial metabolites produced by probiotic bacteria could also influence mitochondrial function [36]. This may be a direct or indirect result of increased generation of SCFA products by gut microorganisms [37]. In addition to dietary ketones, products of the gut microbiome provide the injured brain with alternative energy sources and may contribute to improved bioenergetic performance following TBI [38]. More specifically, butyric acid produced by Clostridium butyricum has been shown to improve neurological impairments, decrease brain edema, reduce neurodegeneration, and improve blood-brain barrier (BBB) function [39]. In addition, treatment with probiotic supplements has also been shown to lower plasma levels of TNF-α, reduce cerebral monocyte infiltration, and decrease microglial activation [40, 41]. Thus, it appears logical that administration of probiotics may afford a multifactorial benefit in terms of reduction of gut dysbiosis post-TBI that is an important consideration for future research in this setting.
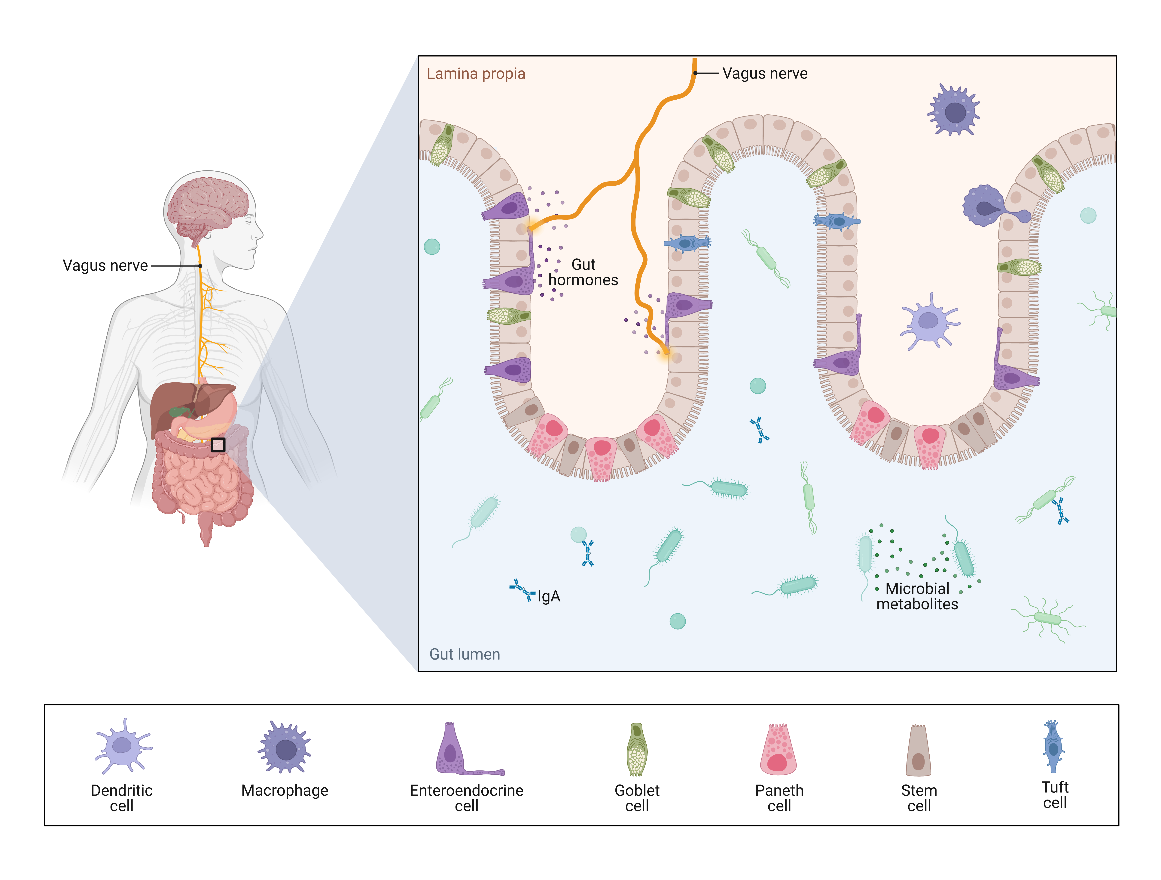
In addition, innovative treatment options targeting dysautonomia, which leads to dysbiosis and secondary brain damage after TBI, may also be effective treatment options that merit consideration. For example, vagal stimulation in the post-TBI setting may reduce intestinal permeability, by way of increased hormone production in the gut (Figure 4). As another possible avenue of therapy, targeted immunomodulatory strategies may also be explored. There is evidence that γδT cells may contribute to worsening neurologic recovery. In murine studies, monoclonal antibody therapy was used to inactivate γδT cells, which led to improved functional neurologic recovery [42]. Thus, it may be possible to develop individualized therapies by suppressing this damage-inducing cell line, although this assertion has not been substantiated by current research.
