Beta-Lactamases and ESBL
Microbial resistance through extended-spectrum β-lactamase (ESBL) was first reported in the early 1980s in Europe and subsequently in the United States soon after the introduction of third-generation cephalosporins in clinical practice. β -Lactamases are enzymes that inactivate β -lactam antibiotics by opening the β-lactam ring of penicillins and cephalosporin and abolish their antimicrobial activity. Beta-lactamases have been described for many species of grampositive and gram-negative bacteria. Some β-lactamases are plasmid mediated while others are chromosomally mediated.
Introduction
Bacterial resistance to antibacterial agents is a condition in which there is no or decreased susceptibility to antibacterial agents that ordinarily cause inhibition of bacterial cell growth or death. Bacterial resistance to antibacterial agents is a quantitative measurement of the efficiency (concentration expressed in micrograms per millilitre or as inhibition zones in millimetres for the diffusion techniques) of an antibacterial agent against a specific bacterium. In vitro measurements of antibacterial activity are based on testing increasing concentrations of an antibacterial agent against a bacterial isolate to find out at which concentration the growth of the bacterium is inhibited. This is known as the minimum inhibitory concentration (MIC) of the drug [1].
The resistance mechanism may be expressed continuously whether an inciting challenge is present or not. This state is referred to as constitutive expression. In contrast, some genes must be “induced” by exposure to the challenge substance before they produce the induced gene product [2].
Some enzymes are secreted actively into the extracellular environment where they can exert their antibacterial action, such as the β-lactamases of staphylococci. In contrast, most of the enzymes of Gram- negative bacteria are cell-bound so that they exert their effects only if the antimicrobial agent enters the bacterial cell wall [3].
Beta-Lactamases
Definition: β-lactamases are bacterial enzymes that are encoded by chromosomal or by plasmid-borne genes, they protect the microorganisms against lethal effects of β-lactam antibiotics by hydrolyzing the β-lactam ring, thus rendering the drugs inert. Based on their amino acid sequences, β-lactamases can be classified into 4 different molecular groups, the Ambler classes A, B, C, and D [4]. Mechanism of Action of Β-Lactamases: The action of serine β-lactamases depends on the association of the enzyme noncovalently with the antibiotic to yield the noncovalent Michaelis complex (Figure 1). The β-lactam ring is then attacked by the free hydroxyl on the side chain of a serine residue at the active site of the enzyme, yielding a covalent acyl ester. Hydrolysis of the ester finally liberates active enzyme and the hydrolyzed inactive drug. This mechanism is followed by β- lactamases of molecular classes A, C, and D but class B enzymes (Metallo β-lactamases) utilize a zinc ion to attack the β-lactam ring [5].
![Figure 1: Mechanism of action of β-lactamases. Nomenclature: Β-Lactamase nomenclature has not followed rational rules. Some enymes have three or four- letter abbreviations. Some were named for properties related to some of the substrates they hydrolyze, such as CARB (for activity against carbenicillin) and OXA and IMP (for activity against oxacillins and imipenem respectively), and some were named for their biochemical properties, such as SHV (for “sulphhydryl variable”). NMC-A stands for “nonmetallo carbapenemase of class A.” The name CTX is an abbreviation of cefotaximase and refers to the potent hydrolytic activity of these enzymes against cefotaxime. The AmpC β-lactamases were named for the genes that encode them and hence for ampicillin hydrolysis. Other enzymes were named for a particular bacterium; for example, PSE refers to _Pseudomonas-_ specific enzymes and Sme refers to the _Serratia_ _marcescens_ enzyme [6].](/fulltextimages/4578/fig_1.png)
Figure 1: Mechanism of action of β-lactamases. Nomenclature: Β-Lactamase nomenclature has not followed rational rules. Some enymes have three or four- letter abbreviations. Some were named for properties related to some of the substrates they hydrolyze, such as CARB (for activity against carbenicillin) and OXA and IMP (for activity against oxacillins and imipenem respectively), and some were named for their biochemical properties, such as SHV (for “sulphhydryl variable”). NMC-A stands for “nonmetallo carbapenemase of class A.” The name CTX is an abbreviation of cefotaximase and refers to the potent hydrolytic activity of these enzymes against cefotaxime. The AmpC β-lactamases were named for the genes that encode them and hence for ampicillin hydrolysis. Other enzymes were named for a particular bacterium; for example, PSE refers to Pseudomonas- specific enzymes and Sme refers to the Serratia marcescens enzyme [6].
