Cognitive Map is Earned with Experience, Effort and Physical Activity: Possible Disfigurement of Memory with Early GPS Use
The systematic use of the Global Positioning system (GPS) as a recursive and repetitive use of feedback about our position and destination provided by the GPS has a negative impact on human intrinsics natural navigation strategies. This negative impact is specially present when the GPS absorbs all the possible exterior stimulation that the driver might receive (visual and auditory stimulation). The auditory cortex is generally not so relevant to build an inner cognitive map, while with the GPS the audition would be the principal source of input. With GPS instructions, attention may never flow beyond the closer sensations (thigmotaxis) to the formation of a cognitive map after brain processing. In this paper we have checked brain processing of elaborating an escape route in a Morris Maze test for different aged mice (1 month vs 4 months). The younger mice were lost at the beginning of the first trial but quickly learned how to escape and were able to better remember the platform position through different trials, compared to the adult group. These new skills were more remarkable in a second session, made thirteen days later than the first one. Probably because their hippocampus and brain network made new connections, which were translated into their more adaptive behavior, so the water environment would have become a more familiar place and an essential cognitive map had arisen from their swimming experiences. This “ability to understand” or cognitive map happened because of their healthy state (one ill mouse could not survive the test). The learning was motivated by biological survival and earned by their hard physical activity during swimming to stay afloat and breathe.
Abbreviations
GPS: Global Positioning system; IACUC: American Institutional Animal Care & Use Committee; SOP: Standard Operating Procedures.
Introduction
The Global Positioning System (GPS) project was started by the United States Department of Defense in 1973 and the full constellation of 24 satellites became operational in
1993. It provides geolocation and time information to a GPS receiver anywhere on or near the Earth where there is an unobstructed line of sight to four or more GPS satellites. That is useful information of positioning, time and movement to many different people and functions, such as civil, athletes (runners), military or commercial users around the world because of its very limited (1-6%) margin of error.
However, recent studies have proved the systematic use of GPS as a recursive and repetitive use of feedback about our position and destination provided by the GPS has a negative impact on human intrinsics natural navigation strategies [1, 2]. This seems to happen because with the use of GPS as a guide to drive, the stimuli attended by the driver are more related to the stimulus-response learning, that is more dependent on the caudate nucleus of the brain, such as navigating on “auto-pilot”, while the non GPS style of driving is related to the external stimuli, the proprioception, sense of movement and the cognitive map created, which are more related to the hippocampus and motor areas in the brain. Using the GPS, the surrounding stimuli are not receiving attention, which are fully dedicated to the GPS instructions, then the proper human cognitive abilities to create a mapping strategy are defeated because of an attention deficit [1]. This negative impact is specially accused when the GPS absorbs all the possible exterior stimulation than the driver might receive (visual and auditory stimulation) because it occupies both channels of information that the driver would need to understand the outside map to interact with the milieu [2]. However, in some places of the world, the classical methods for orientation are still essential (lighthouses): for navigation there are not many satellite systems as failsafe as in the UK and the Irland [3].
That greater GPS habit of use might cause an impairment on hippocampus connections for navigation and this would be of special and relevant importance in those stages of life when the brain structure is getting formed. Our mammalian brain would have an “inner GPS” which is pointing out a very important and necessary cognitive function for our success in movements and survival in changing environments. It is well accepted by the Scientific community that this mental representation or cognitive ability can be investigated rigorously by recording single units of activity of behaving animals that use their brains and external clues to orient, such as their behavior in the Morris water maze [4]. This cognitive function is of a very high importance in some species which live in uncertain domains, for instance the marine ones, such as harbor seals, whales or codfishes or Phoca vitulina seals need to build strong cognitive maps to find and remember, based on their previous experiences, where their food might be [5, 6, 7]. Blue whales depend on their long term memory to define their migration routes and migratory movements with the purpose of maximizing the efficiency of their choices in time and water space [6]. The spatial memory is thought to be a hippocampal dependent event but also involving other structures in the brain such as motor and visual cortex and neocortical regions to organize like prefrontal and anterior cingulate cortex [8]. The auditory cortex would not be so relevant to build an inner cognitive map, while with the GPS it would be the principal source of inputs. The amygdala, on the other hand, might be like the “lighthouse” of the brain, which is constantly monitoring and regulating the environment for stimuli to signal a threat to the organism [9].
We have checked, with a Morris maze test and using domestic rodents, how the hippocampus is better structured and organized with time, experience and age. Comparing junior and adult- senior animals (few “human months’ of differences). After training they learned different strategies of approaching the platform in the Morris Maze, according to their search swimming strategies, from less to more focus on the goal of platform [8], thigmotaxis, random, scanning, chaining, focal, directed and direct: Thigmotaxis defines the behavior of the animal which swims in reaction to the feelings of the water but with not a clear goal. The Random style defines the haphazard movements of the animal in the water without conscious choice. The next manner of behavior in the water is the Scanning strategy that refers to the recursive and repetitive circles that an animal does in the same area and Chaining performance is the same as Scanning but happening in several different areas. Focal performance for swimming happens when the animal’s behavior is clearly aimed to escape from there, but with no defined clear direction. However, Directed and Directed behaviors occur when the platform is the main goal where the animal straights all the movements toward. That means, the attention of the animal flows from the direct physical sensations on their bodies (water, unstable and uninhabitable medium) to understand the maze (cognition) for defining and finding the platform (behavior to escape).