Preclinical Studies
Preclinical models of neurotrauma are fundamental to the understanding of the disease process given the challenges of studying neurotrauma in a clinical setting. The lack of accurate and dependable methods of diagnosis, both emergently and retrospectively, makes it difficult to select diseased participants and healthy controls reliably [43, 44]. It is also challenging to maintain a longitudinal clinical study in which cognitive and neurological symptoms can be analyzed while controlling all potentially confounding variables, such as the use of narcotics or psychiatric diagnoses [45, 46]. Thus, researchers have relied on preclinical models to study the onset, progression, and potential therapies for neurotrauma in a controlled environment [47]. The relationship between neurotrauma, the gut microbiome, and the gut-brain axis has been investigated across numerous preclinical models [48, 49]. In particular, researchers are exploring the bidirectional communication of the gut-brain axis to understand both enteric and neurological disease pathophysiology, and to elucidate potential treatments for disease processes such as Neurotrauma [3, 50].
Preclinical evaluation of neurotrauma and the gut- brain axis usually includes integrating an animal model of neurotrauma, such as the fluid percussion model or a blast-induced trauma model, and some modification or manipulation of the gut Microbiome [47, 51]. The modulatory effects that the gut microbiome has on the brain have been primarily studied using animals with no microbial species (germ-free), animals with fecal microbiota transplantation, and animals depleted of microbes using antibiotics [3, 51, 52, 53, 54]. These studies have provided evidence of the “bottom- up” mechanistic effects that the gut microbiota has on the regulation of various processes within the CNS, including modulation of pain and response to stress [3, 55, 56].
While there is still much work to be done in preclinical studies to understand the specific mechanisms involved, recent studies have started to reveal the influence that gut microbiota has on the gut-brain axis after a neurotraumatic event. Simon et al. used a cortical impact model of TBI on mice with and without administration of an enteric antibiotic cocktail [48]. The researchers concluded that the depletion of the gut microbiota with antibiotics has a neuroprotective effect post-TBI. Mice that were given the antibiotic cocktail before the controlled TBI showed increased CA1 hippocampal neuronal density, reduced Iba-1 positive cells, and reduced associative learning deficit, in comparison to the mice that were not given the antibiotic cocktail [48]. However, researchers at the St. Louis school of medicine completed a similar study showing that modulation of the gut microbiota with antibiotics before TBI results in increased neuronal loss and worsened outcomes [57]. These contrasting results could be due to the use of different antibiotics in the studies, resulting in different changes to the microbiota populations of each group [48, 57]. Although the results of these studies differ, they both show preclinical evidence of changes in the microbiota inducing changes in TBI sequelae. There have also been preclinical investigations that have revealed changes in the gut and fecal microbiota as a result of traumatic neural injury [58, 59]. This recent information supports the observations of Houlden et al., who used an experimental weight drop model to study post-TBI dysbiosis in mice [60]. They found changes in the gut microbiota populations of Porphyromonadaceae, Bacteroidetes, Proteobacteria, and Firmicutes in response to changes in brain function resulting from TBI.
The bidirectional communication of the gut-brain axis is being researched as a potential target for the treatment of neuronal injury. For example, preclinical data indicate that probiotic supplementation is neuroprotective following traumatic neural injury and cerebral ischemia/reperfusion injury [61, 62]. Zhang and Jiang report that probiotics act to strengthen the intestinal barrier, inhibiting the systemic dysregulation of the immune system that occurs after Neurotrauma [61]. Other researchers have found that modulating the gut microbiota of mice with probiotics reduces systemic inflammation and suppresses microglial hyperactivation after induction of a gut inflammatory response [63]. Similarly, Docosahexaenoic acid (DHA) supplementation has been found to have strong anti- inflammatory and neuroprotective effects, reducing axonal damage in an experimental murine model of TBI [64]. These preclinical studies all align with the hypothesis of treating the gut to heal the brain. Regarding neurotrauma, it is believed that neurodegenerative pathology starts in the gut before being observed in the CNS via the vagus nerve [3]. In preclinical models, vagus nerve stimulation (VNS) was also found to have neuroprotective effects when administered both prophylactically and as a treatment following TBI [65]. Although the exact mechanisms have not been explicated, VNS reduced the breakdown of the blood-brain barrier, attenuated cerebral inflammation and edema, and improved cognitive and motor recovery in comparison to control animals [65].
