Chronically Elevated Triglycerides as a Result of High Fat Palatable Diet Resulting in a Vicious Cycle on Reinforcing Reward and Dopamine Signaling: A Possible Cause for the Obesity Epidemic Worldwide in the Food Environment Available-A Comprehensive Review
Obesity is assuming epidemic proportions. We don’t have any medical therapy that can be used for long term. Although bariatric Surgery (BS) results in immediate weight loss neither it can be afforded by all individuals nor is it safe with the comorbidities of obesity. Hence it has become essential to understand the etiopathology of increasing obesity by leaps and bounds to get a long-term sustainable cure. Volkow, et al. gave a comparison of obesity, simulating that of patients of drugs of abuse, where compulsive intake of tasty food, that is high in lipids and sugars, much >the metabolic requirements. Although hypothalamus remains the basic area where integration of circulating signals like leptin, ghrelin and nutrients occurs from the periphery further it is influenced by both higher centers along with sensory influences like tastes and smell along with emotional states. Earlier we had reviewed how different higher centers influence hypothalamus for final intake of food, here we discuss in detail the role of mesolimbic pathways as far as the rewarding part of feeding is concerned. Role of dopamine signaling, its receptors and genes controlling are further discussed in detail.
Introduction
Incidences of obesity and consequent metabolic complications are getting out of hand. Not only is it involving adults but children as well. It has become very important to understand the etiopathogenesis of the same with the increase getting out of proportion and no clear medical therapy in sight. We have been trying to search for the etiopathogenesis of obesity for over a decade [1, 2, 3, 4, 5, 6]. Since the concept of Food Addiction or brain circuitry in obese responds in a manner similar to that of drug addicts by Virchow’s group we have been trying to follow that hypothesis and further emphasized how it is important to study other brain areas involved in cognition food reward emotion separately regarding obesity with different effects that fronto striatal circuits may override the hypothalamic circuits [7]. Here we further try to emphasize on the role of increased triglycerides affecting the mesolimbic circuits and its influence on various genetic factors like the Taq A1 allele carriers and other genes like ANKK1 and non-carriers and effect of other genes like the FTO gene.
Methods
We Carried out a review utilizing the Search Engine PubMed using the MeSH terms mesolimbic brain system, dopaminergic receptors; DRD1-5; Genes related to dopaminergic signaling; high fat diet (HFD) fatty acid (FA) sensing in brain; saturated fats sensing; role of lipoproteins (LP); ANKK1; FTO. As for inclusion and exclusion Criteria are all studies limited to food reward were initially downloaded and then further articles pertaining to fat and lipid metabolism in brain were used excluding those specifically for carbohydrate metabolism and any article whether favoring reward hypothesis or refuting was used. Since limited laboratories are working on this field we took out more articles from cross references and article search further from names of workers method shown in Figure 1.
Kulvinder Kochar Kaur, et al. Chronically Elevated Triglycerides as a Result of High Fat Palatable Diet Resulting in a Vicious Cycle on Reinforcing Reward and Dopamine Signaling: A Possible Cause for the Obesity Epidemic Worldwide in the Food Environment Available-A Comprehensive Review. J Endocrinol 2019, 3(2): 000143.
Copyright© Kulvinder Kochar Kaur, et al.
Results and Discussion
We found a total of 250 articles out of which we selected 135 articles for this review. No meta-analysis was done. Limitations of this study are as not many groups are working on this field namely how TG’s affect brain reward system especially ML system and DA receptor action including reward circuitry incentive salience and, working memory, and other cognitive parameters opinion of only 2-3 groups available on this subject.
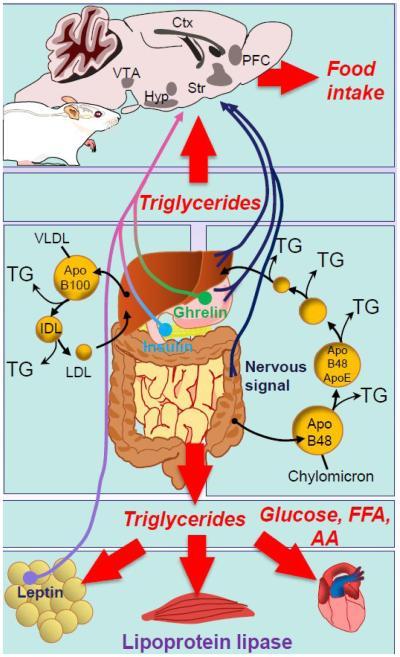
Proper energy homeostasis gets obtained, once energy intake along with needs reach an equilibrium at a definite metabolic set point. With marked evolutionary changes a system that responds markedly and coordinates a variety of signals related to hunger along with satiety via complicated along with needless interaction of neural circuits which had been basically meant for long time energy homeostasis. The hypothalamic-brain stem axis has been observed to be a key controller of energy balance within the central nervous system (CNS). In the circulation the signals that correspond to energy like leptin, ghrelin, insulin along with nutrients get identified by and change the action of specific neuronal areas which subsequently involve neuroendocrine, peripheral nervous and finally behavioral systems to get accustomed to nutrient input and energy demands (Figure 2).
Kulvinder Kochar Kaur, et al. Chronically Elevated Triglycerides as a Result of High Fat Palatable Diet Resulting in a Vicious Cycle on Reinforcing Reward and Dopamine Signaling: A Possible Cause for the Obesity Epidemic Worldwide in the Food Environment Available-A Comprehensive Review. J Endocrinol 2019, 3(2): 000143.
This complicated behavioral consequences which effect food intake depend on integrative processes from higher areas in CNS code for motivation, reward, habit, emotional behavior, along with memory which get influenced by nutritional status and several hypothalamic along with extra hypothalamic networks of brain. Crucial in these being mesolimbic (ML) circuits, where dopamine (DA) secretion has been demonstrated to code for the reinforcing along with motivating qualities of High fat (HF) along with high sugar (HFHS) foods [8, 9]. Especially the projections of DA neurons from the ventral tegmental area (VTA) in the midbrain to the nucleus acumbens (NAc) is a basic neural substrate for the action of drugs of abuse, hence the ML is known as the ‘’brain reward circuit’’. It is clear that energy associated signals like leptin, ghrelin, insulin also target ML [10].
ref no. 65-Central integration of peripheral nervous and hormonal inputs that regulate energy balance. Gut- derived nervous and circulating factors convey a satiety signal and include vagal inputs from stomach or digestive tracts as well as secreted peptides such as cholecystokinin (CCK), PYY3-36, or glucagon-like peptide 1 (GLP-1). Ghrelin is secreted primarily by the stomach and positively regulates feeding while insulin or leptin act as long-term satiety factors. Ghrelin, leptin, and insulin have targets in the hypothalamus as well as the reward circuitry. Dietary lipids are esterified into triglycerides (TG) and packaged in nascent chylomicron (CM) at the level of the gut, secreted first in to the lymphatic system, and then the bloodstream. TG-rich CM gradually loses their lipid content upon action of tissue lipoprotein lipase (LPL) and ultimately recaptured as remnant CM by the liver. In the process CM exchange their native Apolipoprotein B48 (ApoB48) component for the apolipoprotein E (ApoE). Very-low density lipoprotein (VLDL), produced by the liver, represent another source of TG-rich particles which, upon action of LPL, give rise to intermediate-density (IDL) and low-density (LDL) lipoproteins. LPL is also expressed in the brain in both hypothalamic structures and ML structures including the prefrontal cortex (PFC), the hippocampus, ventral tegmental area, and throughout dorsal and ventral striatum. TG hydrolysis in the NAc regulates the rewarding and motivational aspects of food intake and could be an important mechanism linking dietary input with reward.
HFD, Obesity and DA Signaling
Hypothalamic areas are present near the circum ventricular organs (CVO) and are considered to be the Copyright© Kulvinder Kochar Kaur, et al.
primary neural areas affected by nutrient excess [11]. Molecular explanation s concerning increased nutrient consumption and changes in neural function involves FFA metabolism in the hypothalamus [12], nutrient caused endoplasmic reticulum(ER) stress [13], inflammation [14] or resistance to energy associated signals [13, 15]. Further there is proper proof that directly DA signaling gets influenced by HFD in humans along with rodents. Like massive dopamine D2 receptor (D2R) present in the striatum is inversely associated with body weight [16], along with compulsive eating observed in obese rats, when measurement was done of tasty food consumption inspite of aversive conditioned stimulus [17]. Silencing the D2R in the dorsal striatum by genetic means, increases the formation of a reward-absent state along with compulsive eating in rats that have faced HFD. Moreover in rats individual differences in motivational effect of food-associated cues, further predicted how much gain in weight would be there along with their trying to work to obtain the food rewards. Greater craving correlated with a fast alteration in dorsal striatum DA signaling, although not with opioid signaling in the NAc [18].
Earlier studies demonstrated that in humans striatal D2R present in obese people were significantly less and had a negative association with body mass index (BMI) [16, 19]. Moreover blood oxygen level dependent (BOLD) signals in striatal areas, checked using functional magnetic resonance imaging (fMRI) was reduced in obese vis a vis lean patients, pointing to defective neuronal activity in the striatal neurons [20]. Conversely obesity was related to higher BOLD response related to food associated cues in brain areas that controlled reward and motivation. Underactive striatal neurons at baseline response in obese subjects, in contrast to them showing sensitized response to food signals are what above data points. For quantification of striatal D2R-like binding potential (D2BP), Guo, et al. [21], utilized positron emission tomography (PET) and found a correlation between striatal DA binding in obesity, that implicated DA signaling in this process. A negative association was observed of BMI with D2BP in the ventral striatum, in contrast to dorsal striatum, where both BMI along with either habit of eating or eating at any opportunity was positively associated with D2BP. It is still not clear whether changes in DA signaling causes obesity or occurs secondary to weight rise, with this research observing a complicated way of DA signaling abnormalities in obese people, in whom different alterations might generate a state of reward deficiency along with increased habitual response.
Kulvinder Kochar Kaur, et al. Chronically Elevated Triglycerides as a Result of High Fat Palatable Diet Resulting in a Vicious Cycle on Reinforcing Reward and Dopamine Signaling: A Possible Cause for the Obesity Epidemic Worldwide in the Food Environment Available-A Comprehensive Review. J Endocrinol 2019, 3(2): 000143.
Various studies emphasize that the presence of defects that are present within ML function form independent of increased body weight and despite getting exaggerated by obesity state, might be an initial outcome of exposure to dietary fat. Despite degree and what direction these alterations take differ as per the diet, strain or which area of brain has been used; HFD exposure can independently as far as body weight rise is concerned, lead to increased D2R increase, Increase DA turnover rate, function of the DA transporter (DAT), increase amphetamine response and response to food reward [22]. If animals were given calories that had been restricted calories through HFD but not high sugar diet showed reduced attention and increase impulsivity once checked by 5 choice serial reaction task (5CSRT) [23]. Further reduced exposure to a fat source also initiated binge like eating behavior along with sensitivity of ML activity, while mice that had absence of ghrelin receptor did not accelerate palatable food intake that indicated that energy associated signals like ghrelin participate in ML response to energy enriched food [24]. All fat sources equally interfered with ML activity. Exposure to saturate but not unsaturated fat caused alteration in D1R and DAT excess in the ML [25].
