An Update on the Role of PI3K/ PTEN/ AKT Signaling Pathways in the Generation of Germ Cell Along with their Implications in Germ Cell Tumors Besides Ovarian Impairment: A Narrative Review
Numerous studies have pointed that the PI3K/ PTEN/ AKT signaling pathways are the key controllers of generation of germ cells precursors labeled as Primordial germ cells in addition to sustenance of follicular pool. Earlier we have already highlighted the role of MAPK/ERK along with Hippo/MST signaling for escalating of effectiveness as well as avoidance of chemo resistance in various Cancer Therapies besides modes by which chemotherapy with utilization of alkylating Agents result in follicular activation if there was any further deletion of primordial follicles? along with use of manipulation of the signaling pathways inclusive of PI3K/Akt pathway along with Hippo signaling path way in the Yes-associated protein (YAP) transcriptional coactivator with post synaptic density protein, drosophila disc large tumor suppressor [PDZ] and zonula occludens-1-binding motif (TAZ) -besides other controlling constituents in the generation of the EFs correlated with development of Early/ Primordial Follicles for the escalat d ART results in premature ovarian insufficiency (POI) & DOR patients. Here we concentrated on further updating the knowledge regarding germ cell tumors production besides reemphasizing on the dynamics of the ovarian Primordial germ cells pools besides decontrolling of these pathways In the generation of germ cell tumors along with ovarian impairment inclusive of, premature ovarian insufficiency (POI), premature ovarian failure (POF) & diminished ovarian reserve (DOR) patients. Thus here we conducted a review utilizing search engine pubmed, google scholar; web of science; embase; Cochrane review library utilizing the MeSH terms like germ cells origin; Primordial germ cells; PI3K/PTEN/AKT signaling pathways; Hippo/MST pathways; mammalian target of rapamycin complex (mTORC); eukaryotic translation initiation factor 4E binding protein 1( (4E-BP1) c; TAZ; YAP ; MST1/2. We searched from 1990’s till date in 2022 We found 150 articles of which we used 62 articles for this narrative review or updating the pathways implicated and detail these PI3K/ PTEN/AKT signaling pathways mainly from point of view of Primordial germ cells development besides germ cell tumors development.
Introduction
Primordial germ cells (PGC) represent the embryonic germ cells precursors. They are obtained from a subset of epiblast cells which migrate towards the extra embryonic area immediately prior to gastrulation, before reentering the embryo proper along with movement towards the gonadal ridges (GR’s). From them arise oocytes or prospermatogonia (the fetal precursor’s spermatogonia) in the generating fetal ovaries along with testes respectively. This type of stepwise processes takes place in all mammalian species inclusive of humans, despite the factors implicated in addition to timing might vary. However from the experimental proof collected over the last 3 decades a right activation level along with controlling of the phosphatidyl inositol 3-kinase (PI3K)/phosphatase as well as tension homolog(PTEN)/ AKT (protein kinase B) signaling pathways have been the characteristics that have been conserved regarding the PGC generation [1, 2].
Initially the major outcomes derived from numerous laboratories corroborated the belief basically regarding PGC survival, proliferation as well as migration. At a subsequent period later the formation of the follicle stockpile in the ovary along with its dynamics, the PI3K/ PTEN)/AKT signaling pathways persistently possess a key part regarding controlling the activation of the primordial follicle besides oocytes growth. Earlier we had reviewed the role of crosstalk amongst MAPK/ERK along with Hippo/MST signaling for escalating of effectiveness as well as avoidance of chemo resistance in different cancer therapies besides modes by which chemotherapy with utilization of alkylating agents result in follicular activation. We queried is there any further deletion of primordial follicles? Further did the use of manipulation of the signaling pathways inclusive of PI3K/Akt pathway along with Hippo signaling pathway -besides other controlling constituents in the generation of the EFs correlated with development of Early/ Primordial Follicles for the escalated ART results in premature ovarian insufficiency (POI) & diminished ovarian reserve (DOR) patients [3, 4, 5]. Here our major objective is to detail how the decontrolling of the PI3K/ PTEN)/AKT signaling pathways results in generation of the certain kinds of germ cell tumors. Greater details are available in ref by Makker A, et al. [4], along with that by Maidarti M, et al. [5].
PI3K/ PTEN/ AKT Signaling Pathways
Phosphatidyl inositol 3 - kinase (PI3K) represents lipid kinases possessing a central part in the controlling of different cell actions inclusive of apoptosis, autophagy, cell cycle, differentiation, besides motility. Their activation takes place downstream of tyrosine kinase receptors (RTKs) besides G- protein coupled receptor (GPCRs). 4 main extracellular signals, growth factors (GF), cytokines, hormones/ chemokines along with integrin’s possess the capacity of activation of PI3Ks. In the cytoplasm, PI3K phosphorylates the 3’ hydroxyl position of the inositol ring of phosphatidyl inositides results in formation of phosphatidylinositol 3,4,5-trisphosphate (PIP3 ), a 2nd messenger that enrolls AKT (protein kinase B) to the cell membrane by phosphorylation. This specific step is under strict regulation of PTEN, that works in the form of a negative controller of the pathways by transformation of PIP3 back to phosphatidylinositol 4,5-bisphosphate (PIP2). For total activation AKT needs extra phosphorylation by mammalian target of rapamycin inhibitors (mTORC2) subsequent to which it possesses the capacity of phosphorylating numerous substrates. Till date Identification of a minimum of 13 substrates of AKT have been found in mammalian cell [6]. The classification of these substrates is made into 2 main groups
- Apoptosis controlling
- Cell cycle controllers. Of the maximum significant pathways existent downstream of AKT is the)/ mammalian target of rapamycin inhibitors (mTOR). mTOR is a main controller of cell growth besides metabolic state in reaction to nutrients, growth factors in addition to numerous extracellular signals. mTOR congregates into 2 functionally unique complexes mTORcomplex1 (mTORC1 along with mTORC2). With the AKT signaling mTOR amongst the mTORC1 gets stimulated for directly phosphorylating the ribosomal S6 kinases, S6 K1 as well as S6 K2 besides the eukaryotic translation initiation factor (EIF4E) 4E binding protein 1 (4E-BP1 [binding proteins4 EBP1 along with 4 EBP2] with the idea of facilitation of protein generation along with finally cell survival growth as well as proliferation. mTORC1activity in this possesses the capacity of hampering cell apoptosis along with autophagy by different modes (Figure 1) [7].
