Different Types of Stress as Highly Complex Endocrine and Biochemical Phenomena: A Reappraisal
The review is presented for description of physiologic and cellular stress types, together with stress phenomena, factors, reactions, mediators and outcomes. The high complexity of stress phenomena is outlined, discussing various details of HPA axis beyond its classical general scheme. Some ontogenetic and chronobiologic aspects of stress are presented, and stressful procedures of cell culturing are mentioned. It is proposed that future research efforts should be directed especially to borderline area between physiologic and cellular stress and to the comparisons of their subtypes.
Introduction
It is highly improbable that Hans Selye, a founder of stress concept, could preview all the different types of stress. His first publication about stress appeared in 1936, and Selye’s research continued till the eighties in the last century, when the great advances of biochemistry and molecular biology were already well established, since the DNA discovery in 1953. However, the elaboration of the paradigm of cellular stress occurred only in later period, close to transition from the 2nd to 3rd millennium.
Nevertheless, the hormonal investigations including catecholamines (adrenaline and noradrenaline, or epinephrine and norepinephrine) have allowed already during the first decades of 20th century to formulate the pioneering ideas about participation of these bio regulators as mediators of body reactions to adverse stimuli, primarily due to the works of Walter B. Cannon. On the other hand, since the discovery of corticosteroids at the end of forties, Hans Selye and other researchers began to describe in the next decades their contribution as mediators of stress reactions as well. Moreover, it appears that Hans Selye was the first to differentiate mineralo - and glucocorticoids (GC) [1].
Really, catecholamines and GC are considered at present as principal hormonal mediators of so called physiologic or systemic stress reactions, whereas for cellular stress such mediators include stress proteins, first of all heat shock proteins (HSP), metallothioneins and annexins [2]. However, our main goal in this mini-review is to show that as a matter of fact, stress reactions are much more complex and therefore, they surely deserve continued research efforts in near future, especially at the borderline of physiologic and cellular stress. We shall begin this essay from some definitions and concept- forming ideas.
Stress Phenomena: Involvement of Various Aspects
Figure 1 describes schematically, what are the components of stress. First of all, stress is generated during the interactions of adverse environment with live beings, and stress phenomena are the consequences of life events. In human populations these events include environmental accidents (earthquakes, floods and other extreme climatic events), military conflicts, epidemics etc. On the family level such events are the loss or taking care of the 1st degree relatives, marital conflicts and violence, divorce and others. In the interaction of environment and human or animal body it is important to differentiate stress factors (or stressors) that cause in turn stress reactions.
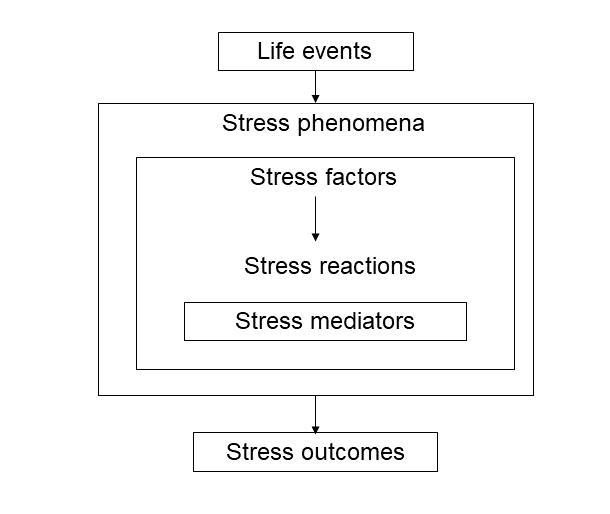
Figure1: General scheme of stress phenomena.
First of all, in humans stress factors include both real and imaginable threats associated with psychosocial stress, such as low socioeconomic position, together with financial and alimentary insecurity, as well as unstable or criminal neighborhood [3]. Stressors include also job strains, especially if an employment is related to dangerous aspects like those in police and military professions or to necessity of shift work and excessive work in general Arandelovic M, et al. [4], as for medical personal during the last pandemics of Covid-19.