Emphasis on TG metabolism was paid, in the form of it being a big predictor of compulsive property. SD rats might be subdivided into obesity prone (OP) and resistant (OR) on the property of metabolic characteristics along with increase in body weight in a small 5-day HF challenge, in which changed TG that is circulating, fat partitioning that has enhanced LPL activity in the adipose tissue (AT), reduced muscle lipid transport were found as OP rats signatures [26]. Following a meal plasma TG predicted markedly, the coming body weight rise in OP rats, which is a higher TG rise following a meal (high TG responders) that was associated with excessive chances to eat extra [27]. Extracellular DA levels as investigated using micro dialysis in the NAc was decreased at both basal condition and in response to HFD or peripheral injection of a fat emulsion (intralipid) in OP high TG responders. Furthermore, on bypassing or sensory reward via systemic injection, TG emulsion and not glucose caused an increase in DA release in the NAc on measuring via micro dialysis [28]. Further impaired cognition in obese mice got better by giving pharmacologically lesser plasma TG, although centrally injecting TG reduced cognition in lean mice [29]. Sun, et al. utilized fMRI in humans showed an association between plasma TG along with ghrelin with the degree of the whole brain BOLD response to food reward. Larger postprandial decreases in ghrelin or rise in TG correlated with a decreased BOLD response to palatable milk shake Copyright© Kulvinder Kochar Kaur, et al.
in the limbic circuit having midbrain, palladium, amygdale, medial orbito frontal cortex (OFC) and hippocampus [30]. Circulating albumin bound FFA, glucose, or insulin didn’t correlate with brain responses to food reward.
Role of TG or FFA
Main constituents of brain are lipids that get formed from both endogenous syntheses along with dietary intake [31, 32]. Plasma lipids can be found as FFA that is bound to albumin and TG rich lipoproteins [33]. Lipoproteins represent a complex interrelation between apolipoproteins and phospholipids, which create a polar environment for lipid passage. Lipoprotein Lipase (LPL) that are expressed in peripheral tissues influences hydrolysis of TG from TG-rich particles like very low density Lipoproteins (VLDL), and gut brought chylomicrons (CM) to produce particles having decreased lipid amount like high density Lipoproteins (HDL).
What is the method by which lipids get access into brain? Utilizing positron emission tomography (PET) tracing research in association with radiolabelling of FA’s that are injected peripherally demonstrated dynamic incorporation of arachidonic acid or palmitic acids in the brain [32, 34]. Metabolic Syndrome (MetS) was found to be correlated with enhanced whole brain FFA uptake and particularly an 88% rise in hypothalamic FFA uptake [35]. Of the FFA, essential FFAs, poly unsaturated FA (PUFA) can cross the blood brain barrier (BBB) via modes poorly understood till recent finding of Mfsd2a (major facilitator super family domain containing 2a) as being the carrier for the absorption of the essential FA’s docosahexaenoic acid (DHA) in the brain [36]. Earlier free DHA was thought to be the main source of brain DHA, this study demonstrated that DHA with long chain fatty acids (LCFA) get transported via Mfsd2a as lysophosphatidyl choline (LPC) but not FFA [37]. This emphasized the complicated and redundant methods used for brain LCFA and DHA homeostasis.
Circulating apolipoproteins vary from 10nm for HDL, 20-30nm for LDL, 30-40nm for IDL, and 5-80nmfor VLDL and CM. Some small HDL may cross the BBB is known but how larger TG rich particles like VLDL or CM has been questioned on earlier tracer studies and with the initial presumption that the brain does not have a lymphatic system [33, 38]. Following food intake lipids get packed in CM, and get secreted via lymphatic system and then into the main circulation, from where these large particles need to cross BBB at the point of fenestrated capillaries to Kulvinder Kochar Kaur, et al. Chronically Elevated Triglycerides as a Result of High Fat Palatable Diet Resulting in a Vicious Cycle on Reinforcing Reward and Dopamine Signaling: A Possible Cause for the Obesity Epidemic Worldwide in the Food Environment Available-A Comprehensive Review. J Endocrinol 2019, 3(2): 000143.
get into the CNS. But recent finding of a lymphatic system in the mouse brain indicates the presence of a new passageway through which TG rich particles might be there for CSF interchange [39].
High levels of various LP receptors like VLDLR, LDLR, oxidized HDL receptors, and accessory proteins like LDL- receptor-related protein (LRP). Selective binding of these apolipoprotein compounds along with genetic along with pharmacological methods have emphasized the role of apolipoprotein signaling in the development of brain and function involving learning as well as memory, in addition to synaptic plasticity [40, 41]. Like apolipoprotein E that is a component of circulating CM and IDL, binds to LDLR is considered a main risk factor of the usual forms of Alzheimer’s disease (AD), although brain specific overexpression of LDLR increases β-apolipoproteins, basically that are produced by astrocytes, and particles roughly the size of HDL can get measured in cerebrospinal fluid (CSF) [42]. Thus demonstrating that both endogenous (astrocyte synthesized) along with peripherally formed (postprandial) LP particles affect brain functioning. Exact mode of action by which LP signaling gets started in brain is not understood and probably might involve canonical pathways, local lipid delivery and/or changes in cell metabolism that might then modulate neuronal action.
TG-lipases that hydrolyse TG into FFA s and mono or diacyl glycerol are expressed in huge numbers in the brain and especially abundant in ML [33, 43]. Brain lipases act on TG for mediating lipid deposition within specific brain nuclei that have essential functional results [44, 45]. LPL is the best explored of these as far as energy balance is concerned. Changed brain FFA and PUFA levels was seen in pan neuronal LPL knockout animals (NFXLPL-/-), who had late onset obesity developing, besides demonstrating age-associated reduction in cognitive function and rise in anxiety. Role of LPL in substructures of brain have been studied with > precision utilizing more restricted gain or loss of function approaches. Like hippocampal LPL controls energy expenditure and autonomic tone via manufacture on the ceramide based signaling pathway [46, 47].
Role of lipases at the time of brain development could probably explain the deficits seen in LPL knockout models; continued expression in isolated nuclei in adulthood can stimulate one to posit that brain TG lipases control local TG breakdown and LCFA availability. Although both TG and LCFA are circulating lipid species they appear in separate time points in the blood stream Copyright© Kulvinder Kochar Kaur, et al.
drastically opposite with regard to food intake. After meal intake there is collection of TG rich particles, while LCFA are liberated by fasting stimulated lipolysis and thus get enhanced at the time of absence of food [48]. Additionally these LCFA s can get transferred via FA transporters easily that are present in great numbers in most brain structures, TG need initial breakdown through TG-lipases that markedly prevents its central presence. Hence brain structures that possess TG-lipases might have rare quality of finding post prandial alterations in dietary lipids. The actual presence of LPL in ML indicates that there is an importance of TG sensing in post prandial changes brought through TG differences in reward valuation. That way breakdown of TG in the ML along with downstream adaptive changes take place once FFA s get released might have a direct effect on DA or other ML signaling pathways for controlling rewards looking behavior.
TG Sensing in Reward ML System
Berland, et al. developed a model where TG emulsion (intralipid TM) got perfused via the carotid artery in the direction of the brain at a concentration along with rate that simulates the post prandial increase in TG, without affecting the systemic lipids. With this model they revealed that TG can act directly in the brain for controlling locomotors activity, food preference, along with food seeking behaviors. Brain TG delivery reduced operant responding for rewards on a continuing ratio schedule, with preference for a tasty HFHS food in a food choice paradigm. Direct brain TG deposition reduced by approximately 50% night locomotor action and amphetamine-induced locomotion. Further TG also opposed D2R agonist-induced locomotion, which suggested a TG evoked control of the DA activity. Knockdown of LPL in NAc selectively had the reverse observations, causing increased motivation to food rewards working along with intake of palatable food.
Normally transiently there is an increase in TG following a meal. But plasma TG remains chronically increased in obesity and is not related with reduced tropism for calorie high foods, that points that adoptive mechanisms take place. To simulate the response of the brain to chronic hypertriglyceridemia they utilized sustained TG perfusion to lean animals comparing it with a model of diet induced obesity (DIO). They modeled hypertriglyceridemia with the use of a model of DIO or with chronic (7days) TG perfusion in the direction of the brain which enhanced brain TG sensing without any effect on plasma triglycerides levels. Both the treatments caused behavior specific desensitization, where central TG Kulvinder Kochar Kaur, et al. Chronically Elevated Triglycerides as a Result of High Fat Palatable Diet Resulting in a Vicious Cycle on Reinforcing Reward and Dopamine Signaling: A Possible Cause for the Obesity Epidemic Worldwide in the Food Environment Available-A Comprehensive Review. J Endocrinol 2019, 3(2): 000143.
sensing was not able to modulate tropism for tasty foods anymore but even now reduced locomotor action. This adaptive action stimulated by chronic increases in circulating TG or brain TG sensing, might explain how continuous intake of HF foods overwhelm the controlling systems to control weight gain. Central TG sensing can directly act in acute reduction in locomotor action which precedes metabolic alterations when animals get a western diet given to them [49]. On acute brain TG sensing it might help to decrease the desire for food reward. But if chronically increased TG sensing modes desensitize or result in compensatory adaptations so that decrease in physical action remain, although motivation for HFD becomes resistant to TG-mediated homeostatic control. Combination of both decreased physical activity and maintained motivation for HF foods will automatically equal to body weight gain.
Thus local TG breakdown in brain structures enriched with TG processing enzymes may have separate impacts as shown by above results. Albumin-bound LCFA’s in the circulation might basically act in the hypothalamus and act to control feeding and glucose production particularly in the period when food is scarce and thus adipose lipolysis causes FFA release in high amounts, as compared to LPL mediated hydrolysis of TG particles collected following a meal in ML structures may take part in the coding for incentive as well as motivation characteristics of food. While acute TG exposure, generally on meal intake, reduces both rewarding and motivational action of food, although chronic exposure causes desensitization, with unregulated excessive feeding behavior.