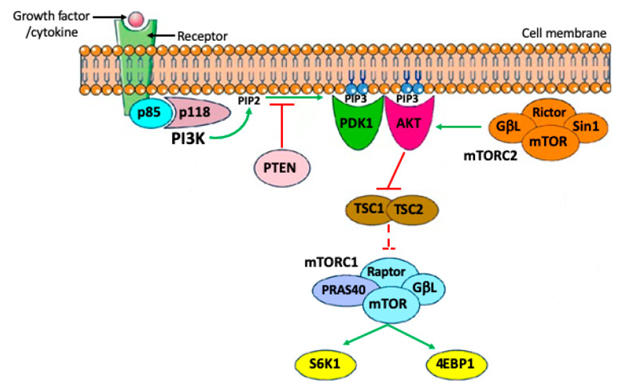
PI3K/ PTEN/ AKT Signaling Pathways Regarding PGC Generation
It has been well corroborated regarding mammalian embryos that the germ line gets determined from a small cell clusters belonging to the epiblast that is believed to form all cell lineages immediately prior to gastrulation. With the inducing action regarding, wingless -related integration site (WNT), Bone morphogenetic proteins (BMP), growth factors signaling, that mainly is dependent on the Sma as well as Mad -related protein (SMAD) besides β-catenin pathways [8], expression of transcription factor like OCT4, SOX2, 17NANOG, [PR (PRD1-BF-RIZ1 homologous) domain containing 16(PRDM16)] along with 14 which start the reprogramming of their genome towards a naive pluripotent status along with simultaneously avoidance of their differentiation into somatic cell lineages [9]. Progression of these events occurs in addition to reinforcement in the wall of the yolksac where these in particular germ cells migrate. Now a newer induction gets initiated which decide before hand these cells in the form of germ cell precursors labeled as Primordial germ cells (PGC). PGC whose destiny is to generate the gametes subsequent to colonization of the gonadal primordium amongst the proper embryo possess the properties of expression of surface markers like alkaline phosphatase (APase) as well as Stage specific embryonic antigen (SSEA), oligosaccharides characteristic of pluripotent stem cells, RNA binding proteins like TIAR, LIN28, DND1 as well as Nanos3, significant controllers of cell survival besides getting self-renewed [10]. At the time of this period PGCs further demonstrated KIT receptor for the Kit ligand ( KL) cytokines along with receptors for the growth factors like Fibroblast growth factor (FGF) or other cytokines like leukaemia inhibitory factors (LIF), the LIF receptor (LIFR) LIFR-gp180 complex [11, 12]. These receptors have been observed on the membrane of migratory as well as post migratory PGCs along with in vitro studies corroborated regarding their activation by the respective ligands, KL or stem cell factor (SCF), basic FGF (bFGF) or (FGF2) as well as LIF or associated cytokines, facilitate PGCs survival, migration along with proliferation [11, 12, 13]. Nevertheless, the downstream effectors regarding activation of these receptors are not clarified. Indeed 3 intracellular signaling pathways of which we possess the knowledge, get activated by KL LIFR- gp180,
- C-Jun-N-terminal kinase (JAK) / Signal Transducers and Activators of Transcription 3 (STAT3);
- PI3K/AKT as well a Srchomology2 (SH2) domain containing tyrosine phosphatase (SHP2) (2]/ mitogen activated protein kinase (MAPK), just this latter possibly can be activated by LIF in PGCs [14]. Regarding the3 probable signaling pathways downstream of fibroblast growth factor receptor (FGFR) being activated are RS, RAF/MAPK/PI3K/AKT besides phospholipase Cγ, the first probably can be activated by FGF in migratory, besides post migratory PGCs as well [11]. On the other hand greater knowledge is accessible regarding KL/KIT system signaling. A key part regarding PGCs survival as well as probably migration in vivo got corroborated by natural or induced mutants just for system [12, 15].
KL/ KIT in PGCs in form of Main Effectors Regarding PI3K/AKT Signaling in PGCs
KL alias KITL or Stem Cell factor (SCF) is present in the form of a membrane fixed as well as soluble isoforms which originate from alternative RNA splicing along with processing involving proteolysis [15]. It has been posited that this membrane fixed KL takes part in signaling implicating short distance signaling that gets conveyed by cell - cell contact contrasted to soluble isoforms participating in longer distance signaling which get conveyed by diffusion via the extracellular medium. Apparently both isoforms possess the capacity of activation of KIT in PGCs [16], however no knowledge regarding their capacity of activation of variable signaling pathway is existent.
KIT is a member of the type III tyrosine kinase receptor family members with the information regarding stimulation of KIT by KL result in dimerization, activation besides inherent tyrosine kinase action along with phosphorylation of crucial tyrosine residues amongst the receptor. The subsequent phosphotyrosine residues function as docking areas for molecules possessing the SH2 in addition to other phosphotyrosine binding domains. On activation KIT further catalyzes direct phosphorylation of substrate proteins. Of the main signaling pathways finally whose activation occurs by KL/ KIT are inclusive of RAS-RAF-MEK-ERK-PLCγ, Src along with PI3K/AKT, of which the latter 2 apparently are the ones possessing maximum significance regarding sustenance of PGC survival, besides participate in migration towards the gonadal primordium [17, 18]. Intriguingly Behrens GHGM, et al. [19], observed that binding of 17β estradiol (E2) to its receptor ERαfurther possessed the capacity of induction of a fast paced PI3K based AKT phosphorylation in mouse PGCs [19].
Like in other cells it is probable to posit regarding post migratory PGCs that the binding of p85 regulatory subunit of PI3K tophosphotyrosine residues Y719 of the KIT getting activated by one or both of its SH2 domains along with crosstalk amongst p85- p110 result in allosteric activation of the catalytic subunit of the enzyme [20, 21]. Once PI3K activation takes place, a series of steps result in classical AKT phosphorylation detailed earlier. Nevertheless, regarding this induced mutations interference with the binding area of Tyr 719 for the p85 regulatory subunit of PI3K in the KIT receptor [6, 14] possessed no significant impact on the generation of mouse PGCs is apparently contradictory to the thought regarding the key part of PI3K in PGCs generation. Conversely, in vitro incubation of mouse PGCs with KL caused a fast phosphorylation of AKT that gets ameliorated by PI3K hampering agents [15]. On reconciliation these seemingly contradictory outcomes, Godin, et al. [16] illustrated that in mouse PGCs, the KIT autophosphorylation action that was efficaciously stimulated by KL was correlated with phosphorylation of AKT that gets modulated by PI3K or Src kinases. Hence once phosphorylated Y719is lacking, it is feasible that p85 gets enrolled by other signaling or scaffold protein, like Grb2 or Src which ultimately result in PI3K activation [16].
In the same publications an innovative part for KL in the form of PGCs chemo attractant along with PI3K/AKT as well as Src kinases in the form of actors implicated in the activation of the PGCs machinery correlated with migration were illustrated. Additionally, corroboration regarding 2 other probable PI3K based KL/ KIT action on PGCs, adhesion to somatic cells [17] besides survival which is secondary to a decreased expression of the proapoptotic Bax gene have been revealed earlier [18]. Gu’s group [13], observed that the membrane bound KL form had a sustenance of greater local amounts of mouse PGCs motility besides deciding their own area of migration.