Method
Two domestic mice rodents Mus musculus (one male, one female) were obtained by acquisition in an official pet store. They mated on February the 14th and subsequent offspring of that couple were used for these experiments. Mice of pet store Mus musculus have an unknown genetic background, but they are usually sold to become pets or to be food for captive reptiles, when they are certainly good mammalian specimens for obtaining scientific behavioral and experimental data to provide information for the Scientific community. They are known as home Mus musculus or domestic Mus Musculus.A total of 12 domestic Mus musculus, an adult group (N = 6) of 4 months old (born 16th March) and a younger group (N = 6) of 1 month old (born 9th and 12th June) were tested with a Morris Maze test. We also included, during the first session of 6 trials as well, one (N = 1) wild mouse Sylvester apodemus sylvaticus, in appropriate hygienic conditions, to have a simple observational comparison between nature vs domestic performances.
For proper cognitive and motor stimulation (proprioception) domestic Mus musculus were housed with interconnected cardboard cylinders, rotating wheels (vertical and diagonal), creative colorful pipe tube tunnels, made with plastic or wood, with directions in their connections, outside and inside spacious nests, several height levels to roam around in each cage, elastic or fixed aerial horizontal ladders made from popsicle sticks and vertical ladder made from bamboo sticks (Figure 1). All those stimuli have been changed and renewed once a week to create an updated and fresh environment to stimulate their roaming entropy and exploratory behavior. Because it has been probed that an enriched environment with novelties ias very important variable for cognitive learning processes, plasticity, brain neurogenesis, reducing anxiety levels and inducing individuality in behavior [10]. All mice were group housed per ages and sexes at 25◦C, under 12 h day/night cycle, (Apodemus sylvaticus separately) in comfortable beds of finely ground Spanish pine sawdust, hay, straw and rodent litter changed at least once every week. All the cages were cleaned, changed and dewormed using antiparasitic spray at least once every week.
All experiments were approved by local authorities and were in accordance with the guidelines for animal experiments of the EU (Directive 2010/63/EU). We also strictly follow the general protocols established for mice by the American Institutional Animal Care & Use Committee (IACUC) concerning the Policy on Investigation Noncompliance and Animal Welfare and the Environmental Enrichment for Animals Standard Operating Procedures (SOP).

Figure 1: Different pictures showing the stabling form (A) and some examples of toys and motor stimulation for the cognitive and motor development of the rodents: different wheels, size and shape of cages (vertical or horizontal), internal and external colorful pipes, little hollow tree trunk with holes, which were changed and cleaned once every week. In picture (B) the Morris Maze, with the dark platform and the visual clues on the walls are visible.
The food provided was carefully chosen in a healthy variety following veterinary instructions for little rodent necessities: natural seeds without additives, nuts (pumpkin seeds, black sunflower seeds, walnuts, peanuts in shell, oatmeals, etc.), pasta (raw macaroni), toasted bread crumbs or bread peaks, fresh and dry proteins (healthy live tenebrio molitor coleoptera, unpoisoned cockroaches, boiled eggs and dry cat food), fresh lettuce, mineral stone for rodents and clean fresh water ad libitum.
Experimental Design: Morris Water Maze Task
In brief, a circular water tank (color: black, height: 31 cm, diameter: 53 cm, and 167 cm of perimeter) was located
35 cm away from the floor. The color of the water tank was transparent and the color of the plexiglass-platform was black as well, no additional color was added into the water. This circular platform (height: 11,5 cm, and diameter: 8,5 cm) was fixed at a particular location during each trial and it changed every time. Additionally, the platform was submerged to a depth of 0,5-1 cm above the water surface, but always depending on the size of the animal´s paws (water above the platform has to be the same to half the length of the rodent lower extremities) (Figure 1B). The recorder was done from a fixed place, located in the distance, providing a good view of the animal behavior, but with one blind spot, that was a traffic area. The recorder was stopped as soon as a mouse sat on a hidden platform or if the mouse swam for a maximum of 60 s per trial without finding the platform. Mice had to learn to find a hidden platform using distal spatial cues because it was assumed they were able to distinguish the clues in the surrounding environment by seeing and all of them were able to swim acceptably, except those that had some health troubles [11, 12].
Two groups of male mice of different ages (N = 12) were compared. Three dependent variables were registered: 1) latency in water till finding the platform (seconds), 2) track length of the swimming (centimeters) and 3) memory of the last positioning of platform (yes or no). The memory was recorded from the second trial following the direction of the animal’s swimming: whether or not, before finding the platform, they went in the direction and surpassed the place where the platform was in the previous trial (yes) or if they went anywhere else (no). Both groups of ages and Apodemus Sylvaticus swam in a first session of 6 trials, in consecutive days, at the same time (around 7pm, when sunsets and their biorhythms start to be more active). The two groups of ages without the Apodemus also made a second session, 13 days later than the first one. Only healthy rodents were able to survive to this deal. This testing environment of water reduces odor trial interferences [11, 12]. Clean water was changed every two trials.