TEM was named after the patient from whom it was isolated in 1963 in Athens, Greece, a young female called Temoniera. The names of some other enzymes refer to a hospital, for example, MIR (for “Miriam Hospital”). The names of other β-lactamases refers to the pharmaceutical company and bacteria with which they are associated, such as FPM (for “Fujisawa’s Proteus mirabilis”), and others refer to the researchers who discovered them, such as HMS (for “Harris, Matthew, Sykes”). Thus the nomenclature does not always refer to the properties of these particular enzymes [7]. Classification: β-lactamases are most commonly classified according to two general schemes: the Ambler molecular classification scheme and the Bush-Jacoby- Medieros functional classification system.
The Ambler scheme divides β-lactamases into four major classes (A to D). The basis of this classification scheme rests upon protein homology (amino acid similarity), and not phenotypic characteristics. In the Ambler classification scheme, β-lactamases of classes A, C and D are active-site serine β-lactamases. But class B or metallo -β-lactamases need zinc to attack the β-lactam ring [8].
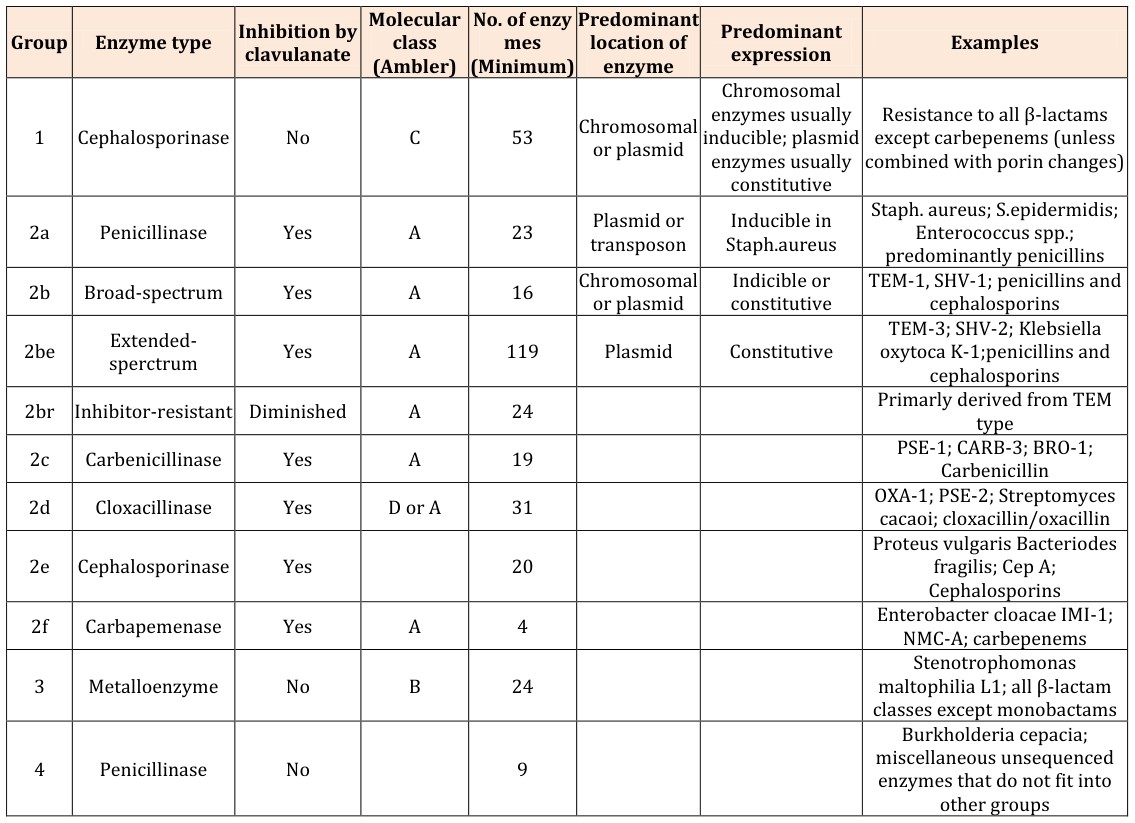
The Bush-Jacoby-Medeiros classification scheme (Table 1) groups β-lactamases according to functional similarities (substrate and inhibitor profile). There are four main groups and multiple subgroups in this system. This classification scheme is of much more immediate relevance to the physician or microbiologist in a diagnostic laboratory because it considers β-lactamase inhibitors and β-lactam substrates that are clinically relevant [9].