Regarding greater 16 downstream substrates of AKT which might possess a direct or indirect implication in PGCs migration in addition to survival are not clarified. Certain of these are pointed in Figure 2 besides detailed by de Felici & Klinger [19].
PI3K/ PTEN/ AKT Signaling Pathways Regarding Development of Germ Cells Tumors From PGCS
Germ cells tumors are neoplasms obtained from germ cells which have the capacity of possessing both immature along with mature elements that might undergo differentiation into different tissue kinds. The initiation of these tumors preferentially takes place in the gonads, nevertheless, they might be observed in the extra gonadal areas (EGCT). The observation of EGCT mainly takes place in children or young adults along with canonically originate in the midline localization with the cranium in addition to sacrococcygeal areas being the commonest regions. In case of testis GCT’s are labeled as testicular GCT’s (T GCT’s), in ovary, ovarian GCT’s (O GCT’s). Regarding histology EGCT along with TGCT’s are constituted by seminomas (SEs) alias germinomas besides non seminomatous tumors (NSTs). NSTs are inclusive of, various histological unique kinds like teratomas, embryonal carcinoma, endodermal sinus tumors (yolk sac tumors), choriocarcinoma as well as tumors of mixed histology. Non-seminomas possessing a pluripotent constituent labelled as embryonal carcinoma (EC) cells are referred to as teratocarcinomas.4 kinds of OGCT’s are existent dysgerminomas, parallel to testicular seminomas, teratomas, endodermal sinus tumors along with choriocarcinoma histologically counterparts of their equivalent testicular tumors. Here greater details are not given. See ref [20, 21] for the same, Our concentration is on the probable shared origination from PGCs.
Actually the posit is that EGCT along with the maximum part of TGCT’s as well as OGCT’s originate from PGCs with changed survival, decontrolled cell cycle besides failure of germ line estimation possibly work together. A hypothesis regarding the tumorigenic probability of extra gonadal germ cells tumors called ‘’germ cell theory’’ was initiated by Gunnar Teilum, pointing that EGCT’s arise from lost or roaming migratory PGCs [22]. Migration of PGCs takes place across the embryo towards the generational gonads where their differentiation into gametes occur needing driving hints regarding survival besides reaching their ultimate target or station. Furthermore on arrival the GR’s are ultimately determined in the form of germ cells along with proliferation action as oogonia prior to entrance to meiosis or pro spermatogonia with the destiny of going via mitotic arrest in the differentiating ovary or testis respectively.
It is feasible that the right formation of PGCs on migration from yolk sac towards the GR’s will be based on the balance amongst cytokines/growth factors signaling pathway which regulate their cell cycle along with survival besides their core transcription factors as well as RNA binding proteins which aid in sustenance of their inherent pluripotent behavior besides preservation of their germ line identity by suppression of differentiation towards somatic lineage. Actually, the latent pluripotency obtained from specification along with the significant hypo methylated status of the chromatin superimposed on PGCs at the time of migration reflects a risk regarding tumorigenic conversion.
The way revealed earlier PGCs is correlated with a wave of KL expression by the somatic cells surrounding which via KIT/PI3K/AKT pathways aid in sustenance of their survival besides promoting migration [13]. Numerous publications have documented regarding escalated activation of the PI3K/AKT pathways subsequent to ablation/hampering of PTEN or persistent AKT hyper activation postponed PGCs along their migratory pathway [17, 19]. In the existence of KL, PGCs intricate to GR’s persist with their migration besides colonizing the gonads whereas those keeping distant in the midline body structures normally demise by the Bax- based apoptotic pathway once KL is lacking [13, 20, 23]. It has been pointed that this might be a refined manner regarding avoidance of abnormal PGCs migration: PGCs that get lost from their migratory pathway get deleted via apoptosis [23]. Nevertheless, on misplacement PGCs possess the capacity of survival in extra gonadal regions in some microenvironment they might continue to be undetermined along with give origin to variable kind of tumors. Actually with time their stimulation might take place for proliferation besides sustenance of germ line specification, give origin to germinomas or subsequent to elimination of the limitation of differentiation, towards somatic lineages, form non seminomatous tumors (NSTs). Variable corroborative evidence pointed that PGCs are not totally determined till they gain entry into the GR’s where they continuously lose the pluripotency besides somatic potential along with acquisition of a definitive germ cell fate. Akin to that subsequent to gonadal colonization decontrolled PGCs survival/proliferation correlated with failure or part determination in the form of gametes is the probable etiology of the generation of TGCT’s as well as OGCT’s. Regarding this innovative experimental work performance in 129/Sv mice prone to spontaneous production of testicular teratomas have illustrated that these tumors are in fact obtained from PGCs [21], besides in these strains PGCs within fetal testis persistently proliferate at the time that lies beyond the time normally they undergo mitotic arrest [24]. Corroborating this belief variable proofs have documented later that maximum parts of TGCT’s in mouse as well as humans take origin from the precursor cells called germ cell neoplasia in situ (GCNIS) that projects the converted PGCs into prospermatogonia [25].
A sophisticated illustration regarding latent pluripotency of migratory as well as gonadal PGCs has been attempted by in vitro conversion of mouse or human PGCs in the existence of a mixture of growth factors like KL, LIF, along with bFGF [26, 27] to produce pluripotent stem cells termed embryonal germ (EG) cells possessing the capacity of generation of teratoma on transplantation in the animal.
Regarding the Identification of the initial modes implicated in conversion of PGCs in the EG cells was the decontrolled cell cycle/survival signals correlated with overactive PI3K/AKT pathways or down regulation of PTEN [15, 16, 28] with up regulation of the transcription factor, c-Myc, Kruppel-like factor4 (KLF4), along with STAT3 [29]. At this juncture both KL along with bFGF possess the chance of being implicated in the activation of the PI3K/ AKT signaling pathways in PGCs. It has been regally illustrated that in PGCs particular mice that are deficient in Pten had AKT hyper phosphorylation besides early teratomatous foci seen in the fetal testis. Within these mice PGCs like in 129/Sv strains underwent proliferation beyond the time of acquisition of routine mitotic arrest along with despite maximum of them demise by apoptosis minimum population ended up surviving besides production of testicular teratomas [16]. The alternate target of overexpression of AKT in the converted PGCs was found in mouse double minute2 homolog (MDM2) whose escalated stability along with nuclear localization resulted in repression of p53 phosphorylation needed for its activation [28] (Figure 2). Regarding any other part of PI3K/AKT signaling pathways apart from proliferation as well as survival in intrinsic preservation of PGCs pluripotent characteristics is not clarified though requires evaluation. Intriguingly, PI3K hampering in human embryonic stem cells (HESC) caused down regulation of pluripotency markers simultaneously with the up regulation of lineage particular genes that together pointed that robustly point to the elimination of pluripotency [30].