Molecular Aspect of ML Lipid Sensing
The presence of lipid sensing in the brain was introduced by Oomura, et al. [50] and has been studied in detail in the past 10yrs. LCFA metabolism controls neuronal activity, autonomic control of insulin release, food intake and liver glucose production [12, 15, 51]. Sensing of LCFA in hypothalamus covers variety of cellular modes of action that are direct entry into the tricarboxylic acid cycle (TCA cycle), amino acid controlled activation of m TOR [52], autophagy, inflammation via nuclear enhancer of kappa-light-chain-enhancer of activated B cells (IKK/NFκB) dependent pathways [13, 53], enhanced mitochondrial lipid β-oxidation, mitochondrial respiration getting adapted along with reactive oxygen species (ROS) scavenging [54], collection of lipid-mediated metabolites like acetyl CoA along with malonyl CoA, direct control of protein kinase C action [55], activation of membrane receptors via lipid Copyright© Kulvinder Kochar Kaur, et al.
mediation, eicosanoids controlled signaling as well as transcription adaptations that get activated by lipids [56].
Though high calories food is universally present, uncontrolled food intake and obesity does not involve every person that implicates that the current food environment may interact directly with both genetic along with epigenetic elements of susceptibility. Taq1A allele is a very good candidate regarding this. This affects 30-40% of population, homozygous dose of the A1 allele directly associates with a 30-40% decrease of striatal D2R excess [57] which has marked association with addiction along with compulsive behavior that has an effect on both drugs of abuse, along with feeding. The A1 allele occurs as a result of single nucleotide polymorphisms (SNP) that is present at the gene which codes for Ankyrin repeat and kinase domain containing 1 (ANKK1) near the gene encoding D2R [58]. ANKK1 is a receptor interacting protein (RIP) kinase; which is a structurally associated family of factors which integrate different stimuli that are inflammation innate immune response that is downstream of tumor necrosis factor alpha (TNFα-R1) receptor and Toll like receptor (TLR) and converge on cjun-N-terminal kinase (JNK), MAPK activity or NFκB signaling pathways [59].
Human protein-protein interaction studied with the use of in silico analysis showed that of the ~30 partners of human ANKK1 that have been anticipated, 50% are present in the NFkB, JNK, or MAPK pathway [60]. In brain ANKK1 is exclusively expressed in astrocytes. These astrocytes integrate different metabolic signals for coordinating neuronal activity and get targeted by lipids directly; particularly saturated fat induced inflammatory responses and ER stress brought about via TLR and IKK/NFκB signaling [61]. The way ANKK1 of astrocyte finally correlated with decreased D2R excess still remains a query, again FA metabolism, may give various probably modes. Like the D2R and ANKK1 promoters have NFκB cis regulatory elements [58, 62]; and lipid induced prostaglandins that are strong neural growth factor (NGF) stimulators, secreted by astrocytes. Further NGF directly controls D2R mRNA transcription in return via the NFκB signaling pathway [63].
Though excessive feeding secondary to an acute HFD exposure was demonstrated to involve astrocyte activation of NFκB, adaptations taking place on long term exposure of HFD may involve another partner, namely peroxisome proliferated activated receptor delta (PPARs). PPARδ is a member of a family of ligand activated transcription factors that control number of cellular Kulvinder Kochar Kaur, et al. Chronically Elevated Triglycerides as a Result of High Fat Palatable Diet Resulting in a Vicious Cycle on Reinforcing Reward and Dopamine Signaling: A Possible Cause for the Obesity Epidemic Worldwide in the Food Environment Available-A Comprehensive Review. J Endocrinol 2019, 3(2): 000143.
metabolic adaptations that initially respond to LCFA and prostaglandins. Further PPARδ has been found to be an essential controller of the ML. Like activation of PPARδ by LCFA or synthetic agonist reduce opioid synthesis or forebrain neurons along with proteins from methyl-4- phenyl-1,2,3,6-tetra hydropyridine (MPTP) induced loss of DA neurons. Striatal PPARδ is directly controlled by TLR/NFKB pathway and a PPAR response element is present in the AKKK1 promoter [64].
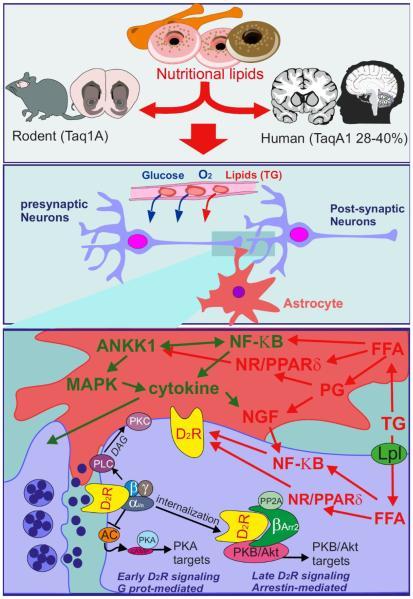
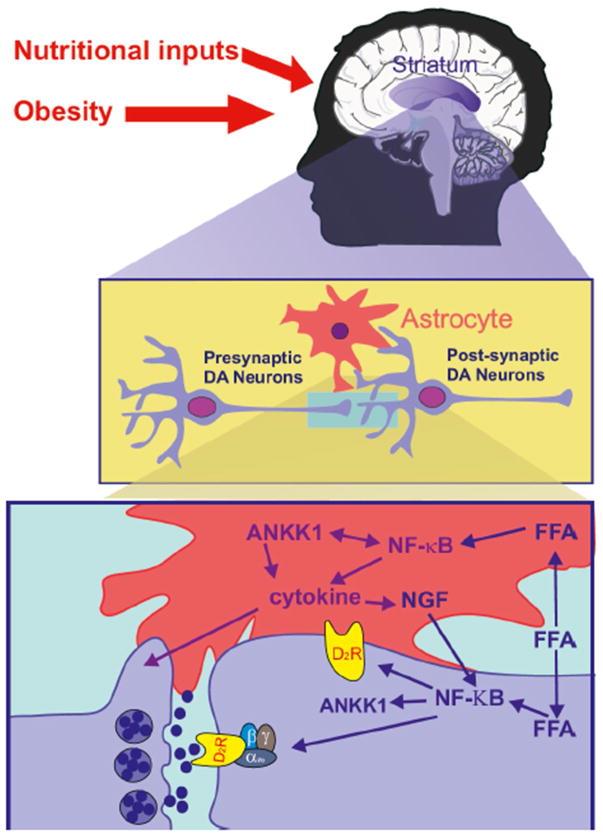
LCFA signaling in the ML might potentially be integrated at the level of the tripartite synapse composed of striatal DA neurons and astrocytes, busy in the coordinated activation of the ANKK1/NFκB/PPARs triad for bringing about the expression of function of D2R (Figure 3). Although the ML response to short term HFD exposure might be brought about by an NBκB/NGF activity on D2R. PPARδ activation by LCFA might cause long term transcriptional adaptations in striatal structures following HF intake. With this thought, changes in lipid sensing in the ML in addition to enhanced exposure to food associated cues, both effects of the modern food environment, would demonstrate along with heighten the result of the ANKK1 polymorphism in compulsive behavior.
Copyright© Kulvinder Kochar Kaur, et al.
Courtesy ref no-65-Nutritional lipids lead to increased synthesis of TG-rich particles and export by the gut. At the level of the brain, the tripartite synapse composed of neurons and astrocytes will detect changes in nutrient availability. Free fatty acids (FFA) enter astrocytes or neurons through lipid receptors/transporters or via lipoprotein lipase (LPL) mediated breakdown of lipoprotein. Once in the cell FFA can enter the TCA-cycle but can also directly activate lipid-activated nuclear receptors (NR) including proliferated activated receptors delta (PPARδ) or the nuclear enhancer of kappa-light- chain-enhancer of activated B cells (NF-κΒ) signaling pathways, or through indirect pathways involving Toll- like receptor activation, inflammatory processes, ER- stress, or prostaglandin (PG) synthesis. In turn, activated NR and NF-κB exert a direct regulation at the transcriptional level on dopamine D2 receptor (D2R). At the level of the astrocyte, NF-κB activation leads to increased cytokine production which promotes the release of the neural-growth factor (NGF). NGF released by astrocyte directly regulates neural D2R abundance. The astrocyte-specific Ankyrin repeat and kinase domain containing 1 (ANKK1) directly interacts with NF-κB and MAP kinases. In that regard, mutation of ANKK1 (TaqIA A1) would directly impact most FFA-induced cellular responses. At the level of DA neuron signaling, early D2R response and late β-arrestin mediated responses could both be affected by a FFA/ANKK1 initiated pathway.
Why Dopamine
In ingestive behavior role of DA involves multiple factors that include cognitive, appetitive along with metabolic domains along with their interactions. Together these domains collect information regarding the nutritive value and sensory characteristics of food along with the state of the organism, for giving priority and changing behavior to efficiently acquire and store energy. Subsequently many pathways exist through which genetic changes which impact DA signaling can affect food intake and obesity. Additionally new research positively indicates that adiposity, metabolic dysfunction along with diets high in saturated fat and sugar adapt in DA functions, are molecular, cellular as well as circuit level for affecting DA based functions varying from working memory and compulsive behaviors to preference of food and sensing of nutrients and glucose metabolism control [65, 66, 67, 68, 69, 70, 71]. Hence variants in DA genetically might affect intake besides by conferring initial risk, affecting brain adaptations [72]. Thus a vicious cycle gets initiated where genetic predisposition affect brain function for helping an unhealthy diet and weight increase in obesogenic Kulvinder Kochar Kaur, et al. Chronically Elevated Triglycerides as a Result of High Fat Palatable Diet Resulting in a Vicious Cycle on Reinforcing Reward and Dopamine Signaling: A Possible Cause for the Obesity Epidemic Worldwide in the Food Environment Available-A Comprehensive Review. J Endocrinol 2019, 3(2): 000143.
environments, that in turn effects function of the brain for promotion of both metabolic and cognitive dysfunction and more weight rise.
Role of DRD2
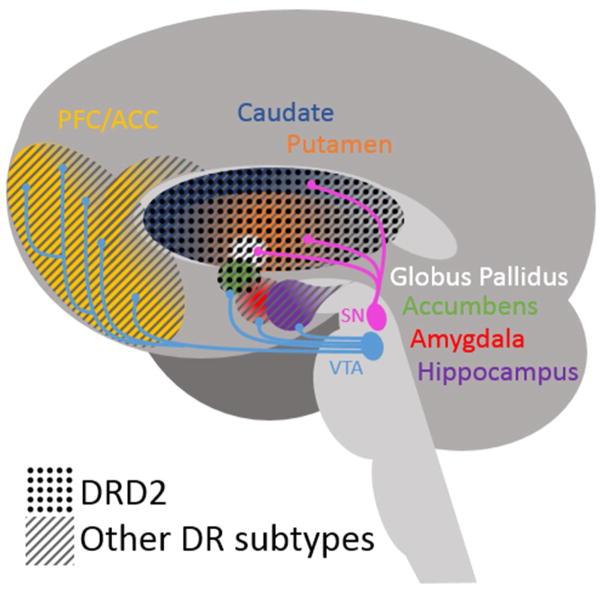
Multiple independent processes control DA signaling, which are DA formation, pre and post synaptic DA receptors and presynaptic DA transporters. Genetic differences influencing these processes might thus affect ingestive behavior and obesity. 5 types of DA receptors exist (DRD1-DRD5) that is basically grouped into 2 major subclasses: DRD2 like, that have DRD2, DRD3 and DRD4 and DRD1 like that include DRD1 and DRD5. Though they have various distribution patterns in the brain (Figure 4), they usually interact to control neurotransmission [73]. DRD2 is emphasized because of
- Differences in the fat mass and obesity associated (FTO) gene represent the most powerful polygenic adiposity determinants with inactivation of this gene impairing DRD2-dependent neurotransmission and function in rodents and DRD2 based learning in humans [74, 75, 76].