Statistical analysis, graphs, tables and figures were made with IBM SPSS Statistic version 29.0.2.0, with JMP Statistical Discovery [Trial 18.0.1 (766619)] and with PowerPoint for Windows. For inter-group comparisons the unilateral analysis of variance, with non parametric Kruskal-Wallis and U-Mann Whitney were performed. Intra groups comparisons were performed with Chi-Square of Pearson Correlation (X2).
Results
It was noticeable how the younger ones started with no clue on how to solve the indicative problem and stressful situation, but learned quite fast to find an escape, till they matched their adult group in their latency and distance. Also they, at a certain point of the experiment, were even better than the group of adults. The same animals of both groups senior and junior were checked in the second session, with a time of 13 days in between and they all have improved their swimming movements in efficiency. They were also able to remember better the platform position through different trials. These new skills were more remarkable in session two, when the water environment would have become a more familiar place and an essential cognitive map had arisen from their swimming experiences, motivated by biological survival and earned by their hard physical activity during swimming to stay afloat and breathe.
Mice style of swimming in session 1 for both groups was from thigmotaxis to direct, with all its intermediate expressions (random, scanning, chaining, focal or directed, and direct) (Figures 2 & 3).
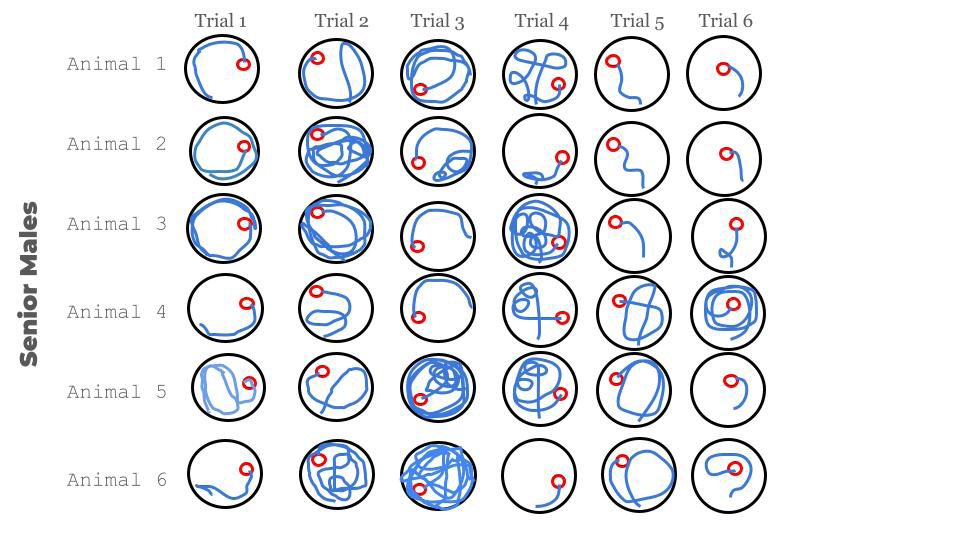
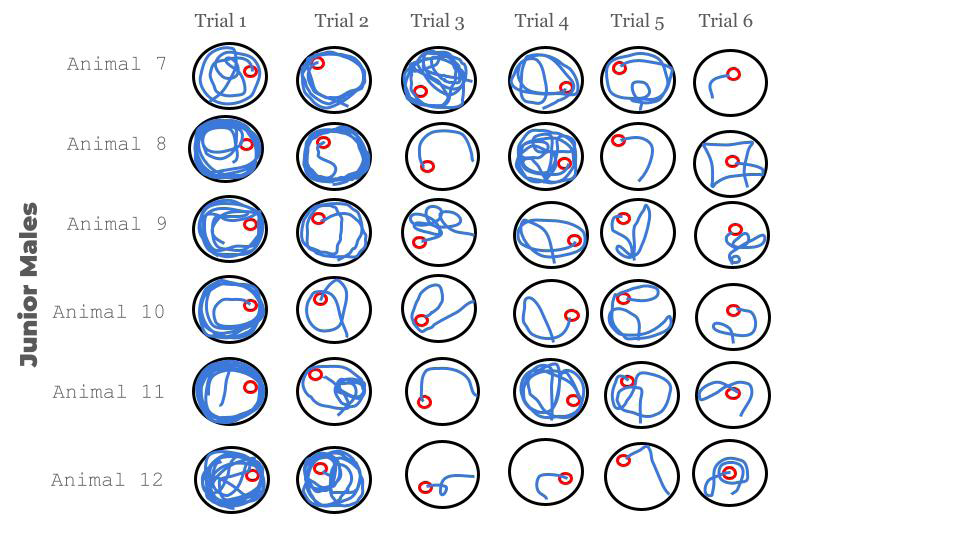
Figure 3: Different paths of the junior mice during the first session of trials. It is noticeable how chaotic the first trials were. It shows how in the beginning they were quite clumsy and scared, visible for their anxious movements in the water. But little by little their hippocampus and brain may be getting more and more prepared to understand and cope with the challenging situation through consecutive trials.