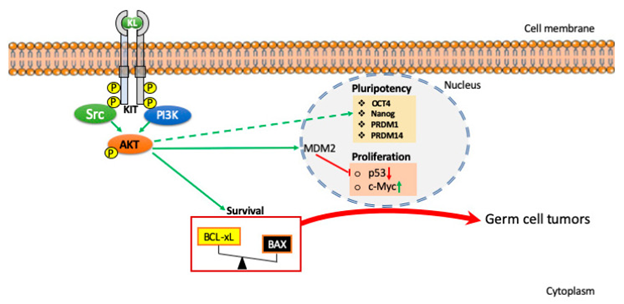
PI3K/ PTEN/ AKT Signaling Pathways Regarding Oogenesis along with Folliculogenesis
Once colonization of the gonads takes place from the female PGCs arise the oogonia which subsequent to mitotic proliferation rounds enter into meiosis 1 in the form of primary oocytes. They are surrounded by pregranulosa cells as well as propagatingly arrested at the diplotene/dictyate stage of prophase I as well as in single are surrounded in a structure known as Primordial follicle (PMF). At the time of the last time period of perinatal generation PMF cohorts get enrolled for growth getting initiated.
Subsequent to birth there is persistence of enrollment from the PMF pool till its elimination takes place at the time when infertility occurs with the female pointed to have entered reproductive senescence, or menopause in humans [24].
Despite, recently it is corroborated that the probability in mammals like in other species (spp) new follicles can be generated subsequent to birth [25]. PMF might persist in quiet stage amongst the ovary for prolonged duration that might be inclusive of numerous decades in case of humans. The elimination of this stockpile takes place via atresia or enrollment into following folliculogenesis stages. Subsequent to enrollment an event known as follicle activation ensues with follicles possessing just two fates to finish folliculogenesis by ovulation or going through atresia (alias follicle demise). Subsequent to activation, the flattened pre granulosa cells surrounding the oocytes in PMF are differentiated into cuboidal cells along with resume mitotic activation besides proliferation guiding the production of numerous granulosa cell layers in addition to transit initially into primary followed by secondary or preantral follicles. Out of the antral follicles ultimately the selection of ovulatory follicle takes place. Just the follicle possessing the capacity of enough response to gonadotrophins ([follicle stimulating hormone (FSH) as well as Luteinizing hormone(LH)]) survive besides finishing folliculogenesis with ovulation while the remainder undergo atresia.
Apparently the atresia of the growing follicles primarily get initiated in the granulosa cells followed by engulfing the oocytes granulosa cells apoptosis is in general based on gonadotrophins along with steroids. On the other hand atresia of PMF is thought to parallel or occur subsequent to oocytes demise.
The basic concept observed regarding PMF’s in adult ovary are required to get controlled by modes which ensure their dormancy besides survival along with simultaneously the preservation of the oocytes quality for longer duration. Apparently apoptosis as well as autophagy coexist in oocytes along with granulosa cells probably control oocytes survival in addition to follicle atresia at the time of folliculogenesis along with PI3K/ AKT signaling pathways for considerably important part in either events.
In the initial nineties this belief regarding PI3K/ AKT signaling pathways possessing the main control of primordial follicle of which major of oocyte in addition to follicle activation got started. Akin to PGCs that the KL/ KIT in oocytes apparently is the main upstream activator of PI3K/AKT signaling pathways despite other growth factors (GF) along with receptors like Insulin like growth factor1 (IGF1)/IGF receptor (IGF1R) possess the chances of aiding to these pathways [27]. The necessary part KL/ KIT in oocytes along with follicle generation events have been well corroborated by utilization of in vivo, in vitro strategies inclusive of naturally occurring or induced mutations [28, 29]. Information is existent regarding KIT being there in oocytes at all the stages of follicular generation in postnatal ovaries of mouse, rat along with humans [30, 31]. Conversely to KIT, KL gets generated by surrounding granulosa cells [30, 31, 32]. Contrasted to that numerous stimuli possess the capacity of controlling the PI3K/ AKT signaling pathways in granulosa cells inclusive of variable growth factors like IGF1, Epidermal growth factor (EGF)as well as physical in addition to mechanical factors (like cell adhesion, ovarian tissue protein, as well as Hippo pathways [33, 34]. Apart of EGF in the sustenance of viability (however not facilitating activation) of primordial follicles by stimulation of AKT phosphorylation has been illustrated in prepubertal domestic cats [35, 36]. Furthermore, PI3K/ AKT most probably possess an enigmatic mode that aids the oocytes in the healing DNA injury along with aid in the preservation of oocytes quality.
It is obvious from these observations that aberrations of PI3K/ AKT signaling pathways might lead to robust ovarian impairment like decreased ovarian reserve (DOR), primary ovarian insufficiency (POI premature ovarian failure (POF) besides poor ovarian reaction (POR) possessing the properties of reduction in PMF/ oocytes survival or aberrations of follicular activation .
PI3K/ PTEN/ AKT Signaling Pathways Regarding Oocytes Survival along with Apoptosis
Subsequent to evaluation of the several accessible outcomes obtained probably one can anticipate that the activation of PI3K/ AKT signaling pathways at their basal levels are significant constituents regarding the numerous factors contributing to oocytes survival amongst the PF’s. This system probably works by sustenance of antiapoptotic pathways along with probably sustenance of the basal amount of autophagy. Nevertheless, the precise AKT substrates implicated for these effects have not been documented till date. In view of the myeloid cells leukaemia 1(MCL1) along with to a lower chance B cell lymphoma- 2(Bcl2) besides B cell lymphoma extra-large (Bcl-xL), along with the proapoptotic Bcl2 correlated X protein (Bax), PUMA as well as NOXA proteins along with the initiator caspase 8 in addition to the effector caspases3 as well as 7 are the ones isolated in the form of main actors for oocytes survival besides apoptosis at this stage (see review35). AKT action might work on the expression, placement along with/or action of these proteins. Furthermore, phosphorylation of AKT could lead to stabilization of the endogenous apoptosis hampering agents, specifically X linked inhibitor of apoptosis protein (XIAP) that hampers the action of caspases 7/9 as pointed by Ene AC, et al. [36], along with Klinger FG, et al. [37]. Simultaneously lesser AKT action correlated with that of mTOR, a well-accepted AKT effector are in agreement with basal autophagy in the comparative situation of quiescent oocyte [38]. Intriguingly, Sun et al. observation was regarding miR378-3pcauses sustenance of the mouse PMF pool by enhancement of autophagy in addition to hampering apoptosis by targeting phosphatidyl inositol - dependent kinase (PDK1), believed to be a master kinase key for activation of AKT along with caspase 9 transcripts respectively [39] (Figure 3).