On the other hand, in humans and animals stress reactions are revealed in complex physiologic responses including nervous, cardiovascular and respiratory systems, and these reactions are mediated not only by catecholamines and GC, but also by many other bioregulators, especially cytokines. Finally, stress reactions lead to stress outcomes that may include homeostatic restoration, together with hormetic enhancement, but also allostatic load and overload that result respectively in pre-disease (for example, pre- hypertension, pre diabetes) and in disorders clinically manifested, or in the extreme cases in partial or total invalidation and even death [5].
Let’s discuss now these components, as referred to cellular stress. Adverse environment for human and animal cells may include physical factors (mechanical and electrical trauma, excessively high and low temperatures, ultraviolet and other types of radiation), various chemical factors (extreme pH values, altered osmolarity, ions of heavy metals, oxidants etc.) and biological factors (for example, viral invasion).
Cellular stress reactions are directed to defense of principal cell components, first of all proteins and nucleic acids [6]. HSP are necessary for preventing protein denaturation, whereas proteolytic mechanisms are employed when protein renaturation cannot be performed. On the other hand, metallothioneins are executing essential function of neutralizing heavy metal ions like Cd2+. At least two stress proteins, heme oxygenase and thioredoxin are important for antioxidant defense, whereas polyamines (spermine, spermidine and putrescine) are chemical chaperones performing the defense of correct conformations and structure of DNA and various types of RNA [7]. Cell stress reactions have rather high energetic cost, therefore they are tightly linked to principal metabolic intracellular regulators, such as mTOR and AMPK (the last one sensitive to decrease in the ratio ATP/AMP) Reiling JH, et al [8], as well as to cell growth, both hyperplastic and hypertrophic. Now we shall discuss the complexity of various stress phenomena.
Physiologic and Cell Stress as Highly Complex Phenomena
Beginning again from physiologic stress, we should outline at first the principal role of hypothalamic-pituitary- adrenal (HPA) axis that is considered as the best studied in endocrinology till the present moment [9]. This axis is composed basically of corticotropin-releasing factor or hormone (CRF/CRH), especially in hypothalamic paraventricular nucleus, adrenocorticotropic hormone (ACTH) or simply corticotropin in anterior pituitary gland and GC in adrenal cortex.
The classical scheme of hormonal regulation includes the stimulation of ACTH release by CRF and of GC (cortisol in humans and corticosterone in rats) production by ACTH, as well as inhibitory actions of GC on CRF and ACTH secretion by means of negative feedback influence (Figure 2). However, a lot of questions still remain unanswered. At first, CRF is not the only stimulator of ACTH release. For example, in ewes arginine vasopressin (AVP) has more important corticotropin-releasing action, as compared to CRF itself, and in humans and rats AVP contributes to this activity by its greater increase in the cases of chronic stress [10].
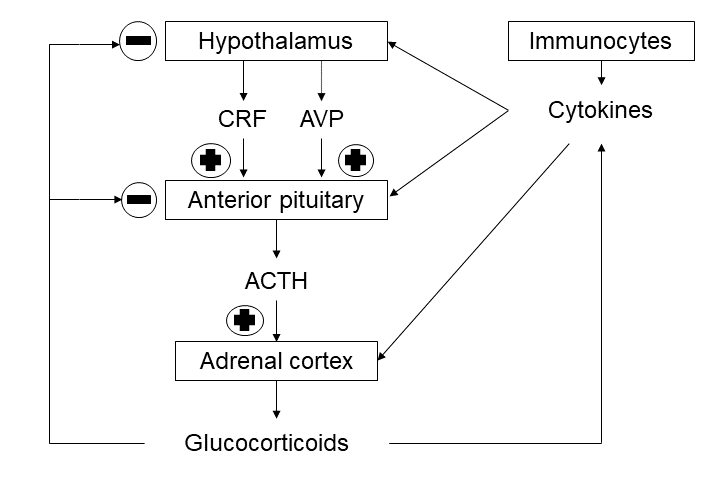
On the other hand, in adrenal cortex ACTH from pituitary gland appears to interact with CRF and ACTH locally produced by adrenal medulla [11, 12, 13, 14]. By the way, local production of central components of HPA axis is characteristic not only for adrenals, but also for the skin and immunocytes, whereas GC is produced locally by epithelial thymic cells [15]. How centrally (and peripherally) driven and locally produced hormonal components of HPA axis interact with each other, is an important issue for future research, but exactly here another complication emerges.