- Molecular
- No. of enzy
- Predominant
- Group
- Enzyme type
- Inhibition by
- Predominant class (Ambler) mes (Minimum) location of expression
- Examples clavulanate enzyme
- Chromosomal enzymes usually inducible; plasmid
- 1
- Cephalosporinase
- No
- C
- 53
- Chromosomal or plasmid enzymes usually constitutive
- 2a
- Penicillinase
- Yes
- A
- 23
- Plasmid or transposon
- Inducible in
- Staph.aureus
- 2b
- Broad-spectrum
- Yes
- A
- 16
- Chromosomal
- Indicible or constitutive or plasmid
- 2be
- Extendedsperctrum
- Yes
- A
- 119
- Plasmid
- Constitutive
- 2br Inhibitor-resistant Diminished
- A
- 24
- 2c
- Carbenicillinase
- Yes
- A
- 19
- 2d
- Cloxacillinase
- Yes
- D or A
- 31
- 2e
- Cephalosporinase
- Yes
- 20
- 2f
- Carbapemenase
- Yes
- A
- 4
- 3
- Metalloenzyme
- No
- B
- 24
- 4
- Penicillinase
- No
- 9
Table 1: Bush-Jacoby-Medeiros Functional Classification Scheme for β-lactamases (With Correlation to the Ambler
Definition: In Gram-negative pathogens, β-lactamases production remains the most important contributing factor to β-lactam resistance. Β- lactamases are bacterial enzymes that inactivate β-lactam antibiotics by hydrolysis, which results in ineffective compounds. One group of β-lactamases, extended-spectrum β lactamases (ESBLs), have the ability to hydrolyse and cause resistance to various types of the newer β-lactam antibiotics, including the expanded-spectrum (or third- generation) cephalosporins (eg, cefotaxime, ceftriaxone, ceftazidime) and monobactams (eg, aztreonam), but not the cephamycins (eg, cefoxitin and cefotetan) and carbapenems (eg, imipenem, meropenem and ertapenem). ESBLs are also inhibited by the β-lactamase inhibitors (clavulanate, tazobactam, sulbactam) Organisms that produce ESBLs remain an important reason for therapy failure with cephalospoins and have serious consequences for infection control. Therefore it is important that clinical microbiology laboratory detects and reports ESBLs [11]. Epidemiology: ESBL -producing organisms have been increasingly detected worldwide. Their prevalence varies from one country to another and from institution to institution. A survey on 81,213 bloodstream infecton pathogens during 1997 - 2002 showed that the Klebsiella spp. with an ESBL phenotype was isolated at a rate of 42.7% in Latin America, 21.7% in Europe and 5.8% in North America [12]. The Pan European Antimicrobial Resistance using Local Surveillance (PEARLS) study (2001 - 2002) showed that the percentages of ESBL production among E. coli and K. pneumoniae and Enterobacter spp. were 5.4, 18.2 and 8.8% respectively for all the study sites. The overall ESBL production rate for the combined Enterobacteriaceae was 10.5%. The highest rates were encountered in Egypt (25.8%) and Greece (27.4%) and lowest in the Netherlands (2%) and Germany (2.6%) [13, 14]. In Japan, the percentage of ESBL production in E.coli and K. pneumonia is low (5%) [15]. Elsewhere in Asia the percentage varies from 4.8% in Korea to 12% in Hong Kong [16]. In the Kingdom of Saudi Arabia in a tertiary care hospital in Riyadh, 48.4% of K. pneumoniae and 15.8% of E.coli blood culture isolates collected from January 2003 through December 2004 were ESBL producers [17]. ESBLs exist in every region of the world and in most genera of enterobacteria [18].