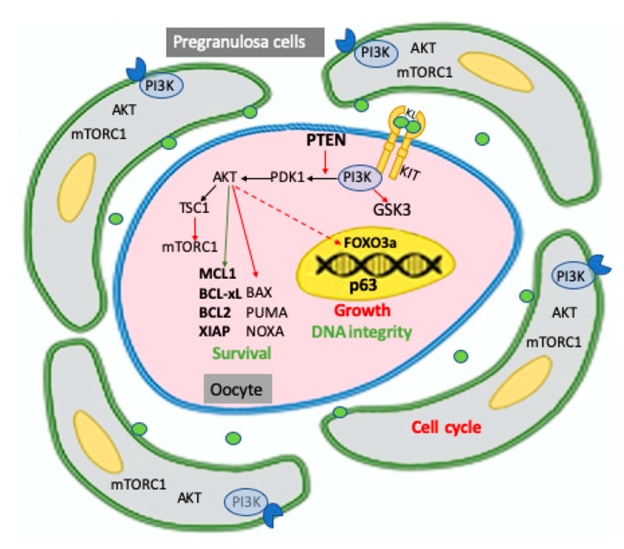
Figure 3: Courtesy ref no-7-Schematic representation of PI3K/PTEN/AKT signaling pathways identified, mainly in the mouse, in the two components of the primordial follicle, pregranulosa cells, and oocyte, and of their major targets involved in maintaining the dormancy of the follicle including cell cycle in pregranulosa cells and survival, growth, and DNA integrity in the oocyte. As extensively discussed in the test, pregranulosa cells are arrested in the reversible GO cell cycle stage while the oocyte within the primordial follicle is maintained in a quiescent status, requiring the prevalence of survival upon proapoptotic factors, preservation of DNA integrity, and the inhibition of transcription necessary to its growth. In oocytes, all or a relevant part of these activities require the basal level of PI3K/AKT activity finely regulated by the KL/KIT system and PTEN. Activation or inhibition of AKT on its substrates are represented as green and red arrows, respectively; the inhibitory action of AKT on FOXO3a is represented with a dotted arrow since it is exerted only after oocyte activation. According to a current model, not illustrated in the figure, but reported in the test, signals from the microenvironment able to activate the PI3K/AKT/ mTORC1 pathways in the pregranulosa cells reverse GO to the G1 stage and increment the production and release of factors, among these KL, which also incremented the PI3K/ AKT/mTORC1 activity in the oocyte, leading to increased protein synthesis and removing the FOXO3a inhibition on its growth; in turn, the growing oocyte releases factors such as BMP15 and GDF9, which stimulate pregranulosa cell proliferation and their maturation into granulosa cells.
PI3K/PTEN/AKT Signaling Pathways Regarding Primordial Follicle Activation along with Generation
Over the past years clarification has been attained regarding the PI3K/ AKT action downstream of KL/ KIT meant for sustenance of continuous survival of oocytes for longer time period does just partial role of PI3K / AKT signaling pathways possess significant part in the sustenance of the PMF pool amongst the mammalian ovary inclusive of humans. Actually these pathways are further implicated in activation of dormant follicles, induction of the oocytes growth besides subsequent follicular maturation [40]. Apparently the elimination of either PDK1 or PTEN that are the main controllers of the AKT action downstream of PI3K can lead to POF however via variable modes. PTEN elimination is correlated with enhanced PMF activation followed by follicular atresia, while PDK1deficiency induces exaggerated clearance of the PMF’s right from their quiescency [8, 41]. PTEN hampering in the form of a central negative controller of this pathway, whose utilization has been done broadly for the activation of PMF in variable species [5].
More recently outcomes have pointed that the action of mTOR downstream of the PI3K/ AKT pathways in granulosa cells is the main trigger regarding the PGCs activation [42]. Intriguing Andrade et al. identified in granulose besides cumulus cells from bovine forming follicles a molecular perspective or view got generated by greater PI3K/ AKT action, lesser PTEN, FOXOa, BAX amounts of miR 20a besides miR494, which lead to down regulation of PTEN amount correlated with greater amounts of oocytes generational probability [43]. This pointed that activation of the PI3K/ AKT pathways in granulosa cells aid in the acquisition of oocytes maturation at the time of follicular production. Furthermore, the activation of mTOR modulated by AKT suppression of rat granulosa cells autophagy at the time of follicular atresia [44].
Potent corroboration pointed that the mTOR is an indirect AKT effector in controlling oocytes growth along with follicular generation [45]. AKT signal via phosphorylation of tuberous sclerosis and cancer (TSC1 &2). TSC1/ TSC2 complex regarding indirect activation of the mTORC complex. A key effector of mTORC1 is 40S ribosomal protein S6 kinase (S6K) that directly controls ribosomal bio generation, cell cycle propagation, protein generation along with metabolism. The enhanced protein generation in the oocytes inclusive of liberated factors like Bone morphogenetic protein 15(BMP) 15along with Growth Differentiation factor 9(GDF9) that causes stimulation of pregranulosa cells proliferation along with sustenance of a molecular crosstalk amongst the oocytes in addition to the corresponding granulosa cells that persist at the time of the remaining duration of folliculogenesis.
Activation of follicle further needs inactivation along with elimination from the oocytes nucleus of the transcription factor FOXO3a along with the cyclin - dependent kinase inhibitors1B (CDKN1B) (alias p27kip1 orp27) of which p27 further gets inactivated in the pregranulosa cells [46]. A substrate for AKT, FOXO3a represents a transcription factor believed to lead to apoptosis, cell cycle arrest in numerous cells kinds. Actually FOXO3a gets phosphorylated by AKT gets excluded from oocytes nucleus along with is repressed from acting in the form of a transcription factor with avoidance of its growth. Considerably overexpression of constitutively active FOXO3a in the nucleus of mouse oocytes aids in preservation of their dormant status [47]. Inactivation of FOXO3a by AKT might have repercussions regarding oocytes apoptosis as well as capacity of healing DNA injury besides countering Reactive oxygen species (ROS) action that FOXO3a in general facilitates [48, 49]. More recently, regarding this it was documented about signaling bySIRT1, a NAD based deacetylase which eliminates acetyl group from different proteins inclusive of histones works via FOXO3a-MnSOD for conferring protection to mouse oocytes of follicles that are in growth phase from oxidative stress (OS) [50]. Furthermore, FOXO3a inactivation downstream of the KL/ KIT based PI3K/ AKT action partially hampered apoptosis taking place in naked rat oocytes Identified from PF [51]. Nevertheless in case besides the mode by which the actions of FOXO3a-in these oocytes get impacted amongst the dormant follicles needs clarification.