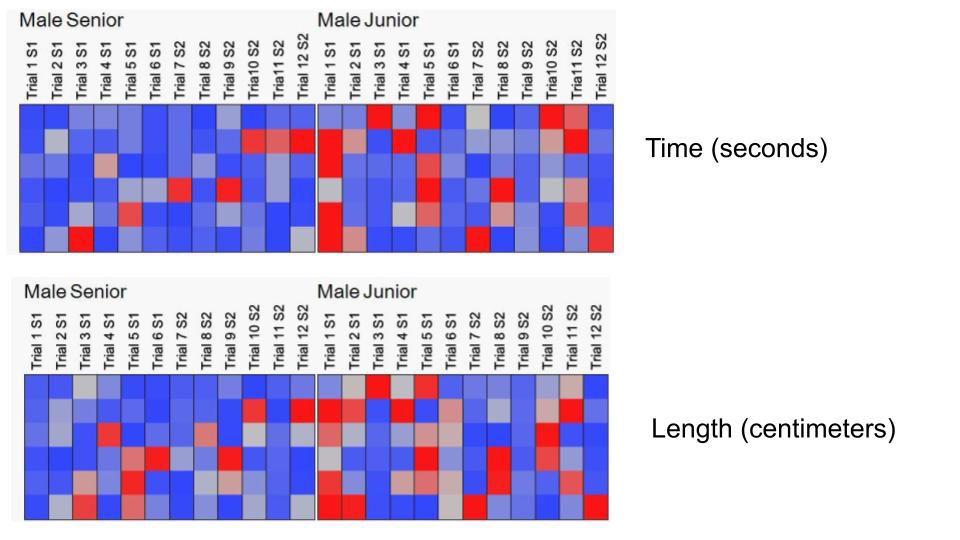
Comparing senior vs junior performances, we found clear statistical group differences in the first session for the first trial distance [F (1,13) =8.308, p = 0.004]) and first trial time [F (1,13) =8.609, p = 0.003]. In the second trial (distance [F (1,13) = 3.692, p = 0.05]), having the junior group the longer marks (Figures 4 & 5).
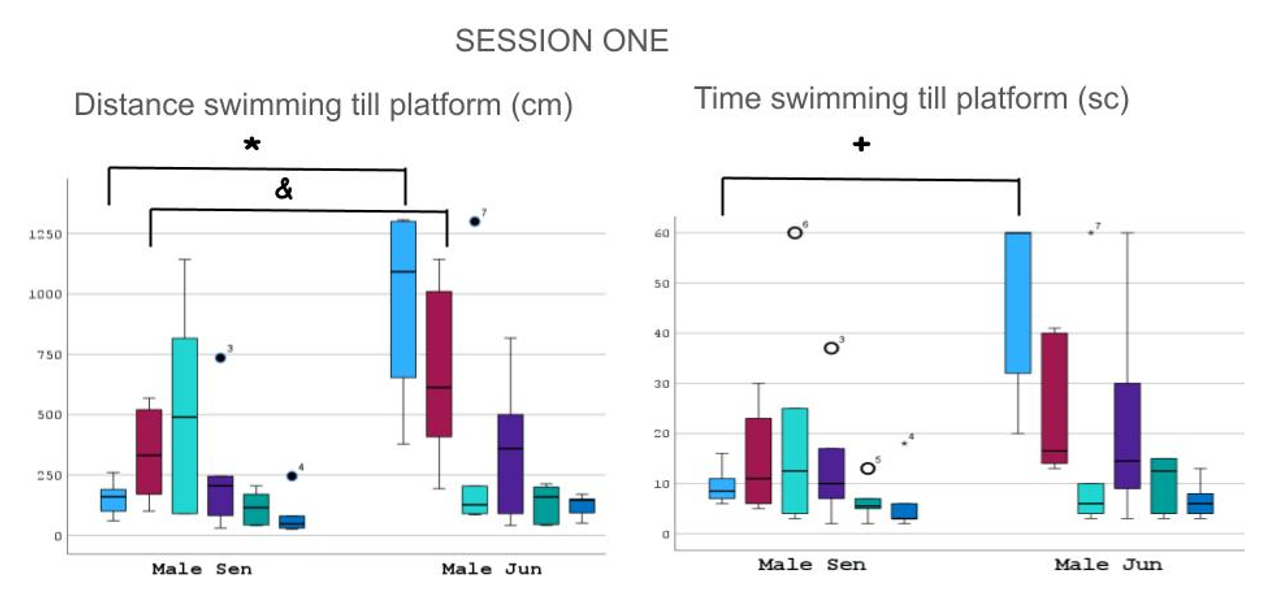
* p < 0.004 in trial 1; + p < 0.003 in trial 1; & p < 0.05 in trial 2. Figure 4: Graphs showing different length in centimeters (left) and time in seconds (right) the animals of both groups worked during swimming tests. There was statistical significance for the first trial, maybe because of the immaturity, bewilderment or apparent fragility of the junior group. However, they were rapidly able to reach the senior group in marks.