In fact, various bioregulators among cytokines and interleukins (IL), especially IL-1, IL-6 and tumor necrosis factor (TNF), are deeply involved in regulation of all the parts of HPA axis [16, 17, 18, 19, 20]. Therefore, at present IL-1, for example, may be considered as intrinsic its component [21]. In addition, leptin interacts with both GC and pro-inflammatory cytokines, probably contributing to their catabolic effects [22, 23].
Unfortunately, we cannot discuss here all the hormones, biogenic amines, neuropeptides and eicosanoids like prostaglandins involved in the control of HPA activity. However, we must mention an important peculiarity of its regulation, namely the difference for HPA functions in acute and chronic stress, since in the last one the levels of circulating GC may decrease, so it is not easy to understand, how chronic stress can result in adverse consequences.
First of all, both excess and deficit of GC can be detrimental, because of higher secretion of pro-inflammatory cytokines in the last case. On the other hand, not only the levels of GC are important, but also cumulative exposure (GC concentration multiplied by the time of action) that obviously may be higher in chronic stress. Nevertheless, the processes of down-regulation or desensitization of GC receptors (GR) are poorly understood, especially as related to their molecular mechanisms.
In addition, the existence of two types of corticosteroid receptors, of mineralocorticoids (MR) and GR, complicates the discussion of GC actions even more, since natural GC are able to bind to both receptor types, and the balance between such binding (MR/GR) may be quite important for regulation of HPA function [24].
On the other hand, there exist two enzymes, types 1 and 2 of 11beta-hydroxysteroid dehydrogenase (11b-HSD) that are responsible for pre-receptor metabolism of natural and some synthetic GC, with their inter conversion between the active (like cortisol) and inactive (cortisone) forms [25]. But only at present this topic becomes the area of intensive research that will probably result in the creation of novel and very interesting drugs in near future.
What for cellular stress, only just recently it became clear that intracellular HSP may be released, and their extracellular counterparts can interact with various cytokines. Moreover, it is admitted that interactions of more than 100 stress proteins with more than 100 cytokines are hardly to be studied and described [26]. Nevertheless, considering that stress proteins are evolutionarily ancient, being present already in prokaryotes and unicellular eukaryotes, whereas GC are characteristic only for vertebrate species, it is clear that cellular stress has a priority over physiologic one.
However, another components of HPA axis (CRF, ACTH) or similar bioregulators are present already in at least some invertebrate species Stefano GB, et al. [27]; Di Giannantonio M, et al. [28], therefore their role in stress phenomena should be studied further. The next item of our journey among the stress phenomena will be ontogenetic one.
Stress Factors and Reactions in Ontogeny
For a long time during the last century our understanding of this topic was quite rudimentary. The situation began to change drastically with the advent of the concept of developmental origins of health and disease (DOHaD) at the transition from the 2nd to 3rd millennium. The rare combination of epidemiologic data with results obtained on experimental models of laboratory animals has led to emergency of ideas about the participation of GC as mediators of biological programming/imprinting and embedding phenomena in the ontopathogeny of chronic non-infectious diseases.
In a rather characteristic mode, these phenomena occur when stress factors and similar processes (malnutrition, infections) or exogenous GC in excess cause intrauterine growth restriction (IUGR) or somatic growth retardation in postnatal period, especially if combined with posterior compensatory, catch-up growth processes. At present the estimations are alarming ones, as follows:
- IUGR can occur in at least 8% and up to 35% of human gestations, depending on the country, and in many cases acute or chronic stress phenomena may be involved [29, 30].
- At least one third of antenatal GC use may occur without proper necessity for such treatment, since in these cases the prematurity is mild or even absent [31].
- More than a half of all children world-wide are exposed to different types of stress [32].