Lastly Liu L, et al. [52], documented that PI3K/ AKT action downstream of the KL/ KIT activation resulted in phosphorylation/inactivation of glycogen synthase kinase 3β (GSK3β) . GSK3β in mouse oocytes that were being cultured pointed that GSK3β might possess a part in controlling the mammalian oocytes growth along with follicular generation. Nevertheless the molecules downstream of the GSK3β in mouse oocytes require Identification.
Despite, all outcomes documented earlier robustly pointed that a central part of KL/ KIT is present regarding PF activation by utilization of a knock in mutation (Kit Y719F) which totally ameliorated PI3K signaling. John GB, et al. [53], found that KIT possessed a particular part in the sustenance of PMF reserve in the transit of primary to secondary follicles however was seen to be not essential in mice PMF activation. The significance of the mTOR/AKT signaling pathway for female fertility was nevertheless illustrated recently in women presenting with POI/POF who subsequent to receipt of AKT activator besides Hippo signaling were illustrated to attain follicular maturation, IVF of oocytes along with a healthy baby [54].
PI3K/ PTEN/ AKT Signaling Pathways Regarding Oocytes DNA Injury Reaction
At the time of stress situations the apoptosis events of oocytes amongst PMF gets modulated by a unique cell monitoring mode which implicates N terminal transactivation domain p63 (TA p63α) [55], that represents ap53 family member. A greater expression of TA p63α oocytes in the quiescence status besides TA p63α works basically for reaction towards a DNA Injury secondary to different agents inclusive of radiotherapy, chemotherapeutic substances, along with chances of ROS. Actually, TA p63α is believed to be the caretaker of the oocyte genome at the time of a long lifespan. Mouse oocytes amongst PF’s further express all essential kinases needed for triggering activation of TA p63α centered in a concomitant crosstalk of CHK2 along with CK1 [56]. PUMA in addition to NOXA undergoing transcriptional activation are the key downstream targets of oocytes apoptosis modulated by TA p63α [41]. Akin to the other 2 members of the family 53 along with p73, p63 possessing the capacity of activation of key genes implicated in healing of DNA injury like Rad51, Brca2 along with Mre1 [57], nevertheless no clarification exists regarding TA p53 in oocytes are possessing such capacity. Akin to that no clarification exists regarding AKT has the capacity of manipulating AKT. As per the concept it is probable with the information present regarding AKT possessing negative impact over regulation of p53 amounts via escalating MDM2 modulated targeting of p53 for breakdown along with on the other hand PTEN facilitates p53 translation along with protein stability [58]. Furthermore, it was documented that p63 possesses a key part in the propagation of esophageal squamous cell carcinoma cells via controlling the cell cycle through Akt signaling pathway [59].
Considerably numerous recent studies have corroborated part of AKT in manipulating DNA damage responses (DDR) along with genome stability [5, 60]. Greater AKT activity can result in repression of ATR/CHK1 signaling along with homology directed repair (HDR) through direct phosphorylation of CHK1 or TopBP1 or indirectly via hampering enrollment of double strand breaks (DSB), resection factors like RPA1, BRCA1 besides RAD51 to areas of Injury. On the other hand AKT activation takes place by DSB in a DNA-PK, or ATM/ATR based way besides in certain situations can aid in generation of radio resistance by stimulation of DNA-healing by non-homologous end joining (NHEJ). A recent publication by Rossi V, et al. [61, 62], documented that addition of KL to the cultured tissue fragments of early postnatal mouse ovaries partially conferred protection to oocytes from PMFs from apoptosis which was cisplatin stimulated however no avoidance or facilitation of the Injury as a result of drug occurred. Nevertheless, if AKT manipulates the reaction of oocytes to DNA Injury needs more evaluation.
Conclusions
PTEN/PI3K/AKT comprise a considerably significant pathways controlled signaling of numerous biological events of female gametogenesis from PGC generation to oocytes growth as well as folliculogenesis. In numerous cell kinds inclusive of PGCs, the manner emphasized here constituents of these pathways have been detailed here in the form of etiological factors for these tumors. Strong corroboration maintains the belief regarding decontrolling of the activity of these constituents in PGCs possess the capacity of changing survival proliferation, along with differentiation of PGCs with provision of significant aiding in the conversion in tumorigenic cells. Additionally, numerous results pointed that elements of these signaling pathways possess necessary part in the preservation of female fertility besides the reproductive life span. This gives a basis regarding the evaluation of probable targeted treatment for hampering of the PI3K/ AKT/ PTEN besides TSC/mTOR signaling series of steps in ovarian impairment inclusive of infertility as well as POF. Noticeably, the outcomes of these studies provide the principle for enhancement of present assisted reproductive technology (ART), is in vitro maturation of immature oocytes besides fertility preservation by in vitro activation (IVA) of follicles in cryopreserved ovarian tissue derived from patients with cancer or from women with DOR [3].
References
-
Kulvinder KK, Allahbadia GN, Singh M (2021) Escalation of efficacy & Prevention of chemoresistance in various Cancer Therapies by the utilization of targeting the crosstalk amongst MAPK/ERK along with Hippo/MST signaling-A Comprehensive Review. Acta Scientific Cancer Biology 5(9): 37-47.
-
Kulvinder KK, Allahbadia GN, Singh M (2021) The Mechanism by Which Chemotherapy with use of with Alkylating Agents Cause Follicular Activation: Is there any Further Mode for the Loss of the Primordial Follicles? A Short Communication. J Gynecol 6(3): 1-6.
-
Kulvinder KK, Allahbadia GN, Singh M (2022) Utilization of Manipulation of the Signaling Pathways Accompanying Generation of Early/ Primordial Follicles for the Enhancement of ART outcomes in POI &DOR patients- An updated Comprehensive Review In Vitro Activation with Clinical Experiences. J Med Adv Clin Case Rep 4(1): 1-13.
-
Makker A, Goel MM, Mahdi AA (2014) PI3K/PTEN /Akt and TSC/MTOR signaling pathways, ovarian dysfunction and infertility: an update. J Mol Endocrinol 53(3): R103-R118.
-
Maidarti M, Anderson RA, Telfer EE (2020) Crosstalk between PI3K/PTEN/Akt signaling and DNA damage in the oocytes: implication for Primordial follicle activation, oocytes quality and ageing. Cells 9(1): 200.
-
Jensen PB, Hunter T (2001) Oncogenic kinase signaling. Nature 411(6835): 355-363.
-
Felici MD, Klinger FG (2021) PI3K/ PTEN/ AKT signaling pathways in germ cell Development and their involvement In germ cells Tumors and ovarian dysfunction. Int J Mol Sci 22(18): 9838.