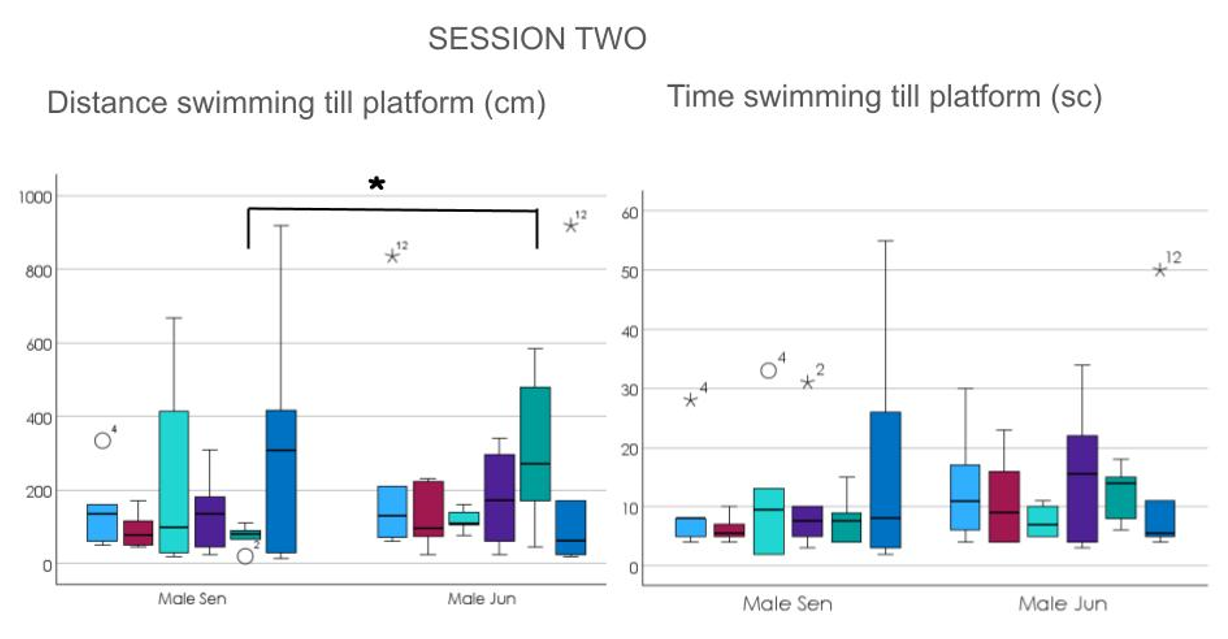
For the second session after 13 days another statistical difference was found in trial 5 for distance [F (1,12) = 4.333, p = 0.037], having the juniors the longer path swan (Figures
5 & 6).But both groups were almost equally good in their performances.
Because the animals were always swimming in the water and not still, the more time they were inside the pool, the longer they swam. That is why there was always a positive correlation inside each trial between the time and distance in session 1: Trial 1 [X2 (1, N = 13) = 0.98; p < 0.001]; Trial
2: [X2 (1, N = 13) = 0.92; p < 0.001]; Trial 3: [X2 (1, N = 13) = 0.96; p < 0.001]; Trial 4: [X2 (1, N = 13) = 0.9; p < 0.001]; Trial 5: [X2 (1, N = 13) = 0.87; p < 0.001]; Trial 6: [X2 (1, N = 13) = 0.77; p < 0.002] (Figure 7).
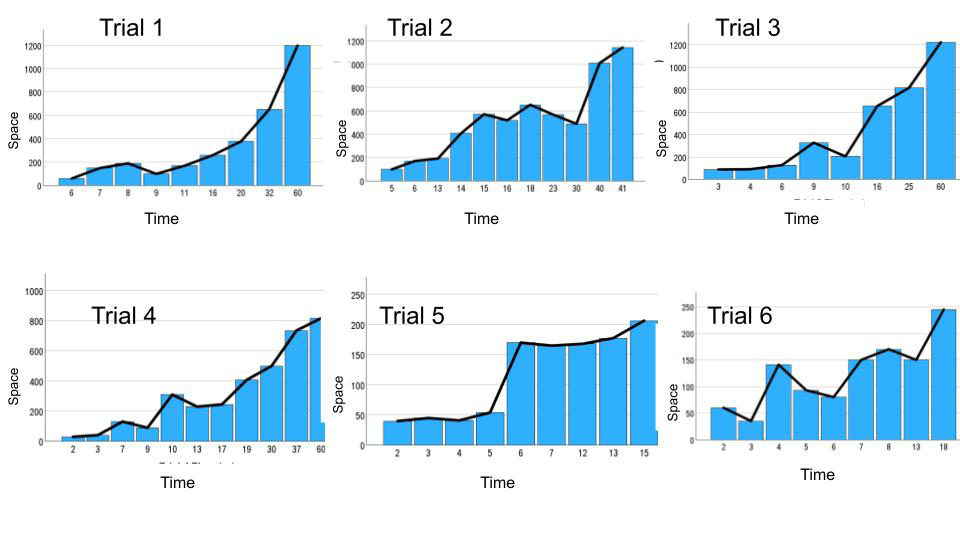
Figure 7: Graphs depicting the positive correlation existing in every trial (from 1 to 6 top left to inferior right corner) between time in the water swimming and the distance traveled. Very rarely the animal was still and calm in the water, then this correlation is pointing out their constant physical activity till finding the escape in the platform.