There has been much preclinical success with microbiome and neurotrauma research, but most of these findings have not resulted in changes to clinical management of Neurotrauma [3]. From a translational perspective, there is a challenge in the fact that overcoming the differences in the gut microflora of experimental animals and humans [66]. It is hopeful that as more work is done to understand the mechanisms underlying many of these preclinical findings, issues with differences in gut microflora will diminish.
Proteinopathy
As previously described, several different insults to the GI system can lead to aberrations in the gut-brain axis resulting in a cyclic process of chronic neuroinflammation [67, 68, 69]. This chronic process can subsequently result in neurodegeneration, often secondary to proteinopathy, as evidenced in many neurodegenerative diseases such as Alzheimer’s disease (AD), Parkinson’s disease (PD), and Amyotrophic lateral sclerosis (ALS) [70, 71]. Proteinopathy, described as the misfolding and aggregation of various proteins and enzymes within the CNS and peripheral nervous system, leads to cycles of inflammation and maladaptive immune responses that eventually lead to neuronal cell dysfunction and death [72]. There currently exists evidence in literature that highlights the role of gut dysbiosis in a defective BBB, changes in inflammatory mediators and immune cells, increased oxidative stress, and defective autophagy, the culmination of which can result in neurotoxic proteinopathy (Figure 5).
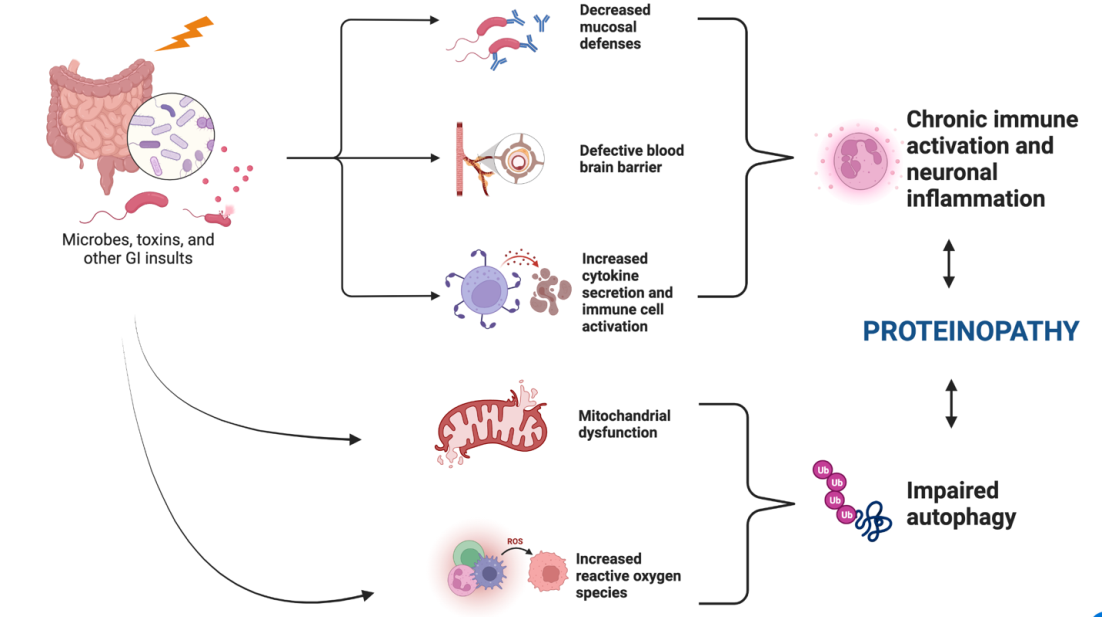
Several studies have shown the association of gut dysbiosis with a defective BBB and diminished mucosal defenses [73]. This encourages the translocation of gut organisms and their associated neurotoxic metabolites to alter immune signaling pathways in the CNS. Immune signaling is altered by these metabolites as they act as pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs), activating pattern recognition receptors of the innate immune system [74]. These immune signaling changes are mediated by functional changes to dendritic cells, glial cells, B and T cells, among other cells in the PNS [75]. The resulting inflammatory response changes lead to chronic activation of proinflammatory cytokines and can be exacerbated by cortisol secretion secondary to CNS stress from a defective BBB [76]. In addition, gut dysbiosis influences changes in the composition of gut microbe metabolites such as SCFAs, which alter gene expression in the CNS and regulate oxidative stress at the BBB [77, 78]. A study conducted by Brenner et al. demonstrated that an abundance of cyanobacteria in the gut microenvironment is associated with depletion of glutathione, a potent cellular antioxidant [79]. Further, Mecocci, et al. previously found increased oxidative damage to mitochondrial DNA in AD brain tissue samples [80]. Wang, et al. also showed that excessive production of reactive oxygen species (ROS) led to endoplasmic reticulum (ER) stress and resulting damage to cellular macromolecules. This resulted in accumulation of misfolded proteins via aberrations in ER homeostasis [81].