-
Chen D, Liu W, Lukianchikov A, Hancock GV, Zimmerma J, et al. (2017) Germline competency of human embryonic stem Cells depends on eomesoderm. Biol Reprod 97(6): 850-861.
-
Chen D, Liu W, Zimmerma J, Pastor WA, Kim R, et al. (2018) The TFAP2C- regulated OCT4 Naive Enhancer is involved in human germline formation. Cell Rep 25(13): 3591-3602.
-
Vertesy A, Arindranto W, Roost MS, Reinius B, Junaneda VT, et al. (2018) Parental single Cell transcriptomics reveal Epigenetic Reprogramming in human female germ cells. Nat Commun 9(1873).
-
Takeuchi Y, Molyneaux K, Runyan C, Schaible K, Wylie C (2005) The roles of FGF signaling in germ cell migration. Development 132(24): 5399-5409.
-
Todaro F, Campolo F, Barrios F, Pellegrini M, Di Cesae S, et al. (2019) Regulation of Kit expression in early mouse embryos and ES cells Development. Stem Cells 37(3): 332-344.
-
Gu Y, Runyan C, Schoemaker A, Wylie C (2009) Steel factors Primordial germ cells survival and motility from the time of their specification in the allantois and provides a continuous niche throughout their migration. Development 136(8): 1295-1303.
-
Durcovaa Hills G, Tang F, Doody G, Tooze R, Surani MA (2008) Reprogramming Primordial germ cells into pluripotent Stem Cells. PLoS ONE 3(10): e3531.
-
Settle C, Dolci S, Geremia R, Rossi P (2000) The role of Stem Cell factor and of alternative C-kit gene products in the establishment, maintenance and function of germ cells. Int JDev Biol 44(6): 599-608.
-
Godin I, Deed R, Cooke J, Zsebo K, Dwxler M, et al. (1991) Effects of the Steel gene products s on mouse Primordial germ cells in culture. Nature 352(6338): 807-809.
-
Farini D, La Scala G, Tedesco M, de Felici M (2007) Chemoattractant action and molecular signaling pathway of -kit ligand on mouse Primordial germ cells. Dev Biol 306(2): 572-383.
-
Miguel MPD, Cheng L, Holland EC, Federspiel MJ, Donovan PJ (2002) Dissection of the C-kit signaling pathway in mouse Primordial germ cells by retroviral mediated genetransfer. Proc Natl Acad Sci USA 99(16): 10458-10463.
-
Moe Behrens GHG, Klinger FG, Eskild W, Grotmol T, Haugen TB, et al. (2003) Akt/ PTEN signaling molecules estrogen dependent mouse Primordial germ cells in vitro. Mol Endocrinol 17(12): 2630-2638.
-
De Felici M ,Klinger FG, Campolo F, Balistreri CR, Barchi M, et al. (2021) To be or not To be germ cells: the extragonadal germ cells Tumors paradigm. Int J Mol Sci 22(11): 5982.
-
Dolci S, Campolo F, De Felici M (2021) Gonadal Development and germ cells tumors paradigm. Int J Mol Sci 22(11): 5982.
-
DiZazzo E, Feola A, Zuchegna C, Romano A, Donni CF, et al. (2014) Thep85 regulatory subunit of PI3Kmediates CAMP-PKA and insulin biological effects on MCF-7 Cell growth and motility. Sci World J 2014: 565839.
-
Ueda S, Mizuki M, Ikeda H, Tsujimura T, Matsumura I, et al. (2002) Critical roles of C-kit tyrosine residues 567 and 719 in Stem Cell factor -induced Chemotaxis. Contribution of Src family kinase and PI3 kinase on Calcium, mobilization and cell migration. Blood 99(9): 3342-3349.
-
Marcozzi S, Rossi V, Salustri A, De Felici M, Klinger FG (2018) Programmed cell death in the human ovary. Minerva Gi necol 70(5): 549-560.
-
Clarkson YL, McLaughlin M, Waterfall M, Dunlop CE, Skehel PA, et al. (2018) Initial characteristic of adult human ovarian cell populations isolated byDDX4 expression and aldehyde dehydrogenase activity. Sci Rep 8(1): 6953.
-
Fujihara M, Comizzoli P, Keefer CL, Wildt DE, Songsasen N (2014) Epidermal growth factor(EGF)sustains in vitro Primordial follicle viability by enhancing stromal cell proliferation via MAPK and PI3K in the prepubertal, but not adult cat ovary. Biol Reprod 90(4): 86-91.
-
Bezerra MES, Barberino RS, Menezes VG, Gouveia BB, Macedo TJS, et al. (2018) Insulin like growth factors- 1(IGF1) promotes Primordial follicle growth and reduces DNA fragmentation through the phosphatidyl inositol 3 - kinase(PI3K) / protein kinase B(PI3K /AKT) signaling pathway. Reprod Fertil Dev 30(11): 1503-1513.
-
Liu K, Rajareddy S, Liu L, Jagarlamundi K, Buman K, et al. (2006) Control of mammalian oocytes growth and early follicular Development by the oocytes PI3K kinase pathway. Dev Biol 299(1): 1-11.
-
Hutt KJ, McLaughlin M, Holland MK (2006) Kit ligand and C- Kit have diverse roles during mammalian Oogenesis along with folliculogenesis. Mol Hum Reprod 12(2): 61- 69.
-
Manova K, Huang EJ, Angeles M, de Leon V, Sanchez S, et al. (1993) The expression pattern of C- Kit ligand in gonads of mice supports role for the C- Kit receptor in oocytes growth and in proliferation of spermatogonia. Dev Biol 157(1): 85-99.
-
Motro B, Bernstein A (1993) Dynamic changes in ovarian C- Kit and Steel expression during the estrous reproductive cycle. Dev Dyn 197(1): 69-79.
-
Ismail RS, Okawara Y, Fryer JN, Vanderhyden BC (1996) Hormonal regulation of the ligand for C- Kit in the rat ovary and its effects on spontaneous oocytes meiotic maturation. Mol Reprod Dev 43(4): 458-469.
-
Kawamura K, Cheng Y, Suzuki N, Deguchi M, Sato Y, et al. (2013) Hippo signaling disruption and Akt stimulation of ovarian follicles for infertility treatment. Proc NatlAcad Sci USA 110(43): 17474-17479.
-
Cheng Y, Feng Y, Jansson L, Sato Y, Deguchi M, et al. (2015) Actin polymerization enhancing drugs mediated ovarian follicular growth by the Hippo signaling effector YAP. FASEB J 29(6): 2423-2430.
-
De Felici M, Klinger FG (2015) Programmed cell death in mouse Primordial germ cells. Int J Dev Biol 59(1-3): 41-49.