The Apodemus sylvaticus, being of a smaller size than the other mice, did the performance almost equally to the other groups, in trials 4, 5 and 6 in the first session but with certain differences: his swimming strategy was clearly different from the other groups. In trial 4 he found the platform as the other mice the first 20 seconds, in trial 5 he found the platform before second 10 as a point of support rather than a surface to stand on and in trial 6, after swimming about 50 seconds, with his nails of the hind and front legs he chose to grab the wall of the labyrinth to rest for a while as if it were the same as on the platform. Referring to his memory between trials, Sylvester Apodemus Sylvaticus did not remember the position of the platform in any of the previous trials. Those wild escape strategies were not seen in the domestic mice (Figure 8). These individual results were not included for the group comparisons.
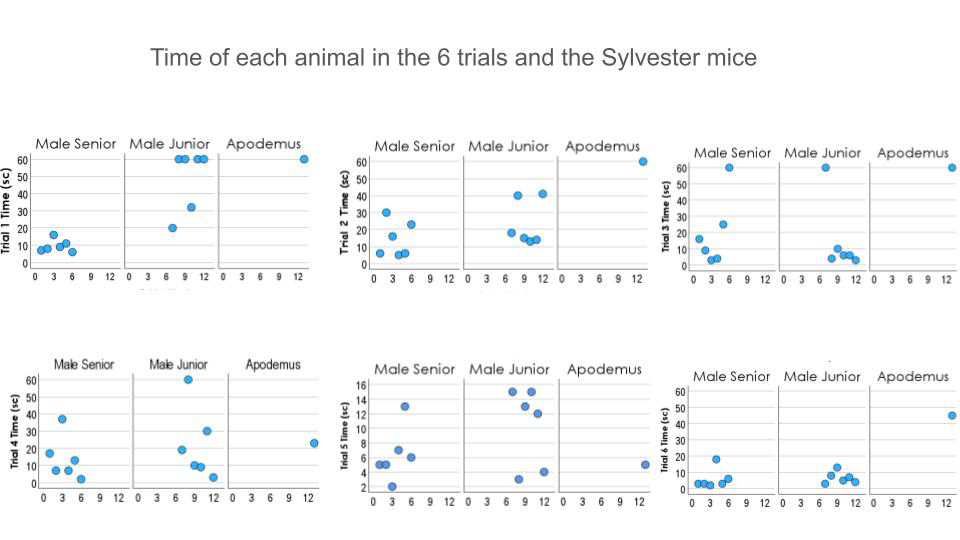
Figure 8: In these graphs we can see the Sylvester Apodemus Sylvaticus performance (animal # 13; third column in every graph) in time of swimming till finding the platform compared with the other two groups. From trial 4 forward (inferior line), the performance of the Apodemus Sylvaticus mouse would be the same as the other groups. However, the swimming strategies were different from the domestic groups.
Referring to the memory of the platform location through different trials, we found a clear statistical difference in second session, where both groups, junior and senior, were more often closer to the higher values or remembering (80%) (Session 1 vs Session 2: H = 4,641; p = 0.031) (Figure 9). With both groups together it is noticeable how the percentage of memory is bigger with more trials, to get almost the complete memory of the maze and the platform locations (Figures 10).
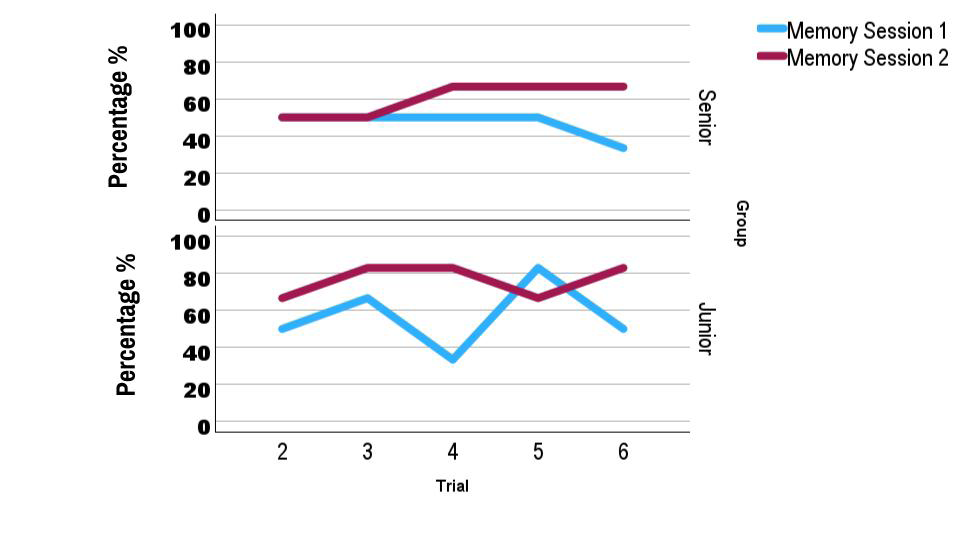
Figure 9: Graphs showing the percentage of animals that remembered the position of the platform in the previous trial (memory), in two different groups (Senior vs Junior) and in both consecutive sessions. In both groups there is an improvement of memory, since the percentage of remembering was closer to 80 % only in the second session (purple line) and for both groups. Session # 2 was significantly different from the first one (p = 0.031).