- The Taq1ARFLP that is related with differences in DRD2 receptor density [77] was demonstrated to interact with an FTO gene variant for affecting adiposity; both central along with peripheral insulin resistance (IR) as well as DRD2 based learning. Both studies showed the effect of FTO in these phenotypes was observed to be higher in persons, who also had a copy of the Taq1 A1at risk allele or based on persons also possessing this genotype.
- Though estimates differ based on ethnicity, approx.
50% (11-67%for rs8050136) of the European people carry an at risk allele for FTO and roughly 35% (23- 56% for rs 1800497) of these people can also be anticipated to carry the Taq1 A1 at risk allele (HapMap1 and 1000 Genomes2). Thus genetic variants affecting DRD2 signaling might affect a big portion of the population. • Lot of studies document reduced DRD2 after DIO, HFD in the absence of obesity, along with metabolic dysfunction.
Simplified schematic of brain dopamine pathways and distribution of DRD2 and other dopamine receptor subtypes in areas important for reward. DRD2 shows the greatest differential expression in striatal areas. SN = Substantia Nigra, VTA = Ventral Tegmental Area, PFC = Prefrontal Cortex, ACC = Anterior Cingulate Cortex.
Copyright© Kulvinder Kochar Kaur, et al.
DA-Based Functions Affecting Intake Behavior
A lot of cognitive functions get controlled by DA signaling, essential for optimizing Intake Behavior. The DA-fronto-striatal loop participates in working memory, cognitive flexibility along with associative learning [78, 79]. Particularly, working memory along with cognitive flexibility work as opposite effects for supporting online stability of task-related representations and helping the flexible update of these representations as an answer to novel input [80]. Thus any deficiencies in working memory along with cognitive flexibility in turn get related to cognitive inflexibility, impulsivity along with compulsive behaviors all being components of addictive like behaviors that include overeating [81].
Big debate exists regarding the exact role of DA in associative learning. One thing is known that various forms of associative learning are controlled by the fronto- striatal loop that includes:
- Model based learning, in which values for the future account for goal related behaviors
- Model free learning, in which earlier learned values cause habit development [82], big altercation on exact role of DA in these works exists. It has been said that DA regulates the exertion of effort [83], signaling of reward prediction errors [84], along with imparting incentive salience to reward associated stimuli [85]. Irrespective of the verified role that DA system imparts on moving from model based (like goal Kulvinder Kochar Kaur, et al. Chronically Elevated Triglycerides as a Result of High Fat Palatable Diet Resulting in a Vicious Cycle on Reinforcing Reward and Dopamine Signaling: A Possible Cause for the Obesity Epidemic Worldwide in the Food Environment Available-A Comprehensive Review. J Endocrinol 2019, 3(2): 000143.
directed/action-object responding) to model free (habitual stimulus-response resonding) learning after continuous exposure to reinforces like drugs as well as food [86], is a basic characteristic of addictive behavior [87] and seen in DIO.
DA projections from the VTA to hippocampus have been thought to be responsible for adaptive memory [88]. With more precision, this circuit helps in carving memories on the side of motivational importance so that whatever is recalled has motivational importance and is thus adaptive. If this change occurs in obesity remains a query, but HFD fed rodents have impairment of contextual memory tasks in which they need to remember getting food rewards present in different places with separate features [89].
DA has a key part in metabolic sensing [90]. Specially, it has been observed that DA function is crucial for the integrity of the ‘’gut-brain‘’ axis, that verifies the transfer of signals regarding the energy and nutrition given by nutrients to homeostatic and reward circuits that regulate their acquiring and metabolism. The “gut-brain” axis is the system where bidirectional communication takes place in circuits in the brain and cellular sensors in the GIT. Initially, ingestive behavior and nutrients metabolism were thought to be working with relative independence. But current work emphasizes on the close relationships with direct control of perception and behavior by peripheral signals with direct influence of central circuits that include DA neurons on nutrients metabolism.
Taq 1A Polymorphisms (rs 1800497)
Taq1A RFLP has been the most extensively explored genetic variant associated with DA Signaling along with obesity. 3 variants exist in humans, namely A1/A1,A1/A2 and A2/A2. Roughly 30 of European, 60% of Asian and 41% of African (Hapmap and 1000 Genomes) contain one or 2 copies of the A1 allele and those that do have approximately 30% fewer DRD2’s in the striatum, have been demonstrated by Candidate gene studies, although this was not replicated by one study [91]. A1 carriers have the possibility of having a waist circumference (WC) [92], BMI and obesity [93], as revealed by many researchers. Similarly GWAS has also suggested these variants in obesity. Associations with WC (p=0.01for men and p=0.07 for women in GIANT3), fasting glycaemia (p=0.02 in MAGIC4), insulin sensitivity (p=0.01 in MAGIC) and risk of T2DM (p=0,08) in DIAGRAM5). A meta-analysis of 33 studies having small samples (<100) in majority displayed no association between BMI and A1 allele status Copyright© Kulvinder Kochar Kaur, et al.
[94]. It does not come out to be astonishing, as this SNP gives an explanation for only a small percent of the variance in BMI of the population, that suggest larger samples, with measures of adiposity with more precision essential for examining trustable associations. Besides obesity, the A1 allele is related to a lot of disorders that have affected DA Signaling that are attention deficit hyperactivity disorder (ADHD), addiction, alcoholism [95, 96, 97]. Following traumatic brain injury bad results, and Parkinson’s disease [98, 99].
A1 carriers have decreased glucose metabolism widely at the neuro behaviour aspect, decreased grey matter density in the substantial nigra, sub thalamic nucleus and anterior cingulate cortex (ACC), and decreased activity in the prefrontal cortex and striatum at the time of reversal learning, working memory, and receipt of monetary reward [100, 101, 102, 103, 104]. In those who are A1 allele carriers have a greater relation with greater impulsivity, bad time estimation, steeper delayed discounting, bad working memory, affected reversal learning, impaired negative outcome learning and bad long term memory [105, 106, 107, 108].
These endophenotypes in turn are related with overeating and risk of obesity. Like carriers in contrast to noncarriers will work for food, especially if also obese, and the ones high in food reinforcement take more snack food vs. non carriers and A1 carriers who show low food reinforcement [109]. Additionally in a cohort of mainly healthy weight Asian American college students, A1 carriers compared to non-carriers experienced higher fast-food and carbohydrate craving and in case of females further there was higher urge of taking highly tasty food [110]. Moreover in carriers but not in noncarriers a positive correlation between response in the OFC to tasty food cues and further gain in body fat [111]. But carriers also display decreased responses in DA source and target regions to the receipt or imagined delivery of an anticipated tasty food, with these responses further predicting future weight gain [111, 112] (Figure 5).
Nutritional inputs such as free fatty acids (FFA) promote the action of pro-inflammatory signaling in astrocytes and neurons. This results in activation of NF- kB (the nuclear enhancer of kappa-light-chain-enhancer of activated B cells). Through astrocyte-neuronal signaling NF-kB regulates the transcription of the DA D2 receptor (D2R). In addition, within the astrocyte NF-kB signaling promotes a cytokine-mediated production of the neural-growth factor (NGF), a potent inducer of neural D2R abundance. ANKK1 is predicted to physically interact with NF-kB in the astrocyte and in the neuron. As such, Kulvinder Kochar Kaur, et al. Chronically Elevated Triglycerides as a Result of High Fat Palatable Diet Resulting in a Vicious Cycle on Reinforcing Reward and Dopamine Signaling: A Possible Cause for the Obesity Epidemic Worldwide in the Food Environment Available-A Comprehensive Review. J Endocrinol 2019, 3(2): 000143.
ANKK1 mutations (TaqIA A1) could directly impact cellular responses mediated by NF-kB, including regulation of D2R signaling and receptor expression). The circles in the cleft represent dopamine release.
This fashion of effects seen has been construed as proof of increased anticipatory and depressed consummatory food reward, supporting the incentive sensitization theory of addiction, along with the anhedonia posit of obesity. But the current data has refuted this observation about decreased responses pointing to anhedonia. 1stly, dorsal striatal response to milk shake is related to measures of impulsive behavior but not of reward of food or reinforcement [113]. 2nd, a positive correlation between weight gain and dorsal striatal milk shake response in A1 carriers but not non carriers was documented recently [114]. Most important, is that milkshake was delivered in an unpredictable way in this particular study. Hence response to that very food, in that very DRD2 rich nucleus has positive or negative correlation with weight gain in A1 carriers based on if milkshake is anticipated or not anticipated. This information requires emphasis since unanticipated but Copyright© Kulvinder Kochar Kaur, et al.
not anticipated outcomes produce error signals. It has been indicated that the reduced effect to anticipated milkshake agrees with a weaker outcome signal, although the response to unanticipated milkshake points to an increased error signal. In the same breath it is hypothesized that the escalated responses to food cues, suggest total increased reward sensitivity in case of obesity [115].
Together it is posited that the A1 allele is correlated with increased error, but milder reward outcome signal production in the relation of total increased reward sensitivity with all these observations. Another publication showed increased midbrain response in A1 carriers vs. noncarriers on the production of positive error signals at the time of probabilistic learning task that supported the earlier study. Further this study demonstrated that the accelerated response was related to poorer performance in the case of negative outcome learning that would be anticipated if reward outcome signals were reduced. Further carriers did not show functional connectivity between midbrain and medial prefrontal cortex at the time of the tasks, implicating a decreased transfer of DA learning signals to prefrontal circuits. Similarly weaker reward response signals would also be anticipated to enhance chances for model free or habitual responding.