The increase in misfolded proteins and damaged macromolecules overwhelms an already dysfunctional autophagy system. Autophagy represents a cellular self- degradative process that maintains a balance of stored cellular debris and other macromolecules. In states of chronic inflammation and oxidative stress, proinflammatory molecules stimulate microglia and astrocytes, further disrupting the immune response and impairing autophagy [82]. Impaired autophagy has been linked to the development of neurodegenerative diseases such as AD [83]. In fact, Yang, et al. showed that in a mouse model for AD, deletion of cystatin B, an inhibitor of lysosomal cysteine proteases, led to a reversal in autophagy pathology [84]. This reversal was associated with decreased accumulation of amyloid- beta peptides among other substrates and rescue of memory impairment.
Management Considerations for Clinicians
Our findings show that there is substantial evidence in the literature implicating the gut microbiota and the gut brain axis in physiologic and behavioral changes. It is therefore imperative that psychologists, physiologists, and other clinicians who are caring for patients recovering from significant neurotrauma and/or other bodily insults keep in mind the importance of the microorganismal composition of the gut and its effect on the gut-brain-axis. For instance, pain is often an important factor in patient recovery and is common following trauma. Luczynski, et al. demonstrated in an in vivo model that germ free mice displayed higher response to visceral pain [85]. In addition, increased expression of toll-like receptors and pro-inflammatory cytokines were observed from the spinal cord of germ-free mice. This is of particular importance as changes in the gut microbiome can influence pain perception, leading to delayed or improved recovery. Furthermore, aberrations in gut-brain axis has negative implications for the ability to sustain emotional stress, temperament, dietary behavior, and social and reproductive behavior [86]. These can influence a patient’s motivation to pursue physical therapy, following up with clinical appointments, and maintaining a positive outlook on their potential for recovery. Clinicians should keep in mind that all these factors influence the rate and overall quality of recovery from a physical and mental capacity. We argue that a greater emphasis should be placed on patient education of proper dietary and lifestyle habits to improve gut microbiota in hopes of alleviating suffering from neurological or mental disorders. Moreover, we recommend clinicians maintain a low threshold for referral to psychological or psychiatric services to aid in management of this patient population for more holistic care.
Conclusion
Recent research has elucidated several mechanisms by which neurotrauma may induce alterations in the gut-brain axis, including the incitement of autonomic dysfunction. Dysfunction in the gut-brain axis can, in turn, lead to the development of secondary neurologic injury through enhanced neuroinflammation and the development of proteinopathy. Much of this data has come from preclinical studies; known differences in gut microflora between animals and humans necessitates further exploration into the mechanisms of gut-brain axis dysregulation in human neurotrauma patients. Nonetheless, innovations such as FMT are beginning to show promise in reducing gut dysbiosis seen in neurotrauma patients, thereby reducing the potential for secondary neurologic injury.
References
-
Dinan TG, Cryan JF (2017) Brain-Gut-Microbiota Axis and Mental Health. Psychosom Med 79(8): 920-926.
-
Osadchiy V, Martin CR, Mayer EA (2019) The Gut-Brain Axis and the Microbiome: Mechanisms and Clinical Implications. Clin Gastroenterol Hepatol 17(2): 322-332.
-
Sundman MH, Chen NK, Subbian V, Chou YH (2017) The bidirectional gut-brain-microbiota axis as a potential nexus between traumatic brain injury, inflammation, and disease. Brain Behav Immun 66: 31-44.
-
Margolis KG, Cryan JF, Mayer EA (2021) The Microbiota- Gut-Brain Axis: From Motility to Mood. Gastroenterology 160(5): 1486-1501.
-
Gomaa EZ (2020) Human gut microbiota/microbiome in health and diseases: a review. Antonie Van Leeuwenhoek 113(12): 2019-2040.
-
Quigley EMM (2017) Microbiota-Brain-Gut Axis and Neurodegenerative Diseases. Curr Neurol Neurosci Rep 17(12): 94.
-
Cryan JF, O’Riordan KJ, Cowan CSM, Sandhu KV, Bastiaanssen TFS, et al. (2019) The Microbiota-Gut- Brain Axis. Physiol Rev 99(4): 1877-2013.