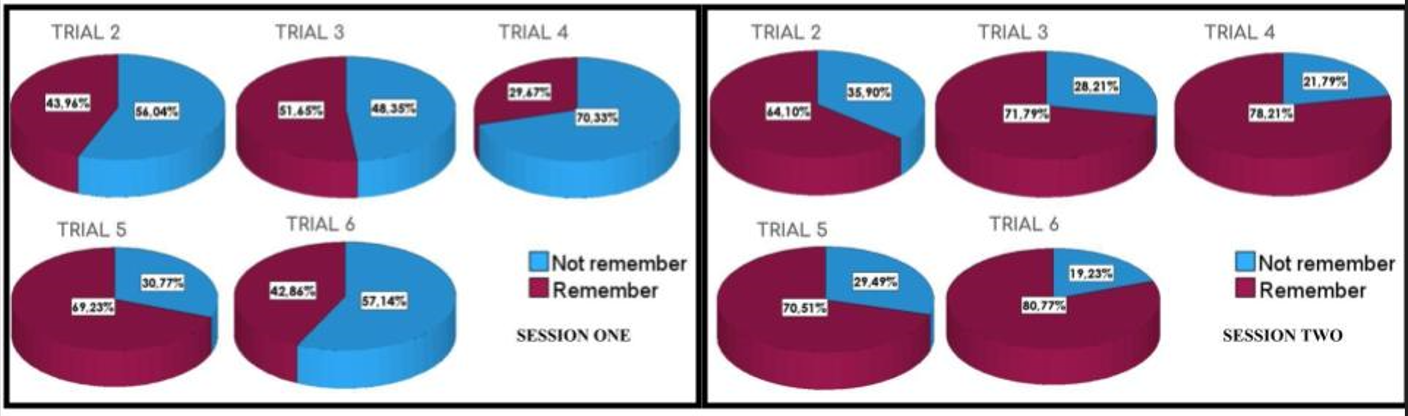
Figure 10: Graphs showing the percentage of memory in two different groups and in both consecutive sessions. It is noticeable how the percentage of remembering (purple color) is increasing more and more with trials and with experiences through different sessions through time. It shows how animal behaviors and likely their brain connections, are learning strategies and getting more and more comprehension of the situation, the environment and their escape alternatives (memory for a cognitive map).
Discussion
The formation of cognitive maps is not an easy goal and it clearly affects the quality of driving and the safety of drivers. Its importance relies on the fact that confidence is a very relevant tool to make the right choices when speed is bigger than our common velocity, that is which one our brains are prepared for. However, very often we trust artificial methods to define the path we will cross by without giving extra attention to the external stimuli that are essential to build a solid cognitive map.
There is a greater GPS habit of use that might cause an impairment on our hippocampus connections for navigation and this would be of special and relevant importance in those stages of life when the brain structure is getting formed. Our mammalian “inner GPS” is very relevant for our success in movements and survival in changing environments. An artificial tool that substitutes that biological function and underactivates our underlying brain mechanisms through the use of external GPS might affect human cognitive map formation and perhaps other brain functions related. In driving schools for different motor vehicles (cars, motorbikes, or even boats), could be included a teaching area dedicated to the use of biological GPS with “artisanal” forms of orientation using compasses and other physical indications.
The cognitive map is motivated by “curiosity” rather than goal-directed action [13]. Curiosity is not neutral, however, it is dependent on several functions constrained biologically to balance opportunity such as real risk for life and possibilities of approaching reward or avoidance of pain/stress. Curiosity might be present in early stages of development. Motivational factors may provide the impetus and momentum to the creation of different cognitive maps and succeeding in this way might enforce the brain connection for that particular route. Repetition of the experience would replace motivation to find by seeking of reward for having found one. With the GPS instructions motivation is silenced and attention never flows beyond the closer sensations (thigmotaxis) to the formation of an escape route to the destination after a whole brain processing, which include new brain connections with the environment.