From these results strong proof is also added regarding A1 allele provides risk by generating baseline reductions in DA signaling causing variations in DA dependent reinforcement and cognition. Going with this probability pharmacological manipulation of DA signaling often gets separate but opposite changes in carriers as compared to noncarriers. Like methyl phenidate, that raises extracellular DA, has less efficacy in reducing food intake in A1 carrier vs A1 noncarriers children in ADHD, while the DRD2 agonist bromocriptine is more efficacious at decreasing craving and anxiety in A1 allele carrying alcoholics [116, 117]. In healthy people, bromocriptine, and not placebo administration raises activity in the nucleus accumbens at the time of a reinforcement task and increases performance in A1 carriers but not noncarriers. Giving cabergoline, another agonist at the time of reversal learning task resulted in increased activation of the medial orbito frontal cortex (OFC), insula, ventral putamen and ACC in A1 carriers but reduces response in these very areas in non-carriers. This implicates that like DA agonists, behavioral measures which enhance DA signaling, like exercise might be more efficacious in A1 carriers. In total, these results agree with the inverted U shaped action of DA function Kulvinder Kochar Kaur, et al. Chronically Elevated Triglycerides as a Result of High Fat Palatable Diet Resulting in a Vicious Cycle on Reinforcing Reward and Dopamine Signaling: A Possible Cause for the Obesity Epidemic Worldwide in the Food Environment Available-A Comprehensive Review. J Endocrinol 2019, 3(2): 000143.
superimposed on baseline differences in DA signaling given by genotype. Precisely the action of DA agonists would shift A1 carriers from suboptimal (low) to optimal (intermediate) and A1 noncarriers from optimal (intermediate) to suboptimal (high) dopaminergic functioning [8107], and hence get opposite effects.
Longitudinal studies data that examined effect of Taq1A A1RFLP on weight increase agree with the pharmacological data, showing opposite correlations between responses to food in DA source or target regions and weight gain as a function of genotype. More precisely, response of caudate to predicted milkshake receipt is negatively correlated with weight gain in carriers, but positively correlated in non-carriers, while the reverse is true for amygdalar response to milkshake in sated state. It is not clear if these actions are a cause or effect of obesity. But the dorsal striatal effects have a possibility of being a result, as decreased responses correlate with weight rise but not with risk for obesity as checked by parental BMI [118]. But, as response to milkshake in this area is correlated with impulsivity, it is likely that A1 allele imparts risk for adaptations in striatal circuits that in turn cause impulsivity. If that is the case, then 1 of the actions of the allele might be to increase risk for neural adaptations which result in impulsivity and increase of weight increase that might explain why carriers have decreased outcomes with weight loss interventions [119]. It is feasible in relation that adaptations take place as a result of factors separate from diet, adiposity or metabolic change like age, poor sleep quality, chronic stress and physical inactivity [120]. In agreement with this thought, old although not young carriers demonstrate decreased dorsal striatal response and a memory task that is impaired [121].
Molecular Mechanisms
The Taq1A RFLP has a correlation with a mutation generating a single amino acid alteration in the substrate binding domain of the ANKK1 protein and its linkage disequilibrium with the DRD2 locus. Exactly how this mutation in theAnkk1 gene locus alters DRD2 density and function is under investigation one probability is that the mutation affects ANKK1 action that subsequently affects DA receptors via its biochemical actions. ANKK1 represents a big family of receptor-interacting protein (RIP) kinases, which act as necessary sensors of cellular stress, starting responses to environmental factors, like nutrient intake via activation of transcription factors like NFκB. Chronic exposure to increased nutritional lipids is instigate inflammatory-like response in the brain ,which Copyright© Kulvinder Kochar Kaur, et al.
are partially mediated by NFκB, that in response, acts as a direct along with indirect transcriptional regulator of DA receptor amount and signaling. Consistently, in silico analysis of the possible human protein-protein interactions show that of the 30~ anticipated possible partners of human ANKK1 protein [122], 50% are found in pathways associated with inflammatory responses that include the NFκB, cytokine pathways. Particularly in the brain ANKK1 is highly represented, if not only specially expressed in astrocytes [123]. Astrocytes might be the targets for highly saturated fat stimulated inflammatory responses and ER stress mediated via TLR and IKK/NFκB signaling pathways. Additionally eicosanoids derived from lipids are strong inducers of NGF secretion by astrocytes that also act directly for control of DRD2 expression via an NFκB dependent mechanism. Further HFD-mediated central inflammation enhances DA neurons susceptibility to 1methyl-4phenyl-12,3,6 tetra hydropyridine (MPTP) [124, 125]. Hence there are modes that can verify that a privileged association between astrocytes and DRD2 containing neurons causing the selective actions of lipids on DRD2 through inflammatory cascades, just like the ones already revealed for hypothalamus [126]. Hence probably the mutation of ANKK1 brings about the inflammatory-like responses which particularly influence DRD2 excess and function, although some proof exists that other DA receptor subclasses might be also influenced.
Model Proposed
The above information points that a probability is present that ANKK1 has an important part in bringing about the effect of HFD on DRD2 expression through the nutrient stimulated inflammatory responses. Thus a model was posited by the group of Small DM, in which presence of A1 allele increases susceptibility to cellular inflammatory responses that further changes DA signaling along with DA dependent cognition, metabolism and behavior. Since DA has a major part in intertwining inputs regarding the nutritional value along with sensory features of foods, in addition to the state of the person for decision and act for achieving and store energy, multiple probable pathways through which adaptations in DA might add risk to overeat and thus for obesity [127]. Consistently, obese vs. lean carriers but not noncarriers exhibit decreased performance on a letter number sequencing task which checks the executive functions [128]. Thus by extending this DA signaling adaptations might further add the risk for other impulse control disorders and problems in which changed DA signaling acts in causing etiopathology, like DM, and ADHD.
Kulvinder Kochar Kaur, et al. Chronically Elevated Triglycerides as a Result of High Fat Palatable Diet Resulting in a Vicious Cycle on Reinforcing Reward and Dopamine Signaling: A Possible Cause for the Obesity Epidemic Worldwide in the Food Environment Available-A Comprehensive Review. J Endocrinol 2019, 3(2): 000143.
Important take home message of this is that ANKK1 is mainly connected to diet instead of adiposity or metabolic dysfunction. From this angle correlation between genotype and BMI and adiposity are due to the action of diet. In the same way this model anticipates that a HFD also causes alterations in DA dependent cognition through affecting DRD2. Importance of this lies in the proof being unraveled that DM, obesity and diet are correlated with cognitive impairments, with so many of these abnormalities being dependent on DA signaling [129]. No research has directly assessed the effect of diet on the correlation between the Taq1A A1 allele and adiposity, metabolic and /or cognitive dysfunction. But from animal experiments data reflect that diet might affect DA signaling and function unrelated to body weight, adiposity, and /or metabolic dysfunction [66, 70, 130]. In accord, circulating lipids directly modulate DA-mediated food reward and obesity-related cognitive dysfunction gets improved by decreasing circulating lipids independent of adiposity.
Conclusion
Although till now it is unexplained how TG reaches the brain and alters its function, same is the case with the molecular methods by which local FFA increase by breakdown of these TG’s and modulates neural effects along with reward. Both direct along with indirect proof in both humans and animals exist, as per the action of diet TG on reward and motivation. Hydrolysis of TG’s locally in the striatum might inhibit locomotor activity and for a little time decrease the incentive salience of calorie rich HFHS foods. But in the presence of chronically escalated TG’s in plasma, that is the result of the current scenario of food prevalence and obesity, the natural homeostatic mechanisms that usually reduce the hedonic effect of HFHS fails. From that angle, a positive feedback loop in which a chronic enhanced high amounts of TG’s, like that found in obesity, damages the homeostatic processes that limit food intake and changes in reward encoding, uncontrolled calories consumption and decreased physical activity. This will definitely push towards gain in weight. More studies are needed for understanding the physiology and molecular modes of sensing of TG’s in the brain and if or how genetic susceptibility loci like the Taq1 A allele might exacerbate the adaptive methods related to brain TG sensing and in the end the down ward trend compulsive eating that is dissociated from the homeostatic requirements. Further Wei, et al. demonstrated that chronic stress aggravates the susceptibility to food addiction by increasing the CRF R1 in the NAc shell and core but reduced in VTA in mice fed a Copyright© Kulvinder Kochar Kaur, et al.
highly palatable food. It increased FAS along with levels of DR2 and MOR IN THE NAc. Thus dysregulation of the CRF signaling pathway in the reward system and increased expression of DR2 and MOR contributes to stress related obesity [131]. Although it is tempting to posit that the 2 common genetic variants, ANKK1 and FTO confer risk for obesity and neurocognitive impairment by their effects on DRD2 the recent in depth articles by the group of Archer, et al. are contradictory to the genetic and addictive theory by their explanation of compartmentalization and how they have given convincing explanations regarding it is not the food addiction theory nor the genetic theory but how that materno-fetal metabolic exposure and the DoHaD Hypothesis causes the neonate to acquire higher adipocytes and poor quality muscle cells and in the end we are back to square one in trying to finding the answers of finding a solution to put a brake in the obesity epidemic [132, 133, 134, 135]. Clinical implications of these lie in trying to better utilize naltrexone/bupropion (contrive) in those who are Taq1 A allele carriers, and do genetic typing like in a case report Digbi Health Personalized Obesity Management program use of DNA and gut biome along with testing. Participants use the Digbi Health app to input 10 key lifestyle and wellness markers, including weight, sleep, hunger, cravings, stress, meditation, super foods, morning energy, foods to avoid and exercise on a daily basis and take photos of the food that they consume. As per these markers they decide whether exercise will work or not and as per genetic profile gut biome treatment is finalized and are able to achieve weight loss based on individualization of subjects, thus in those having abnormal TaqA1 alleles use of DA agonists like bromocriptine/cabergoline and behavioral measures which enhance DA signaling, like exercise might be more efficacious in A1 carriers and preferably use of contrave for obesity treatment in these subjects along with contrave use.
References
-
Kulvinder Kochar Kaur, Gautam Allahbadia, Mandeep Singh (2013) Current Management of Obesity in an Infertile Female-Recent Advances and Future Prospective Drugs. Journal of Pharmacy and Nutrition Sciences 3(3): 178-190.
-
Kulvinder Kochar Kaur, Gautam Allahbadia, Mandeep Singh (2016) An Update on a Etiopathogenesis and Management of Obesity. Obes Control Ther P 3(1): 1- 17. Kulvinder Kochar Kaur, et al. Chronically Elevated Triglycerides as a Result of High Fat Palatable Diet Resulting in a Vicious Cycle on Reinforcing Reward and Dopamine Signaling: A Possible Cause for the Obesity Epidemic Worldwide in the Food Environment Available-A Comprehensive Review. J Endocrinol 2019, 3(2): 000143.
-
Kulvinder Kochar Kaur, Gautam Allahbadia, Mandeep Singh (2016) Further update on the management of Obesity with Emphasis on Genetic Perspective. BAOJ Obe Weigt Manage 3(10): (1-11).
-
Kulvinder Kochar Kaur, Confused hypothalamus-A cause for obesity worldwide. Gynecological Endocrinology Abstract P132; 2006: 22(Suppl 1) Book of Abstracts for the 12th world congress of gynaecological endocrinology, Florence, Italy.
-
Kochar Kaur K, Role of dysregulation of autonomic nervous system versus altered hypothalamo- pituitary-adrenal (H-P-A) axis in drug induced obesity 7th IBRO World Congress of Neurosciences, Melbourne Australalia12-17 july 2007.Book of Abstracts Neuroendocrinology Sun 092-13-14: 214.