-
Blanke EN, Holmes GM, Besecker EM (2021) Altered physiology of gastrointestinal vagal afferents following neurotrauma. Neural Regen Res 16(2): 254-263.
-
Opeyemi OM, Rogers MB, Firek BA, Feldman KJ, Vagni V, et al. (2021) Sustained Dysbiosis and Decreased Fecal Short-Chain Fatty Acids after Traumatic Brain Injury and Impact on Neurologic Outcome. J Neurotrauma 38(18): 2610-2621.
-
Urban RJ, Pyles RB, Stewart CJ, Ajami N, Randolph KM, et al. (2020) Altered Fecal Microbiome Years after Traumatic Brain Injury. J Neurotrauma 37(8): 1037- 1051.
-
Hanscom M, Loane DJ, Donohue TS (2021) Brain-gut axis dysfunction in the pathogenesis of traumatic brain injury. J Clin Invest 131(12): e143777.
-
Lemke DM (2004) Riding out the storm: sympathetic storming after traumatic brain injury. J Neurosci Nurs 36(1): 4-9.
-
Khalid F, Yang GL, McGuire JL, Robson MJ, Foreman B, et al. (2019) Autonomic dysfunction following traumatic brain injury: translational insights. Neurosurg Focus 47(5): E8.
-
Patterson TT, Nicholson S, Wallace D, Hawryluk GWJ, Grandhi R (2019) Complex Feed-Forward and Feedback Mechanisms Underlie the Relationship Between Traumatic Brain Injury and the Gut-Microbiota-Brain Axis. Shock 52(3): 318-325.
-
Uesaka T, Young HM, Pachnis V, Enomoto H (2016) Development of the intrinsic and extrinsic innervation of the gut. Dev Biol 417(2): 158-167.
-
Mittal R, Debs LH, Patel AP, Nguyen D, Patel K, et al. (2017) Neurotransmitters: The Critical Modulators Regulating Gut-Brain Axis. J Cell Physiol 232(9): 2359-2372.
-
Shin SS, Dixon CE (2015) Alterations in Cholinergic Pathways and Therapeutic Strategies Targeting Cholinergic System after Traumatic Brain Injury. J Neurotrauma 32(19): 1429-1440.
-
Griffin SL, Reekum RV, Masanic C (2003) A review of cholinergic agents in the treatment of neurobehavioral deficits following traumatic brain injury. J Neuropsychiatry Clin Neurosci 15(1): 17-26.
-
Costantini TW, Bansal V, Peterson CY, Loomis WH, Putnam JG, et al. (2010) Efferent vagal nerve stimulation attenuates gut barrier injury after burn: modulation of intestinal occludin expression. J Trauma 68(6): 1349- 1354.
-
Browning KN, Travagli RA (2014) Central nervous system control of gastrointestinal motility and secretion and modulation of gastrointestinal functions. Compr Physiol 4(4): 1339-1368.
-
Van Der Zanden EP, Boeckxstaens GE, de Jonge WJ (2009) The vagus nerve as a modulator of intestinal inflammation. Neurogastroenterol Motil 21(1): 6-17.
-
Cailotto C, Gomez-Pinilla PJ, Costes LM, van der Vliet J, Di Giovangiulio M, et al. (2014) Neuro-anatomical evidence indicating indirect modulation of macrophages by vagal efferents in the intestine but not in the spleen. PLoS One 9(1): e87785.
-
Toklu HZ, Sakarya Y, Tumer N (2017) A Proteomic Evaluation of Sympathetic Activity Biomarkers of the Hypothalamus-Pituitary-Adrenal Axis by Western Blotting Technique Following Experimental Traumatic Brain Injury. Methods Mol Biol 1598: 313-325.
-
Yoo BB, Mazmanian SK (2017)The Enteric Network: Interactions between the Immune and Nervous Systems of the Gut. Immunity 46(6): 910-926.
-
Song J, Wang T, Zhang X, Li B, Zhu C, et al. (2020) Upregulation of gastric norepinephrine with beta- adrenoceptors and gastric dysmotility in a rat model of functional dyspepsia. Physiol Res 69(1): 135-143.
-
Mailman D (1982) Blood flow an intestinal absorption. Fed Proc 41(6): 2096-2100.
-
Kharrazian D (2015) Traumatic Brain Injury and the Effect on the Brain-Gut Axis. Altern Ther Health Med 21(Suppl 3): 28-32.