-
Ene AC, Park S, Edelmann W, Taketo T (2013) Caspase 9 is constitutively activated in mouse oocytes andplays a key role in oocytes elimination during meiotic prophase progression. Dev Biol 377(1): 213-223.
-
Klinger FG, Rossi V, De Felici M (2015) Multifaceted Programmed cell death inthe mammalianfetal ovary. Int J Dev Biol 59(1-3): 51-54.
-
De Felici M, Lobascio AM, Klinger FG (2008) cell death in fetal oocytes: many players for multiple pathways. Autophagy 4(2): 240-242.
-
Sun X, Klinger FG, Liu J, De Felici M, Shen W, et al. (2020) miR378-3p maintains the size of mouse Primordial follicle pool by regulating cell autophagy and apoptosis. Cell Death Dis 11(9): 737.
-
Hsueh AJ, Kawamura K, Cheng Y, Fauser BCJM (2015) Intraovarian control of early folliculogenesis. Endocr Rev 36(1): 1-24.
-
Kerr JB, Hutt KJ, Michalak EM, Cook MV, Enberg CJ, et al. (2012) DNA damage induced Primordial follicle oocytes apoptosis and loss of fertility require TA p63 mediated induction of Puma and Noxa. Mol Cell 48(3): 343-352.
-
Zhang M, Su YQ, Sujiura K, Xia G, Eppig JJ (2010) Granulosa cells ligand NPPC and its NPR2 maintain meioticarrest in mouse oocytes. Science 330(6002): 366-369.
-
Andrade GM, daSilveria JC, Perrini C, del Collado M, Gebremedhri S, et al. (2017) The role for PI3K/ AKT signaling pathways in the developmental competence. PLoS ONE 12(9): e185045.
-
Choi J, Jo M, Lee E, Choi D (2014) AKT is invoved in Granulosa cells autophagy regulation via mTOR signaling during rat follicular development and atresia. Reproduction 147(1): 73-80.
-
Gorre N, Adhikari D, Lindkvist R, Brannstrom M, Liu K, et al. (2014) mTORC1 signaling in oocytes is dispensable for the survival of primordial follicles and female fertility. PLoS ONE 9(10): e110491.
-
Rajareddy S, Reddy P, Du C, Jagarlamudi K, Tang W, et al. (2007) p27kip1(cyclin - dependent kinase inhibitors1B) controls ovarian development by suppressing follicle endowment and activation and promoting follicle atresia in mice. Mol Endocrinol 21(9): 2189-2202.
-
Pelosi E, Omari S, Michel M, Di ngJ, Amano T, et al. (2013) Constitutively activeFoxo3 in oocytes preserves ovarian reserve in mice. Nat Commun 4: 1843.
-
Nguyen QN, Zerafa N, Liew SH, Morgan FH, Strasser A, et al. (2018) Loss of PUMA protects the ovarian reserve during DNA damaging chemotherapy therapy and preserves fertility. Cell Death Dis 9: 618.
-
Liu H, Luo LL, Qian YS, Fu YC, Sui XX, et al. (2009) FOXO3a is involved in the apoptosis of naked oocytes of primordial follicles from neonatal rat ovaries. Biochem Biophys Res Commun 381(4): 722-727.
-
Di Emidio G, Falone S, Vitti M, DAllessandro AM, Vento M, et al. (2014) SIRT1 signaling protects mouse oocytes against Oxidative stress and is deregulated duringaging. Hum Reprod 29(9): 2006-2017.
-
Liu H, Luo LL, Qian YS, Fu YC, Sui XX, et al. (2009) FOXO3a is involved in the apoptosis of naked oocytes of primordial follicles from neonatal rat ovaries. Biochem Biophys Res Commun 381(4): 722-727.
-
Liu L, Rajareddy S, Reddy P, Jagarlamudi K, Du C, et al. (2007) phosphorylation and inactivation of glycogen synthase kinase by soluble kit ligand1 in mouse oocytes during early follicular development. J Mol Endocrinol 38(1-2): 137-146.
-
John GB, Shidler MJ, Besmer P, Castrillon DH (2009) KIT signaling via PI3K promotes ovarian follicle maturation and is dispensable for Primordial follicular activation. Dev Biol 331(2): 292-299.
-
Kawamura K, Kawamura N, Hsueh AH (2016) Activation of dormant follicles:a new treatment for premature ovarian failure? Curr Opin Obstet Gynaecol 28(3): 217- 222.
-
Lena AM, Rossi V, Osterburg S, Smirnov A, Osterburg C, et al. (2021) The P63 C terminus is essential for murine oocytes integrity. Nat Commun 12(1): 383.
-
Tuppi M, Kehrloesser S, Coutandin D, Rossi V, Luh LM, et al. (2018) DNA damage quality control requires consecutive interplay of CHK2 and CK1 to activate p63. NatStruct Mol Biol 25(3): 261-269.
-
Lin YL, Sengupta S, Gurdziel K, Bell GW, Jacks T, et al. (2009) P63 and P73 transcriptionally regulate genes involved in DNA repair. PLoS Genet 5(10): e1000680.
-
Abraham AG, ONeill E (2014) PI3K/Akt mediated regulation of P53 in Cancer. Biochem Soc Trans 42(4): 798-803.
-
Ye S, Lee KB, Park MH, Lee JS, Kim SM (2014) P63 regulates growth of esophageal squamous cell carcinoma cells via the Akt signaling pathway. Int J Oncol 44(6): 2153-2159.
-
Xu N, Lao Y, Zhang Y, Gillespie DA (2012) Akt: A double edged sword .in cell proliferation and genome stability. J Oncol 2012: 1-15.
-
Rossi V, Lispi M, Longobardi S, Mattei M, Rella FD, et al. (2017) LH prevents cisplatin induced apoptosis in oocytes and preserves female fertility in mouse. Cell Death Differ 24(1): 72-82.
-
Jagarlamui K, Liu L, Adhikari D, Reddy P, Idahl A, et al. (2009) Oocyte-specific deletion of Pten in micereveals a stage-specific function of PTEN/PI3K signaling in oocytes in controlling the follicular activation. PLoS ONE 4(7): e6186.
- Shaping Healthy Futures: Pediatric Endocrine Breakthroughs of 2025
- Precision Medicine in Obesity: Customizing Treatment for 2025
- The Thyroid Revolution: How 2025 is Redefining Hormone Health
- Editorial- Targeting Immunometabolism for Generating Innovative Therapies for Cancer
- Current Knowledge of Chickenpox
- Correlation of Preinjection Values of Gonadotropins and Estradiol Level with Clinical and Radiologic Evidence of Sufficient Pubertal Suppression in Girls with Central Precocious Puberty