In Cairo, studied the presence of β-lactamases with an extended spectrum of activity in clinical Escherichia coli isolates [19]. Forty-six E.coli isolates were collected from patients with urinary tract infections at Al-Azhar University Hospital 2001. Phenotypic characterisation identified a very high ESBLs rate of 60.9%. Pulsed-field gel electrophoresis and plasmid profiles revealed eight different clonal groups. All ESBL producers were polymerase chain reaction-positive for _bla_TEM and _bla_CTX-M genes. Within the CTX-M family, three different enzymes, CTX-M-14, CTX-M-15 and CTX-M-27, were found. The ESBL producers carried multiple plasmids and further plasmid-encoded resistances. In several strains, genes for up to six aminoglycoside-modifying enzymes were detected. A linkage to fluoroquinolone resistance was not observed. This study confirms the high rate of ESBLs in Egypt and further demonstrates the worldwide spread of genes coding for CTX-M enzymes in clinical isolates.
ESBL Types
TEM Beta-Lactamases (class A): The TEM-type ESBL are derivatives of TEM-1and TEM-2. TEM-1, TEM-2 and TEM- 13 are not ESBLs. TEM-1 was first reported in 1965 from an E.coli isolate from a patient in Athens, Greece, named Temoneira (hence the designation TEM). More than 100 TEM-type ß-lactamases have been described, the majority of which are ESBLs. Although TEM-type ESBLs are most often found in E.coli and K.pneumoniae, they are also found in other species of Gram-negative bacteria with increasing frequency. TEM-type ESBLs have been reported in other genera of Enterobacteriaceae such as Enterobacter aerogenes, Enterobacter cloacae, Morganella morganii, Proteus mirabilis and Salmonella spp. Furthermore, TEM-type ESBL have been found in non-Enterobacteriaceae Gram - negative bacteriae, e.g., Pseudomonas aeruginosa [20]. SHV Beta-Lactamases (Class A): The SHV-type of ESBL may be found in clinical isolates more frequently than any other type of ESBLs. Unlike the TEM-type ß-lactamases, there are relatively few derivatives of SHV-1. The majority of SHV variants possessing an ESBL phenotype are characterized by the substitution of a serine for glycine at position 238. Also some have a substitution of lysine for glutamate at position 240. The serine residue at position 238 is critical for efficient hydrolysis of ceftazidime and lysine residue is critical for the efficient hydrolysis of cefotaxime. More than 50 SHV varieties are described worldwide. SHV-type of ESBLs has been detected in a wide range of Enterobacteriaceae. Outbreaks of SHV- producing Pseudomonas aeruginosa and Acinetobacter spp. have been reported [21]. CTX-M Beta-Lactamases (Class A) & Toho Beta- Lactamases: These enzymes were named for their greater activity against cefotaxime than other oxyimino- beta-lactam substrates (eg, ceftazidime, ceftriaxone, or cefepime). Rather than arising by mutation, they represent examples of plasmid acquisition of β-lactamase genes normally found on the chromosome of Kluyvera species, a group of rarely pathogenic commensal organisms. These enzymes are not very closely related to TEM or SHV β -lactamases in that they show only approximately 40% identity with these two commonly isolated β-lactamases. More than 40 CTX-M enzymes are currently known. Despite their name, few are more active on ceftazidime than cefotaxime. They have mainly been found in strains of Salmonella enterica serovar Typhimurium and E. coli, but also have been described in other species of Enterobacteriaceae and are the predominant ESBL type in parts of South America [22].