-
Kigerl KA, Hall JC, Wang L, Mo X, Yu Z, et al. (2016) Gut dysbiosis impairs recovery after spinal cord injury. Journal of Experimental Medicin 213(12): 2603-2620.
-
Benakis C, Brea D, Caballero S, Faraco G, Moore J, et al. (2016) Commensal microbiota affects ischemic stroke outcome by regulating intestinal γδ T cells. Nature medicine 22(5): 516-523.
-
Round JL, Mazmanian SK (2009) The gut microbiota shapes intestinal immune responses during health and disease. Nature reviews immunology 9(5): 313-323.
-
31. Borody TJ, Khoruts A (2012) Fecal microbiota transplantation and emerging applications. Nature reviews Gastroenterology & hepatology 9(2): 88-96.
-
Aroniadis OC, Brandt LJ (2013) Fecal microbiota transplantation: past, present and future. Current opinion in gastroenterology 29(1): 79-84.
-
Singh V, Roth S, Llovera G, Sadler R, Garzetti D, et al. (2016) Microbiota dysbiosis controls the neuroinflammatory response after stroke. Journal of Neuroscience 36(28): 7428-7440.
-
Erny D, Hrabe de Angelis A, Jaitin D, Wieghofer P, Staszewski O, et al. (2015) Host microbiota constantly control maturation and function of microglia in the CNS. Nature Neuroscience 18: 965-977.
-
Di Giacinto C, Marinaro M, Sanchez M, Strober W, Boirivant M (2005) Probiotics ameliorate recurrent Th1- mediated murine colitis by inducing IL-10 and IL-10- dependent TGF-β-bearing regulatory cells. The Journal of Immunology 174(6): 3237-3246.
-
Han B, Sivaramakrishnan P, Lin C-CJ, Neve IA, He J, et al. (2017) Microbial genetic composition tunes host longevity. Cell 169(7): 1249-1262.
-
Kasubuchi M, Hasegawa S, Hiramatsu T, Ichimura A, Kimura I (2015) Dietary gut microbial metabolites, short-chain fatty acids, and host metabolic regulation. Nutrients 7(4): 2839-2849.
-
Stilling RM, van de Wouw M, Clarke G, Stanton C, Dinan TG, et al. (2016) The neuropharmacology of butyrate: the bread and butter of the microbiota-gut-brain axis? Neurochemistry international 99: 110-132.
-
Lu J, Frerich JM, Turtzo LC, Li S, Chiang J, et al. (2013) Histone deacetylase inhibitors are neuroprotective and preserve NGF-mediated cell survival following traumatic brain injury. Proceedings of the National Academy of Sciences 110(26): 10747-10752.
-
Li H, Sun J, Du J, Wang F, Fang R, et al. (2018) Clostridium butyricum exerts a neuroprotective effect in a mouse model of traumatic brain injury via the gut‐brain axis. Neurogastroenterology & Motility 30(5): e13260.
-
D’Mello C, Ronaghan N, Zaheer R, Dicay M, Le T, et al. (2015) Probiotics improve inflammation-associated sickness behavior by altering communication between the peripheral immune system and the brain. Journal of Neuroscience 35(30): 10821-10830.
-
Sun G, Yang S, Cao G, Wang Q, Hao J, et al. (2018) γδ T cells provide the early source of IFN-γ to aggravate lesions in spinal cord injury. J Exp Med 215(2): 521-535.
-
McKinlay A, Lin A, Than M (2018) A comparison of emergency department medical records to parental self-reporting of traumatic brain injury symptoms. Concussion 3(1): CNC52.
-
Pozzato I, Meares S, Kifley A, Craig A, Gillett M, et al. (2020) Challenges in the acute identification of mild traumatic brain injuries: results from an emergency department surveillance study. BMJ Open 10(2): e034494.
-
Chen AJ, D’Esposito M (2010) Traumatic brain injury: from bench to bedside to society. Neuron 66(1): 11-14.
-
Furger RE, Nelson LD, Brooke Lerner E, McCrea MA (2016) Frequency of Factors that Complicate the Identification of Mild Traumatic Brain Injury in Level I Trauma Center Patients. Concussion 1(2): CNC11.
-
Risling M, Davidsson J (2012) Experimental animal models for studies on the mechanisms of blast-induced neurotrauma. Front Neurol 3: 30.
-
Simon DW, Rogers MB, Gao Y, Vincent G, Firek BA, et al. (2020) Depletion of gut microbiota is associated with improved neurologic outcome following traumatic brain injury. Brain Res 1747: 147056.