- What for exogenous GC use, especially early postnatal treatment with them is criticized, since it may provoke life-long consequences, probably related, unfortunately, to permanent alterations in various organs, including the lungs and kidneys, heart and liver and particularly the brain.
We became involved in these studies already 30 years ago, when higher sensitivity to GC of macromolecular biosynthesis in cultured pituitary cells of neonatal rats was shown, as compared to more mature animals [33]. In subsequent investigations we confirmed the capacity of neonatally administered dexamethasone, one of the most potent synthetic GC, to provoke irreversible (or only partially reversible) growth retardation of rats in vivo [34].
Our recent multiple evaluations have clearly demonstrated the involvement of exo- and endogenous GC in pathogenic mechanisms of various age-related disorders, especially cardiometabolic and neuropsychiatric ones Goudochnikov VI, et al. [35], what resulted in the inclusion of stress and GC to central positions in the ontopathogenic model.
Although the progress in this area, as a whole, is quite substantial, nevertheless there are still important lacunes. First of all, the role of stress and GC in periconception and early embryonal periods was not well studied yet. However, on sheep it has been clearly shown already that short 2-day dexamethasone treatment in early gestation is sufficient to cause the biological programming of postnatal hypertension [36, 37]. Importantly enough, the period of GC treatment in this case corresponds to 8-week pregnancy in humans, when many women even don’t know yet that they are pregnant and therefore, they can use exogenous GC inadvertently [38].
Although it is clear enough that in early embryonal period hormonal mechanisms are not mature yet [39], nevertheless the possibility remains of non-direct GC influence via placenta and its hormones [40, 41]. In this regard, the experiments on cultures of human induced pluripotent stem cells demonstrated clearly the effects of GC and other hormones on this cell type [42]. These studies open very interesting perspectives for understanding the hormonal effects in early gestation.
What for the role of stress and GC in aging, there exist also many nice possibilities to explore, particularly as referred to programming/imprinting phenomena determining long- term, sometimes life-long consequences, as interconnection between development and aging. However, we cannot forget that interspecies differences are particularly large for the brain and neuro behavior, since only some animals like primates have the central nervous system relatively close to humans, and even in these cases the similarity is not great, therefore psychometric tests should be performed by psychologists in human volunteers exclusively. In this regard, another (and final) topic to discuss will be chronobiologic one, where the specialists in psychology are also engaged.
Conclusion
In addition to physiologic and cellular, acute and chronic stress, there are many other types and subtypes, for example, immunologic and inflammatory, surgical and occupational, oxidative and replicative ones. In this regard, cell culturing procedures also represent various stressor types that are not always considered by cell biology researchers [46]. Moreover, quite scarce publications affirm the maintenance of at least some biorhythms in cell cultures that can be influenced by rather stressing routine techniques, such as subculturing, medium change etc.
The mini-review presented here does not pretend to be comprehensive one. In contrast, it could provide some new ideas for those who are involved in only one paradigm, even if it is a multidisciplinary one, like DOHaD. The high complexity of stress phenomena is surely challenging, but we hope that comparisons between stress types can help in confronting this problem, really difficult to resolve.
References
-
Selye H (1955) Stress and disease. Science 122(3171): 625-631.
-
Goudochnikov VI (2015) Role of stress proteins and hormones in bioregulation of ontogeny. Probl Endokrinol 61(4): 49-53.
-
Taylor SE, Stanton AL (2007) Coping resources, coping processes, and mental health. Annu Rev Clin Psychol 3: 377-401.
-
Arandelovic M, Ilic I (2006) Stress in workplace - Possible prevention. Facta Universitatis 13(3): 139-144.
-
Goudochnikov VI (2018) Hormesis against allostasis: What is more important for interaction of aging and stress? Moip Reports: Section of Gerontology 65: 129- 136.
-
Matsumura F, Vogel CFA (2006) Evidence supporting the hypothesis that one of the main functions of the aryl hydrocarbon receptor is mediation of cell stress responses. Biol Chem 387(9): 1189-1194.
-
Rhee HJ, Kim EJ, Lee JK (2007) Physiological polyamines: Simple primordial stress molecules. J Cell Mol Med 11(4): 685-703.