-
Kulvinder Kochar Kaur, Allahbadia G, Singh M (2017) Hypothalamic inflammation and glioses as aetiopathogenetic factor in high fat diet induced obesity and various therapeutic options to resolve it. Obes Res Open J 4(2): 44-60.
-
Kulvinder Kochar Kaur, Allahbadia G, Singh M (2019) Therapeutic Utilization of Neuro Imaging Studies in Obesity for Optimal Utilization of Drugs used in Treatment for Obesity-Lessons Learnt from Bariatric Surgery. J Ageing Restor Med 2(2): 89-97.
-
Di Chiara G, Imperato A (1988) Drugs abused by humans preferentially increase synthetic dopamine concentrations in the mesolimbic system of freely moving rats. Proc Natl Acad Sci USA 85(14): 5274- 5278.
-
Dallman MF, Pecoraro NC, La Fleur SE (2005) Chronic stress and comfort foods: self-medication and abdominal obesity. Brain Behav Immunol 19(4): 275- 280.
-
Stouffer MA, Woods CA, Patel JC, Lee CR, Witkovsky P, et al. (2015) Insulin enhances striatal dopamine release by activating cholinergic interneurons and thereby signaling rewards. Nat Commun 6: 8543.
-
Blouet C, Schwartz GJ (2010) Hypothalamic nutrient sensing in the control of energy homeostasis. Behav Brain Res 209(1): 1-12.
-
Lam TK, Schwartz GJ, Rossetti L (2005) Hypothalamic sensing of fatty acids. Nat Neurosci 8(5): 578-584. Copyright© Kulvinder Kochar Kaur, et al.
-
Milanski M, Degasperi G, Coope A, Morari J, Denis R, et al. (2009) Saturated fatty acids produce an inflammatory response predominantly through the activation of TLR4 signaling in hypothalamus: implications for the pathogenesis of obesity. J Neurosci 29(2): 359-370.
-
Gao Y, Ottaway N, Schriever SC, Legutko B, Garcia- Caceres C, et al. (2014) Hormones and diet, but not body weight, control hypothalamic microglial activity. Glia 62(1): 17-25.
-
Lopez M, Tovar S, Vasquez MJ, Nogueiras R, Senaris R, et al. (2005) Sensing the fat: fatty acid metabolism in the hypothalamus and the melanocortin system. Peptides 26(10): 1753-1758.
-
Volkow ND, Wang GJ, Telang F, Fowler JS, Thanos PK, et al. (2008) Low dopamine striatal D2 receptors are associated with prefrontal metabolism in obese subjects: possible contributing factors. Neuroimage 42(4): 1537-1543.
-
Johnson PM, Kenny PJ (2010) Dopamine D2 receptors in addiction-like reward dysfunction and compulsive eating in obese rats. Nat Neurosci 13(5): 635-641.
-
Robinson MJ, Burghardt PR, Patterson CM, Nobile CW, Akil H, et al. (2015) Individual differences in Cue- induced Motivation and Striatal System in rats Susceptible to diet induced obesity. Neuropsychopharmacology 40(9): 2113-2123.
-
Wang GJ, Volkow ND, Logan J, Pappas NR, Wong N, et al. (2001) Brain dopamine and obesity. Lancet 357(9253): 354-357.
-
Stice E, Spoor S, Bohon C, Veldhuizen MG, Small DM (2008) Relation of reward from food intake and anticipated food intake in obesity: a functional magnetic resonance imaging study. J Abnorm Psychol 117(4): 924-935.
-
Guo J, Simmons WK, Herscovitch P, Martin A, Hall KD (2014) Striatal Dopamine D2 -like receptor correlation patterns with human obesity and opportunistic eating behavior. Mol Psychiatry 19: 1078-1084.
-
South T, Huang XF (2008) High fat diet exposure increases Dopamine D2 receptor and decreases dopamine transporter receptor binding density in the Kulvinder Kochar Kaur, et al. Chronically Elevated Triglycerides as a Result of High Fat Palatable Diet Resulting in a Vicious Cycle on Reinforcing Reward and Dopamine Signaling: A Possible Cause for the Obesity Epidemic Worldwide in the Food Environment Available-A Comprehensive Review. J Endocrinol 2019, 3(2): 000143. nucleus accumbens and caudate putamen of mice. Neurochem Res 33(3): 598-605.
-
Adams WK, Sussman JL, Kaur S, D’Souza AM, Kieffer TJ, et al. (2015) Longterm, calorie restricted intake of a High fat diet in rats reduces impulse control and ventral striatal D2 receptor signaling: two markers of addiction vulnerability. Eur J Neurosci 42(12): 3095- 3104.
-
Valdivia S, Conejo MP, Reynaldo M, De Francesco PN, Perello M (2015) Escalation in High fat intake in a binge eating model differentially engages dopamine neurons of the ventral tegmental area and requires ghrelin signaling. Pschyconeuroendocrinology 60: 206-216.
-
Hryhorczuk C, Florea M, Rodaros D, Poirier J, Daneault C, et al. (2015) Dampened mesolimbic dopamine function and signaling by Saturated but not Monounsaturated Dietary Lipids. Neuropsychopharmacology 41(3): 811-821.
-
Dourmashkin JT, Chang CO, Hill JO, Gayles EC, Fried SK, et al. (2006) Model for predicting and phenotyping at normal weight the long-term propensity for obesity in Sprague-Dawley rats. Physiol Behav 87(4): 666-678.
-
Karatayev O, Gaysinkaya V, Chang GO, Leibowitz SF (2009) Circulating triglycerides after a high fat meal: predictor of increased caloric intake, orexigenic peptide expression, and dietary obesity. Brain Res 1298: 111-122.
-
Rada P, Avena NM, Barson JR, Hoebel BG, Leibowitz SF (2012) A high fat meal, or intraperitoneal administration of a fat emulsion, increased extracellular dopamine in the nucleus accumbens. Brain Sci 2(2): 242-253.
-
Farr SA, Yamada KA, Butterfield DA, Abdul HM, Xu L, et al. (2008) Obesity and hypertriglyceridemia produce cognitive impairment. Endocrinology 149(5): 2628-2636.
-
Sun X, Veldhuizen MG, Wray AE, De Araujo IE, Sherwin RS, et al. (2014) The neural signature of satiation is associated with ghrelin response and triglyceride metabolism. Physiol Behav 136: 63-73.
-
Edmond J (2001) Essential poly unsaturated fatty acids and the barrier of the brain: the components of Copyright© Kulvinder Kochar Kaur, et al. a model for transport. J Mol Neurosci 16(2-3): 181- 193.
-
Rapoport SI, Chang MC, Spector AA (2001) Delivery and turnover of plasma-derived essential PUFA’s in mammalian brain. J Lipid Res 42(5): 678-685.
-
Wang H, Eckel RH (2014) What are lipoproteins doing in the brain?. Trends Endocrinol Metab 25(1): 8-14.
-
Rapoport SI (2001) In vivo fatty acids incorporation into brain phospholipids in relation to plasma availability, signal transduction and membrane remodeling. J Mol Neurosci 16(2-3): 243-261.
-
Karmi A, Iozzo P, Viljanen A, Hirvonen J, Fielding BA, et al. (2010) Increased brain fatty acids uptake in metabolic syndrome. Diabetes 59(9): 2171-2177.
-
Nguyen LN, Ma D, Shui G, Wong P, Cazenave –Gassiot A, et al. (2014) Mfsd2a is a transporter for the essential omega 3 fatty acid docosa hexaenoic acid. Nature 509(7501): 503-506.
-
Ouellet M, Emond V, Chen CT, Julien C, Bourasset F, et al. (2009) Diffusion of docosahexaenoic and eicosapentaenoic acids through the blood brai barrier: An in situ cerebral perfusion study. Neurochem Int 55(7): 476-482.
-
Koch S, Donarski N, Goetzke K, Kreckel M, Stuerenberg HJ, et al. (2001) Characterisation of four lipoprotein classes in human cerebrospinal fluid. J Lipid Res 42(7): 1143-1151.
-
Louveau A, Smirmov I, Keyes TJ, Eccles JD, Rouhani SJ, et al. (2015) Structural and functional features of central nervous system lymphatic vessels. Nature 523(7560): 337-341.
-
Evola M, Hall A, Wall T, Young A, Grammas P (2010) Oxidative stress impairs learning and memory in apoE knockout mice. Pharmacol Biochem Behav 96(2): 181-186.
-
De Oliveira J, Moriera EL, Dos Santos DB, Piermartiri TC, Dutra RC, et al. (2014) Increased susceptibility to amyloid beta induced neurotoxicity in mice lacking the low density lipoprotein receptor. J Alzheimers Dis 41(1): 43-60.
-
Borroni B, Grassi M, Agosti C, Costanzi C, Archetti S, et al. (2006) Genetic correlates of behavior Kulvinder Kochar Kaur, et al. Chronically Elevated Triglycerides as a Result of High Fat Palatable Diet Resulting in a Vicious Cycle on Reinforcing Reward and Dopamine Signaling: A Possible Cause for the Obesity Epidemic Worldwide in the Food Environment Available-A Comprehensive Review. J Endocrinol 2019, 3(2): 000143. endophenotypes in Alzheimers Disease: Role of COMT-5HTTLPR and APOE Polymorphisms. Neurobiol Ageing 27(11): 1595-1603.
-
Wang H, Eckel RH (2012) Lipoprotein Lipase in the brain and nervous system. Annu Rev Nutr 32: 147- 160.
-
Yu T, Taussig MD, Di Patrizio NV, Astarita G, Piomelli D, et al. (2015) Deficiency of Lipoprotein Lipase in Neurons Decreases AMPK Receptor Phosphorylation and leads to Neurobehavioral Abnormalities in Mice. PLoSOne 10(8): e0135113.
-
Cansell C, CasteL J, Denis RG, Rouch C, Delbes AS, et al. (2014) Dietary triglycerides act on mesolimbic structures to regulate the rewarding and motivational aspects of feeding. Mol Psychiatry 19(10): 1095-1105.
-
Wang H, Astarita G, Taussig MD, Bhardwaj KG, Di Patrizio NV, et al. (2011) Deficiency of Lipoprotein Lipase in neurons modifies the regulation of energy balance and leads to obesity. Cell Metab 13(1): 105- 113.
-
Picard A, Rouch C, Kassis N, Moulle VS, Croizier S, et al. (2014) Hippocampal Lipoprotein Lipase regulates energy balance in rodents. Molecular Metab 3(2): 167-176.
-
Ruge T, Hodson L, Cheseman J, Dennis AL, Fielding BA, et al. (2009) Fasted to fed tracking of fatty acids in human adipose tissue reveals a novel regulatory step for enhancing fat storage. J Clin Endocrinol Metab 94(5): 1781-1788.