-
O’Connor G, Jeffrey E, Madorma D, Marcillo A, Abreu MT, et al. (2018) Investigation of Microbiota Alterations and Intestinal Inflammation Post-Spinal Cord Injury in Rat Model. J Neurotrauma 35(18): 2159-2166.
-
Dabrowski W, Siwicka-Gieroba D, Kotfis K, Zaid S, Terpilowska S, et al. (2021) The Brain-gut Axis-where are we now and how can we Modulate these Connections? Curr Neuropharmacol 19(8): 1164-1177.
-
Mayer EA, Tillisch K, Gupta A (2015) Gut/brain axis and the microbiota. J Clin Invest 125(3): 926-938.
-
Bercik P, Denou E, Collins J, Jackson W, Lu J, et al. (2011) The intestinal microbiota affect central levels of brain- derived neurotropic factor and behavior in mice. Gastroenterology 141(2): 599-609, 609.e1-3.
-
Collins SM, Kassam Z, Bercik P (2013) The adoptive transfer of behavioral phenotype via the intestinal microbiota: experimental evidence and clinical implications. Curr Opin Microbiol 16(3): 240-245.
-
Sudo N, Chida Y, Aiba Y, Sonoda J, Oyama N, et al. (2004) Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. J Physiol 558(Pt 1): 263-275.
-
Dinan TG, Cryan JF (2012) Regulation of the stress response by the gut microbiota: implications for psychoneuroendocrinology. Psychoneuroendocrinology 37(9): 1369-1378.
-
Bercik P, Collins SM, Verdu EF (2012) Microbes and the gut-brain axis. Neurogastroenterol Motil 24(5): 405-413.
-
Celorrio M, Abellanas MA, Rhodes J, Goodwin V, Moritz J, et al. (2021) Gut microbial dysbiosis after traumatic brain injury modulates the immune response and impairs neurogenesis. Acta Neuropathol Commun 9(1): 40.
-
Treangen TJ, Wagner J, Burns MP, Villapol S (2018) Traumatic Brain Injury in Mice Induces Acute Bacterial Dysbiosis Within the Fecal Microbiome. Front Immunol 9: 2757.
-
You W, Zhu Y, Wei A, Du J, Wang Y, et al. (2022) Traumatic Brain Injury Induces Gastrointestinal Dysfunction and Dysbiosis of Gut Microbiota Accompanied by Alterations of Bile Acid Profile. J Neurotrauma 39(1-2): 227-237.
-
Houlden A, Goldrick M, Brough D, Vizi ES, Lénárt N, et al. (2016) Brain injury induces specific changes in the caecal microbiota of mice via altered autonomic activity and mucoprotein production. Brain Behav Immun 57: 10-20.
-
Zhang X, Jiang X (2015) Effects of enteral nutrition on the barrier function of the intestinal mucosa and dopamine receptor expression in rats with traumatic brain injury. JPEN J Parenter Enteral Nutr 39(1): 114-123.
-
Sun J, Wang F, Ling Z, Yu X, Chen W, et al. (2016) Clostridium butyricum attenuates cerebral ischemia/ reperfusion injury in diabetic mice via modulation of gut microbiota. Brain Res 1642: 180-188.
-
D’Mello C, Ronaghan N, Zaheer R, Dicay M, Le T, et al. (2015) Probiotics Improve Inflammation-Associated Sickness Behavior by Altering Communication between the Peripheral Immune System and the Brain. J Neurosci 35(30):10821-10830.
-
Mills JD, Bailes JE, Sedney CL, Hutchins H, Sears B (2011) Omega-3 fatty acid supplementation and reduction of traumatic axonal injury in a rodent head injury model. J Neurosurg 114(1): 77-84.
-
Neren D, Johnson MD, Legon W, Bachour SP, Ling G, et al. (2016) Vagus Nerve Stimulation and Other Neuromodulation Methods for Treatment of Traumatic Brain Injury. Neurocrit Care 24(2): 308-319.
-
Blaser M, Bork P, Fraser C, Knight R, Wang J (2013) The microbiome explored: recent insights and future challenges. Nat Rev Microbiol 11(3): 213-217.
-
Borre YE, O’Keeffe GW, Clarke G, Stanton C, Dinan TG, et al. (2014) Microbiota and neurodevelopmental windows: implications for brain disorders. Trends Mol Med 20(9): 509-518.
-
Luan H, Wang X, Cai Z (2019) Mass spectrometry-based metabolomics: Targeting the crosstalk between gut microbiota and brain in neurodegenerative disorders. Mass Spectrom Rev 38(1): 22-33.