In this paper we show how aged different mice react statistically differently to the same stimuli (Water Maze), proving their brains are fully prepared to deal with these situations despite their lack of previous experience. Looking at the human drivers, the early use of GPS in new young drivers might have a detriment on the capacities to understand and create the cognitive maps which are suitable to improve their driving skills.
Researching with domestic mice has some particularities. These animals, in genes, behavior, brain material, etc. are very similar to experimental animals in an animal house of a lab (Mus Musculus). However, they are allowed to have a different treatment, more affectionate, by the researcher or caregivers, because they are like pets. This new perspective might be included in the animal experiments protocols of the labs, because we, researchers, cannot forget we are a sort of “rodent breeders”. In exchange for our care, rodents do not give milk, meat or eggs, but numbers, graphs and tables which are our livelihood and modus vivendi. This research is evidence that these mice are certainly good specimens for obtaining scientific behavioral and experimental data to provide information to the scientific community. As an ethical suggestion we would say that an inclusion of the mice domestic Mus Musculus sold in pet stores as lab rodent animals might increase the information for the neuroscientific community. That is certainly more ethical than being food to captive reptiles, especially when all the steps taken have been aimed to ensure the rodent’s well- being. They are an interesting strain to inform about rodent individual and social behaviors, but also genetically speaking to provide data about inbreeding or consanguinity.
Conclusions
The use of GPS for driving is very widespread nowadays, however, its function could go beyond helping us: the cognitive map and its configuration requires attention and skills that are relevant from the very beginning early in our mammals lifespans and as fast drivers. Preserving and protecting our own tools might be of relevance to defend our adaptation to the natural environment.
Acknowledgements
We would like to thank Prof. Swaab at the Netherlands Institute for Neuroscience for his good suggestions, didactic criteria and inspirational ideas. We would like to thank the reviewers for their improvements. Moreover, I would like to thank my cats who helped me to learn about the mice world.
References
-
Dahmani L, Bohbot VD (2020) Habitual use of GPS negatively impacts spatial memory during self-guided navigation. Sci Rep 10(1): 6310.
-
Seminati L, Hadnett HJ, Joiner R, Petrini K (2022) Multisensory GPS impact on spatial representation in an immersive virtual reality driving game. Sci Rep 12: 7401.
-
Raby AC, Antonini A, Pappas A, Dassanayake DT, Brownjohn JMW, et al. (2019) Wolf Rock lighthouse: past developments and future survivability under wave loading. Philos Trans A Math Phys Eng Sci 377(2155): 20190027.
-
Shapiro M (2015) A limited positioning system for memory. Hippocampus 25(6): 690-696.
-
Hazen EL, Friedlaender AS, Goldbogen JA (2015) Blue whales (Balaenoptera musculus) optimize foraging efficiency by balancing oxygen use and energy gain as a function of prey density. Sci Adv 1(9): e1500469.
-
Abrahms B, Hazen EL, Aikens EO, Savoca MS, Goldbogen JA, et al. (2019) Memory and resource tracking drive blue whale migrations. Proc Natl Acad Sci U S A 116(12): 5582-5587.
-
Iorio MV, Graham IM, Hewitt RC, Aarts G, Pirotta E, et al. (2022) Prey encounters and spatial memory influence use of foraging patches in a marine central place forager. Proc R Soc B 289(1970): 20212261.
-
Rai SP, Krohn M, Pahnke J (2020) Early Cognitive Training Rescues Remote Spatial Memory but Reduces Cognitive Flexibility in Alzheimer’s Disease Mice. J Alzheimers Dis 75(4): 1301-1317.
-
Usunoff KG, Itzev DE, Rolfs A, Schmitt O, Wree A (2006) Brain stem afferent connections of the amygdala in the rat with special references to a projection from the parabigeminal nucleus: a fluorescent retrograde tracing study. Anat Embryol (Berl) 211(5): 475-496.
-
Zocher S, Schilling S, Grzyb AN, Adusumilli VS, Bogado LJ, et al. (2020) Early-life environmental enrichment generates persistent individualized behavior in mice. Sci Adv 6(35): eabb1478
-
Bromley BK, Deng Y, Song W (2011) Morris water maze test for learning and memory deficits in Alzheimer’s disease model mice. J Vis Exp 20(53): 2920.
-
Vorhees CV, Williams MT (2006) Morris water maze: procedures for assessing spatial and related forms of learning and memory. Nat Protoc 1(2): 848-858.
-
Keefe JO (1990) A computational theory of the hippocampal cognitive map. Prog Brain Res 83: 301-312.
- Occupational Stress and Mental Health Outcomes Among Police Officers: A Mini Review
- The Experience of Counterproductive Leadership on Mental Health and Impact on Retention in U.S. Marines: A Phenomenological Study
- Nomophobia in the Digital Age: A Study on College and University Students
- Emotional Regulation in Children with Autism and Learning Disabilities
- Antisemitism on American College Campuses and Its Impact on Jewish Students
- Exploring the Role of Empathy in the Associations of Family Functioning and Purpose in Life with Attitude towards Abortion among Undergraduates: A Moderation Analysis