-
Reiling JH, Sabatini DM (2006) Stress and mTORture signaling. Oncogene 25(48): 6373-6383.
-
Dokoumetzidis A, Iliadis A, Macheras P (2002) Nonlinear dynamics in clinical pharmacology: The paradigm of cortisol secretion and suppression. Br J Clin Pharmacol 54(1): 21-29.
-
Aguilera G (1994) Regulation of pituitary ACTH secretion during chronic stress. Front Neuroendocrinol 15(4): 321-350.
-
Hagan PM, Poole S, Bristow AF (1993) Corticotrophin- releasing factor as a mediator of the acute phase response in rats, mice and rabbits. J Endocrinol 136(2): 207-216.
-
Andreis P, Malendowics LK, Belloni AS, Nussdorfer GG (1995) Effects of pituitary adenylate cyclase activating peptide (PACAP) on the rat adrenal secretory activity: Preliminary in-vitro studies. Life Sci 56(20: 135-142.
-
Licinio J, Wong ML, Gold PW (1996) The hypothalamic- pituitary-adrenal axis in anorexia nervosa. Psychiat Res 62(1): 75-83.
-
Bornstein SR, Chrousos GP (1999) Clinical review 104: Adrenocorticotropin (ACTH)- and non-ACTH-mediated regulation of the adrenal cortex: Neural and immune imputs. J Clin Endocrinol. Metab 84(5): 1729-1736.
-
Vacchio MS, Papadopoulos V, Ashwell JD (1994) Steroid production in the thymus: Implications for thymocyte selection. J Exp Med 179(6): 1835-1846.
-
Rothwell NJ (1991) The endocrine significance of cytokines. J Endocrinol 128(2): 171-173.
-
Kennedy RL, Jones TH (1991) Cytokines in endocrinology: Their roles in health and disease. J Endocrinol 129(2): 167-178.
-
Jones TH (1994) Interleukin-6 as endocrine cytokine. Clin Endocrinol 40(6): 703-713.
-
Chesnokova V, Melmed S (2002) Minireview: Neuro- immune-endocrine modulation of the hypothalamic- pituitary-adrenal (HPA) axis by gp130 signaling molecules. Endocrinology 143(5): 15871-1574.
-
Michalska AG, Tadeusz J, Rachwalska P, Bugajski J (2013) Cytokines, prostaglandins and nitric oxide in the regulation of stress-response systems. Pharmacol Rep 65(6): 1655-1662.
-
Goudochnikov VI (2017) The contribution of stress and its mediators to the ontopathogeny of infections: A focus on the interactions of cytokines and glucocorticoids. Adv Gerontol 7: 25-28.
-
Goudochnikov VI (2003) Possible contribution of leptin, in combination with other hormones, for catabolic influence of glucocorticoids. Rev Contexto Saude 3: 9-15.
-
Goudochnikov VI (2022) Glucocorticoid-induced growth inhibition: An update. OA J Endocrinol 6(1): 1-5.
-
De Kloet ER (2003) Hormones, brain and stress. Endocrine Regul 37(2): 51-68.
-
Nobel S, Abrahmsen L, Oppermann U (2001) Metabolic conversion as a pre-receptor control mechanism for lipophilic hormones. Eur J Biochem 268(15): 4113-4125.
-
Henderson B, Kaiser F (2013) Do reciprocal interactions between cell stress proteins and cytokines create a new intra- / extra-cellular signaling nexus? Cell Stress Chaperones 18(6): 685-701.
-
Stefano GB, Smith EM (1996) Adrenocorticotropin-a central trigger in immune responsiveness: Tonal inhibition of immune activation. Med Hypotheses 46(5): 471-478.
-
Di Giannantonio M, Frydas S, Kempuraj D, Karagouni E, Hatzistilianou M, et al. (2005) Cytokines in stress. Int J Immunopathol Pharmacol 18(1): 1-5.
-
Hoet JJ, Hanson MA (1999) Intrauterine nutrition: Its importance during critical periods for cardiovascular and endocrine development J Physiol 514(pt 3): 617- 627.