-
Chidambaram SB, Essa MM, Rathipriya AG, Bishir M, Ray B, et al. (2022) Gut dysbiosis, defective autophagy and altered immune responses in neurodegenerative diseases: Tales of a vicious cycle. Pharmacol Ther 231: 107988.
-
Liao YZ, Ma J, Dou JZ (2022) The Role of TDP-43 in Neurodegenerative Disease. Mol Neurobiol 59(7): 4223- 4241.
-
Hayes LR, Kalab P (2022) Emerging Therapies and Novel Targets for TDP-43 Proteinopathy in ALS/FTD. Neurotherapeutics.
-
Cryan JF, Dinan TG (2012) Mind-altering microorganisms: the impact of the gut microbiota on brain and behaviour. Nat Rev Neurosci 13(10): 701-712.
-
Galea I (2021) The blood-brain barrier in systemic infection and inflammation. Cell Mol Immunol 18(11): 2489-2501.
-
de Vos WM, de Vos EA (2012) Role of the intestinal microbiome in health and disease: from correlation to causation. Nutr Rev 70(1): S45-S56.
-
Subbarayan MS, Hudson C, Moss LD, Nash KR, Bickford PC (2020) T cell infiltration and upregulation of MHCII in microglia leads to accelerated neuronal loss in an α-synuclein rat model of Parkinson’s disease. J Neuroinflammation 17(1): 242.
-
Hoban DB, Connaughton E, Connaughton C, Hogan G, Thornton C, et al. (2013) Further characterisation of the LPS model of Parkinson’s disease: a comparison of intra-nigral and intra-striatal lipopolysaccharide administration on motor function, microgliosis and nigrostriatal neurodegeneration in the rat. Brain Behav Immun 27(1): 91-100.
-
Cenit MC, Sanz Y, Codoñer-Franch P (2017) Influence of gut microbiota on neuropsychiatric disorders. World J Gastroenterol 23(30): 5486-5498.
-
Dalile B, Van Oudenhove L, Vervliet B, Verbeke K (2019) The role of short-chain fatty acids in microbiota-gut- brain communication. Nat Rev Gastroenterol Hepatol 16(8): 461-478.
-
Brenner SR (2013) Blue-green algae or cyanobacteria in the intestinal micro-flora may produce neurotoxins such as Beta-N-Methylamino-L-Alanine (BMAA) which may be related to development of amyotrophic lateral sclerosis, Alzheimer’s disease and Parkinson-Dementia- Complex in humans and Equine Motor Neuron Disease in horses. Med Hypotheses 80(1): 103.
-
Mecocci P, MacGarvey U, Beal MF (1994) Oxidative damage to mitochondrial DNA is increased in Alzheimer’s disease. Ann Neurol 36(5): 747-751.
-
Senft D, Ronai ZA (2015) UPR, autophagy, and mitochondria crosstalk underlies the ER stress response. Trends Biochem Sci 40(3): 141-148.
-
Schwartz M, Baruch K (2014) The resolution of neuroinflammation in neurodegeneration: leukocyte recruitment via the choroid plexus. Embo J 33(1): 7-22.
-
Wilson CA, Murphy DD, Giasson BI, Zhang B, Trojanowski JQ, et al. (2004) Degradative organelles containing mislocalized alpha-and beta-synuclein proliferate in presenilin-1 null neurons. J Cell Biol 165(3): 335-346.
-
Yang DS, Stavrides P, Mohan PS, Kaushik S, Kumar A, et al. (2011) Reversal of autophagy dysfunction in the TgCRND8 mouse model of Alzheimer’s disease ameliorates amyloid pathologies and memory deficits. Brain 134(1): 258-277.
-
Luczynski P, Tramullas M, Viola M, Shanahan F, Clarke G, et al. (2017) Microbiota regulates visceral pain in the mouse. Elife 6: e25887.
-
Liang S, Wu X, Jin F (2018) Gut-Brain Psychology: Rethinking Psychology from the Microbiota-Gut-Brain Axis. Front Integr Neurosci 12: 33.
- A Review of Gene Therapy for Parkinson's Disease to Control Dopaminergic Neurons
- Late-Onset Myasthenia Gravis in a Patient with Recurrent Breast Cancer: A Case Report
- Covid-Induced Dystonia and Opsoclonus: A Case Report
- Generalized Tonic-Clonic Seizure in a Pediatric Patient with Sunflower Syndrome: A Case Report
- Comparison of Doppler Guided Seldinger Technique Versus Classic Palpatory Seldinger Technique for Radial Artery Cannulation-an Open Label Randomized Controlled Trial
- Brown Sequard Syndrome: Understanding the Complexities of Spinal Cord Injury