-
Bjursell M, Gerdin AK, Lelliot CJ, Egecioglu E, Emelgren A, et al. (2008) Acutely reduced locomotor activity is a major contributor to Western diet induced obesity in mice. Am J Physiol Endocr Metab 294(2): 251-260.
-
Oomura Y, Nakamura T, Sugimori M, Yamada Y (1975) Effect of free fatty acid on the rat lateral hypothalamic neurons. Physiol Behav 14(4): 483-486.
-
Obici S, Feng Z, Morgan K, Stein D, Karkanias G, et al. (2002) Central administration of oleic acid inhibits glucose production and food intake. Diabetes 51(2): 271-275.
-
Prouix K, Cota D, Woods SC, Seely RJ (2008) Fatty acid Synthase inhibitors modulate energy balance via Copyright© Kulvinder Kochar Kaur, et al. mammalian target of rapamycin complex1 signaling in central nervous system. Diabetes 57(12): 3231- 3238.
-
Milanski M, Arruda AP, Coope A, Ignacio-Souza LM, Nunez CE, et al. (2012) Inhibitors of hypothalamic inflammation reverses diet induced insulin resistance in the liver. Diabetes 61(6): 1455-1462.
-
Andrews ZB, Liu ZW, Wallingford N, Erion DB, Brook E, et al. (2008) UCP2 mediates ghrelin’s action on NPY/AgRP neurons by lowering free radicals. Nature 454(7206): 846-851.
-
Yue JT, Abraham MA, LaPierre MP, Mighiu PI, Light PE, et al. (2015) A fatty acid-dependent hypothalamic DVC neurocircuitry that regulates hepatic secretion of triglyceride rich lipoprotein. Nat Commun 6: 5970.
-
Zhang L, Reith ME (1996) Regulation of functional activity of human dopamine transporter by the arachidonic acid pathway. Eur J Pharmacol 315(3): 345-354.
-
Stice E, Yokum S, Bohon C, Marti N, Smolen A (2010) Reward circuitry responsitivity to food predicts future increases in body mass: moderating effects of DRD2 and DRD4. Neuroimage 50(4): 1618-1625.
-
Hoenicka J, Quinones-Lombrana A, Espana-Serrano L, Alvira-Bottero X, Kremer L, et al. (2010) The ANKK1 gene associated with addictions is expressed in astroglial cells and upregulated by apomorphine. Biol Psychiatry 67(1): 3-11.
-
Meylan E, Tschopp J (2005) The RIP Kinases: crucial integrators of cellular stress. Trends Biochem Sci 30(3): 151-159.
-
Kotlyar M, Pastrello C, Pivetta F, Lo Sarda A, Cumbaa C, et al. (2015) In siico prediction of physical protein interactions and characterization of intractome orphans. Nat Methods 12(1): 79-84.
-
Buckman LB, Thompson MM, Lippert RN, Blackwell TS, Yull FE, et al. (2015) Evidence for a novel functional role of astrocytes in the acute homeostatic response to high fat diet intake in mice. Mol Metab 4(1): 58-63.
-
Bontempi S, Fiorentini C, Busi C, Guerra N, Spano P, et al. (2007) Identification and characterization of two nuclear factor kappa sites in the regulatory region of Kulvinder Kochar Kaur, et al. Chronically Elevated Triglycerides as a Result of High Fat Palatable Diet Resulting in a Vicious Cycle on Reinforcing Reward and Dopamine Signaling: A Possible Cause for the Obesity Epidemic Worldwide in the Food Environment Available-A Comprehensive Review. J Endocrinol 2019, 3(2): 000143. the Dopamine D2 receptor. Endocrinology 148(5): 2563-2570.
-
Fiorentini C, Guerra N, Facchetti M, Finardi A, Tiberio L, et al. (2002) Nerve growth factor regulates Dopamine D(2) receptor expression in prolactinoma cell lines via p75(NGFR)-mediated activation of nuclear factor-kappa B. Mol Endocrinol 16(2): 353- 366.
-
Christyakov DV, Aleshin S, Sergeeva MG, Reiser G (2014) Regulation of peroxisome proliferator activated receptor beta/delta expression and activity levels by toll like receptor agonists and MAP Kinase inhibitors in rat astrocytes. J Neurochem 130(4): 563- 574.
-
Berland C, Cansell C, Hnasko TS, Magnan C, Luquet S (2016) Dietary triglycerides as signaling molecules that influence reward and motivation. Curr Opin Behav Sci 9: 126-135.
-
Adams WK, Sussman JL, Kaur S, D'souza AM, Kieffer TJ, et al. (2015) Longterm, calorie-restricted intake of high fat diet in rats reduces impulse control and ventral striatal D2 receptor signaling-two markers of addiction vulnerability. Eur J Neurosci 42(12): 3095- 3104.
-
Johnson PM, Kenny PJ (2010) Dopamine D2 receptors in addiction-like reward dysfunction and compulsive eating in obese rats. Nat Neurosci 13(5): 635-641.
-
Coppin G, Nolan-Poupart S, Jones-Gotman M, Small DM (2014) Working memory and reward association learning impairments in obesity. Neuropsychologia 65: 146-155.
-
Bauer LO, Manning KJ (2016) Challenges in the detection of Working Memory and Attention Decrements among overweight Adolescent Girls. Neuropsychobiology 73(1): 43-51.
-
Han W, Tellez LA, Niu J, Medina S, Ferreira TL, et al. (2016) Striatal Dopamine links Gastrointestinal Rerouting on Altered Sweet Appetite. Cell Metab 23(1): 103-112.
-
Diepenbroek C, van der Plasse G, Eggels L, Rijnsburger M, Feenstra MGP, et al. (2013) Alterations in blood glucose and plasma glucagon concentrations during deep brain stimulation in the Copyright© Kulvinder Kochar Kaur, et al. shell region of nucleus accumbens in rats. Front Neurosci 7: 226.
-
Stice E, Dagher A (2010) Genetic variations in Dopaminergic reward in humans. Forum Nutr 63: 176-185.
-
Jackson DM, Westlind-Danielsson A (1994) Dopamine-receptors-Molecular Biology, Biochemistry and Behavioral-Aspects. Pharmacology and Therapeutics 64(2): 291-370.
-
Frayling TM, Timpson NJ, Weedon MN, Zeggini E, Freathy RM, et al. (2007) A common variant in the FTO gene is associated with body mass index and predisposes to childhood and adult obesity. Science 316(5826): 889-894.
-
Hess ME, Hess S, Meyer KD, Verhagen LAW, Koch L, et al. (2013) The fat mass and obesity associated gene (Fto) regulates activity of the dopaminergic midbrain circuitry. Nat Neurosci 16: 1042-1048.
-
Sevgi M, Rigoux L, Kühn AB, Mauer J, Schilbach L, et al. (2015) An Obesity-Predisposing Variant of the FTO Gene regulates D2R-Dependent Reward Learning. J Neurosci 35(36): 12584-12592.
-
Richie T, Noble EP (2003) Association of seven polymorphisms of the D2 Dopamine receptor gene with brain receptor-binding characteristics. Neurochem Res 28(1): 73-82.
-
Heni M (2016) Evidence for an interaction between the obesity risk gene FTO and the Dopamine D2 receptor gene ANKK/Taq1A on insulin sensitivity. Diabetologia.
-
Frank MJ, Fossella JA (2011) Neurogenetics and pharmacology of learning, motivation, and cognition. Neuropsychopharmacology 36(1): 133-152.
-
Cools R, D’Esposito M (2011) Inverted U shaped dopamine actions on human working memory and cognitive control. Biol Psychiatry 69(12): e113-125.
-
Guo J, Simmons WK, Herscovitch P, Martin A, Hall KD (2014) Striatal Dopamine D2 like receptor correlation patterns and human obesity and opportunistic earing behavior. Mol Psychiatr 19(10): 1078-1084.
-
Dayan P, Berridge KC (2014) Model based and model free Pavlovian reward learning: revaluation, revision Kulvinder Kochar Kaur, et al. Chronically Elevated Triglycerides as a Result of High Fat Palatable Diet Resulting in a Vicious Cycle on Reinforcing Reward and Dopamine Signaling: A Possible Cause for the Obesity Epidemic Worldwide in the Food Environment Available-A Comprehensive Review. J Endocrinol 2019, 3(2): 000143. and revelation. Cogn Affect Behav Neurosci 14(2): 473-492.
-
Salamone JD (2007) Functions of mesolimbic dopamine: changing concepts and shifting paradigms. Psychopharmacology 191(3): 389-389.
-
Schultz W (2016) Dopamine reward prediction-error signaling: a2 component response. Nature Reviews Neurosci 17(3): 183-195.
-
Berridge KC (1996) Food reward: Brain substrate of wanting and liking. Neurosci and Biobehavioral Reviews 20(1): 1-25.
-
Voon V, Derbyshire K, Rück C, Irvine MA, Worbe Y, et al. (2015) Disorders of Compulsivity: a common bias towards learning habits. Mol Psychiatr 20(3): 345- 352.
-
Everitt BJ, Robbins TW (2013) From the ventral to the dorsal Striatum: Developing views of their roles in drug addiction. Neurosci Biobehav Rev 37(9 Pt A): 1946-1954.
-
Sohamy D, Adcock RA (2010) Dopamine and Adaptative memory. Trends Cogn Sci 14(10): 464- 472.
-
Kanoski SE, Davidson TL (2011) Western diet consumption and cognitive impairment: links to hippocampus dysfunction and obesity. Physiol Behav 103(1): 59-68.
-
deAraujo IE, Ren X, Ferreira JG (2010) Metabolic sensing in brain Dopamine systems. Results Probl Cell Differ 52: 69-86.
-
Larulle M, Gelernter J, Innis RB (1998) D2 receptor binding potential is not affected by Taq1 polymorphism in the D2 receptor gene. Mol Psychiatr 3(3): 261-265.
-
Neville M, Johnstone EC, Walton RT (2004) Identification and characterization of ANKK1: a novel kinase gene closely linked to DRD2 on chromosome bad11q 23.1. Hum Mut 23(6): 540-545.
-
Noble EP, Noble RE, Ritchie T, Syndulko K, Bohlman MC, et al. (1994) D2 Dopamine receptor gene and obesity. Int J Eat Disord 15(3): 205-217. Copyright© Kulvinder Kochar Kaur, et al.
-
Benton D, Young HA (2016) A meta-analysis of the relationship between brain Dopamine receptors and obesity: a matter of changes in behavior rather than food addiction. Int J Obes (Lond) 40(1): 12-21.
-
Pan YQ, Qiao L, Xue XD, Fu JH (2015) Association between ANKK1 (rs 1800497) polymorphism of DRD2 gene and attention deficit hyperactivity disorder: A Meta-analysis. Neurosci Lette 590: 101- 105.
-
Comings DE, Blum K (2000) Reward deficiency syndrome: genetic aspects of behavioral disorders. Prog Brain Res 126: 325-341.