-
Thapar A, Harold G, Rice F, XiaoJia G, Boivin J, et al. (2007) Do intrauterine or genetic influences explain the foetal origins of chronic disease? A novel experimental method for disentangling effects. BMC Med Res Methodol 7: 25.
-
Whittle WL, Patel FA, Alfaidy N, Holloway AC, Fraser M, et al. (2001) Glucocorticoid regulation of human and ovine parturition: The relationship between fetal hypothalamic-pituitary-adrenal axis activation and intrauterine prostaglandin production. Biol Reprod 64(4): 1019-1032.
-
Fenoglio K, Brunson KL, Baram TZ (2006) Hippocampal neuroplasticity induced by early-life stress: Functional and molecular aspects. Front Neuroendocrinol 27(2): 180-192.
-
Gudoshnikov VI, Fedotov VP (1993) The heightened sensitivity of hypophyseal cells of neonatal rats to corticosteroids. Neurosci Behav Physiol 23(2): 107-111.
-
Goudochnikov VI (1997) Pathogenesis of glucocorticoid- induced growth retardation, evaluated in experimental studies using laboratory animals and cell cultures as the models. NewsLab (22): 90-100
-
Goudochnikov VI, Santos Goudochnikov NV (2022) The role of stress and its hormonal mediators in the ontopathogeny of cardiovascular disorders. Medpress Cardiol Vasc Med 2: 202204001.
-
Dodic M, Peers A, Coghlan JP, Wintour M (1999) Can excess glucocorticoid in utero predispose to cardiovascular and metabolic disease in middle age? Trends Endocrinol Metab 10(3): 86-91.
-
Dodic M, Samuel C, Moritz K, Wintour EM, Morgan J, et al. (2001) Impaired cardiac functional reserve and left ventricular hypertrophy in adult sheep after prenatal dexamethasone exposure. Circ Res 89(7): 623-629.
-
Wintour EM, Johnson K, Koukoulas I (2003) Programming the cardiovascular system, kidney and the brain -A review. Placenta 24: 65-71.
-
Gluckman PD, Hanson MA, Cooper C, Thornburg KL (2008) Effect of in utero and early-life conditions on adult health and disease. N Engl J Med 359(1): 61-73.
-
Hewitt DP, Mark PJ, Waddell BJ (2006) Glucocorticoids prevent the normal increase in placental vascular endothelial growth factor expression and placental vascularity during late pregnancy in the rat. Endocrinology 147(12): 5568-5574.
-
Kunz LH, King JC (2007) Impact of maternal nutrition and metabolism on health of the offspring. Semin Fetal Neonat Med 12(1): 71-77.
-
Choi SA, An JH, Lee SH, Lee GH, Yang HJ, et al. (2019) Comparative evaluation of hormones and hormone-like molecules in lineage specification of human induced pluripotent stem cells. Int J Stem Cells 12(2): 240-250.
-
Charmandari E, Chrousos GP, Lambrou GI, Pavlaki A, Koide H, et al. (2011) Peripheral CLOCK regulates target- tissue glucocorticoid receptor translational activity in a circadian fashion in men. PLoS ONE 6(9): e25612.
-
Nader N, Chrousos GP, Kino T (2010) Interactions of the circadian CLOCK system and the HPA axis. Trends Endocrinol Metab 21(5): 277-286.
-
Cagampang FR, Poore KR, Hanson MA (2011) Developmental origins of the metabolic syndrome: Body clocks and stress responses. Brain Behav Immun 25(2): 214-220.
-
Wright WE, Shay JW (2002) Historical claims and current interpretations of replicative aging. Nat Biotechnol 20(7): 682-688.
- Shaping Healthy Futures: Pediatric Endocrine Breakthroughs of 2025
- Precision Medicine in Obesity: Customizing Treatment for 2025
- The Thyroid Revolution: How 2025 is Redefining Hormone Health
- Editorial- Targeting Immunometabolism for Generating Innovative Therapies for Cancer
- Current Knowledge of Chickenpox
- Correlation of Preinjection Values of Gonadotropins and Estradiol Level with Clinical and Radiologic Evidence of Sufficient Pubertal Suppression in Girls with Central Precocious Puberty