-
Blum K (1990) Allele Association of human Dopamine D2 receptor gene in Alcoholism. JAMA 263(15): 2055- 2060.
-
Yue K, Pronger AM, Ferguson AR, Temkin NR, Sharma S, et al. (2015) Association of a common genetic variant within ANKK1 with six months cognitive performance after traumatic brain injury. Neurogenetics 16(3): 169-180.
-
McGuire V, Van Den Eeden SK, Tanner CM, Kamel F, Umbach DM, et al. (2011) Association of DRD2 and DRD3 polymorphisms with Parkinson’s disease In a multiethnic consortium. J Neurol Sci 307(1-2): 22-29.
-
Noble EP, Gottschalk LA, Fallon JH, Ritchie TL, Wu JC (1997) D2 dopamine receptor polymorphism and brain regional glucose metabolism. Am J Med Genet 74(2): 162-166.
-
Cerasa A, Gioia MC, Tarantino P, Labate A, Arabia G, et al. (2009) The DRD2 Taq1A polymorphism associated with changed midbrain volumes in healthy individuals. Genes Brain and Behav 8(4): 459-463.
-
Klien TA, Neumann J, Reuter M, Hennig J, von Cramon DY, et al. (2007) Genetically determined differences in learning from errors. Science 318(5856): 1642-1645.
-
Nymberg C, Banaschewski T, Bokde AL, Büchel C, Conrod P, et al. (2014) DRD2/ ANKK1 polymorphism modulates the effect of ventral Striatal activation of working memory performance. Neuropsychopharmacology 39(10): 2357-2365.
-
Cohen MX, Young J, Baek JM, Kessler C, Ranganath C (2005) Individual differences in extraversion and Kulvinder Kochar Kaur, et al. Chronically Elevated Triglycerides as a Result of High Fat Palatable Diet Resulting in a Vicious Cycle on Reinforcing Reward and Dopamine Signaling: A Possible Cause for the Obesity Epidemic Worldwide in the Food Environment Available-A Comprehensive Review. J Endocrinol 2019, 3(2): 000143. dopamine genetics predict neural reward responses. Brain Res Cogn Brain Res 25(3): 851-861.
-
Wiener M, St John N, McD Young R, Saunders JB, Noble EP, et al. (2014) Impulsivity-related cognition in alcohol dependence: Is it moderated by DRD2/ ANKK1 gene status and executive dysfunction?. Addictive Behaviors 39(11): 1663-1669.
-
Berry hill ME, Wiener M, Stephens JA, Lohoff FW, Coslett HB (2013) COMT and ANKK1-Taq-la Genetic polymorphisms influence Visual working memory. PLoSOne 8(1): e55862.
-
Cohen MX, Krohn-Grimberghe A, Elger CE, Weber B (2007) Dopamine gene predicts the brain response to dopaminergic drug. Eur J Neurosci 26(12): 3652- 3660.
-
Persson J, Rieckmann A, Kalpouzos G, Fischer H, Bäckman L (2015) Influence of a DRD2 polymorphism on updating long-term memory representations and caudate BOLD activity: Magnification in aging. Human Brain Mapp 36(4): 1325-1334.
-
Epstein LH, Wright SM, Paluch RA, Leddy JJ, Hawk LW Jr, et al. (2004) Relation between food reinforcement and dopamine genotypes and its effects on food intake in smokers. Am J Clin Nutr 80(1): 82-88.
-
Yeh J, Trang A, Henning SM, Wilhalme H, Carpenter C, et al. (2016) Food cravings, food addiction and a dopamine resistant (SRD2A1) receptor polymorphism in Asian American College Students. Asia Pac J Clin Nutr 25(2): 424-429.
-
Stice E, Burger KS, Yokun S (2015) Reward region Responsivity Predicts Future Weight Gain and Moderating Effects of the TaqAa1 allele. J Neurosci 35(28): 10316-10324.
-
Stice E, Spoor S, Bohon C, Small DM (2008) Relation between obesity and blunted striatal response to food is moderated by TaqAa1 allele. Science 322(5900): 449-452.
-
Babbs RK, Sun X, Felsted J, Chouinard-Decorte F, Veldhuizen MG, et al. (2013) Decreased caudate response to milkshake is associated with higher body mass and greater impulsivity. Physiol Behav 121: 103-111. Copyright© Kulvinder Kochar Kaur, et al.
-
Sun X, Kroemer NB, Veldhuizen MG, Babbs AE, de Araujo IE, et al. (2015) Basolateral amygdale response to food cues in the absence of hunger is associated with weight gain susceptibility. J Neurosci 35(20): 7964-7976.
-
Kroemer NB, Small DM (2016) Fuel not fun: Reinterpreting attenuated brain response to reward in obesity. Physiol Behav 162: 37-45.
-
Leddy JJ, Epstein LH, Jaroni JL, Roemmich JN, Paluch RA, et al. (2004) Influence of methamphetamine on eating in obese men. Obes Res 12(2): 224-232.
-
Lawford BR, Young RM, Rowell JA, Qualichefski J, Fletcher BH, et al. (1995) Bromocriptine in the Treatment of the alcoholics in the D2 Dopamine Receptor A1 Allele. Nat Med 1(4): 337-341.
-
Stice E, Yokum S, Blum K, Bohon C (2010) Weight gain is associated with reduced striatal response to palatable food. J Neurosci 30(39): 13105-13109.
-
Cameron JD, Riou ME, Tesson F, Goldfield GS, Rabasa-Lhoret R, et al. (2013) The Taq1ARFLP is associated with attenuated interventional-induced body weight loss and increased carbohydrate intake in post-menopausal obese women. Appetite 60: 111- 116.
-
Beeler J, Faust RP, Turkson S, Ye H, Zhuang X (2016) Low Dopamine D2 Receptor Increases Vulnerability in obesity via Reduced Physical Activity, Not Increased Appetite Motivation. Biol Psychiatry 79(11): 887-897.
-
Zhang Y, Bertolino A, Fazio L, Blasi G, Rampino A, et al. (2007) Polymorphisms in human Dopamine D2 Receptor gene affect gene expression, splicing and neuronal activity during working memory. Proc Natl Acad Sci USA 104(51): 20552-20557.
-
Kotlyar M, Pastrello C, Pivetta F, Lo Sardo A, Cumbaa C, et al. (2015) In silico prediction physical protein interactions and characterization of interactome orphans. Nat Methods 12(1): 79-84.
-
Espana-Serrano L, Guerra Martín-Palanco N, Montero-Pedrazuela A, Pérez-Santamarina E, Vidal R, et al. (2016) The Addiction Related Protein ANKK1 is differentially expressed during the Cell Cycle in Neural Precursors. Cereb Cortex 27(5): 2809-2819. Kulvinder Kochar Kaur, et al. Chronically Elevated Triglycerides as a Result of High Fat Palatable Diet Resulting in a Vicious Cycle on Reinforcing Reward and Dopamine Signaling: A Possible Cause for the Obesity Epidemic Worldwide in the Food Environment Available-A Comprehensive Review. J Endocrinol 2019, 3(2): 000143.
-
Buckham LB, Thompson MM, Lippert RN, Blackwell TS, Yull FE, et al. (2015) Evidence for a novel functional role of astrocytes in the acute homeostatic response to high-fat diet intake in mice. Mol Metab 4(1): 58-63.
-
Choi JY, Jang EH, Park CS, Kang JH (2005) Enhanced susceptibility to 1methyl-4phenyl-12,3,6-tetra hydropyridine neurotoxicity in high fat diet induced obesity. Free Radic Biol Med 38(6): 806-816.
-
Thaler JP, Choi SJ, Schwartz MW, Wisse BE (2010) Hypothalamic inflammation and energy homeostasis: resolving the paradox. Front Neuroendocrinol 31(1): 79-84.
-
Stoeckel LE, Arvanitakis Z, Gandy S, Small D, Kahn CR, et al. (2016) “White Paper” meeting summary and catalyst for future inquiry. Complex mechanisms like neurocognitive dysfunction to insulin resistance and other metabolic dysfunction. F1000Res 5: 353.
-
Ariza M, Garolera M, Jurado MA, Garcia-Garcia I, Hernan I, et al. (2012) Dopamine genes (DRD2/ANKK1-TaqA1and DRD4-7R) and Executive Function: Their Interaction with obesity. PLoS One 7(7): e41482.
-
Burke MV, Small DM (2016) Effects of modern food environment on stiatal function, cognition and regulation of ingestive Behavior. Curr Opin Behav Sci 9: 97-105.
-
Cansell C, Luquet S (2016) Triglyceride sensing in the reward circuitry: A new insight in feeding behavior regulation. Biochemie 120: 75-80.
-
Wei NL, Quan ZF, Zhao T, Yu XD, Xie Q, et al. (2019) Chronic stress increases the susceptibility to food addiction by increasing the levels of DR2 and MOR in the nucleus accumbens. Neuropsychiatr Dis Treat 15: 1211-1229.
-
Archer E, Lavie CJ, Hill JO (2018) The Contributions of 'Diet', 'Genes', and Physical Activity to the Etiology of Obesity: Contrary Evidence and Consilience. Prog Cardiovasc Dis 61(2): 89-102.
-
Archer E, Pavela G, McDonald S, Lavie CJ, Hill JO (2018) Cell-specific ‘’Competition for Calories‘’ Drives Assymmetric Nutrient-Energy Partitioning, Obesity, and Metabolic Diseases in Human and Non-Human Animals. Front Physiol 9: 1053. Copyright© Kulvinder Kochar Kaur, et al.
-
Archer E, Lavie CJ, Hill JO (2018) The failure to Measure Dietary Intake Engendered a Fictional Dicourse on Diet-Disease Relations. Front Nutr 5: 105. Kulvinder Kochar Kaur, et al. Chronically Elevated Triglycerides as a Result of High Fat Palatable Diet Resulting in a Vicious Cycle on Reinforcing Reward and Dopamine Signaling: A Possible Cause for the Obesity Epidemic Worldwide in the Food Environment Available-A Comprehensive Review. J Endocrinol 2019, 3(2): 000143.
-
Sun X, Luquet S, Small DM (2017) DRD2: Bridging the Genome and ingestive behavior. Trends Cogn Sci 21(5): 372-384. Copyright© Kulvinder Kochar Kaur, et al.
- Shaping Healthy Futures: Pediatric Endocrine Breakthroughs of 2025
- Precision Medicine in Obesity: Customizing Treatment for 2025
- The Thyroid Revolution: How 2025 is Redefining Hormone Health
- Editorial- Targeting Immunometabolism for Generating Innovative Therapies for Cancer
- Current Knowledge of Chickenpox
- Correlation of Preinjection Values of Gonadotropins and Estradiol Level with Clinical and Radiologic Evidence of Sufficient Pubertal Suppression in Girls with Central Precocious Puberty