Toho-1 and Toho-2 are β-lactamases related structurally to CTX-M-type β-lactamases. Toho refers to the Toho University School of Medicine Omori Hospital in Tokyo, where a child who was infected with Toho-1 β- lactamase-producing Escherichia coli was hospitalized_. Like most CTX-M-type β-lactamases, the hydrolytic activity of the Toho-1 and Toho-2 enzymes is more potent against cefotaxime than ceftazidime [23]. _OXA Beta-Lactamases (Class D): The OXA-type ß- lactamases are another growing family of ESBLs. The OXA-type ESBLs were originally discovered in Pseudomonas aeruginosa isolates from a single hospital in Ankara, Turkey. Several of the OXA-type ESBLs have been derived from the original OXA-10 ß-lactamase (e.g., OXA -11, 14, 6 and 17). In contrast to the majority of OXA- type ESBLs, which confer resistance to ceftazidime, the OXA-17 ß-lactamase confers resistance to cefotaxime and ceftriaxone but provides only marginal protection against ceftazidime. A novel ESBL (OXA-18) was reported to be inhibited by clavulanic acid. Many of the newer members of OXA-type of ESBLs have been found mainly in Pseudomonas aeruginosa isolates originating from Turkey and France [24]. PER-Type Beta-Lactamases: The PER-type ESBLs share only around 25 to 27% homology with known TEM- and SHV-type ESBLs. PER-1 β-lactamase efficiently hydrolyses penicillins and cephalosporins and is susceptible to clavulanic acid inhibition. PER-1 was first detected in Pseudomonas aeruginosa, and later in Salmonella enteric serovar Typhimurium and Acinetobacter isolates as well. In Turkey, as many as 46% of nosocomial isolates of Acinetobacter spp. and 11% of Pseudomonas aeruginosa were found to produce PER-1. Isolates of Pseudomonas aeruginosa producing PER-1 have been detected in France, Italy, and Belgium. Additionally, a high prevalence of PER-1 in Acinetobacter spp. from Korea has been noted. PER-2, which shares 86% homology to PER-1, has been detected in S.enterica serovar Typhimurium, Escherichia coli, Klebsiella pneumonia, Proteus mirabilis, and Vibrio cholera O1 El Tor. PER-2 has only been found in South America [18]. VEB-1, BES-1 and Other ESBLs: A variety of other β- lactamases which are plasmid- mediated or integron- associated class A enzymes have been discovered. They are not simple point mutant derivatives of any known β- lactamases. They are remarkable for their geographic diversity. They include VEB-1which has greatest homology with PER-1 and PER-2(38%). It confers high- level resistance to ceftazidime, cefotaxime, and aztreonam, which is reversed by clavulanic acid. The gene encoding VEB-1 was found to be plasmid mediated; such plasmids also confer resistance to non- β-lactam antibiotics. The patient from whom the β-lactamase was originally described was a Vietnamese infant hospitalized in France. An identical β-lactamase has also been found in Escherichia coli, Klebsiella pneumonia, Enterobacter cloacae, Enterobacter sakazakii, and Pseudomonas aeruginosa isolates in Thailand. Other VEB enzymes have also been detected in Kuwait and China [25]. GES, BES, TLA, SFO, and IBC are other examples of non- TEM, non-SHV ESBLs and have been found in a wide range of geographic locations [26].
References
-
Livermore DM (2003) Bacterial resistance, origins, epidemiology and impact. Clin Infect Dis 36(1): 811- 823.
-
Maiden MC (1998) Horizontal genetic exchange, evolution and spread of antibiotic resistance in bacteria. Clin Infect Dis 27: 12-20.
-
Hall RM, Partridge SR (2001) Evolution of multiple antibiotic resistances by acquisition of new genes. In: Antibiotic Development and Resistance. Hughes D, Andersson DL, (Eds.), Taylor & Francis, London, United Kingdom, pp: 37-51.
-
Lamotte Brasseur JJ, Knox J, Kelly A, Paulette C, Eveline F, et al. (1994) The structures and catalytic mechanisms of active-site serine beta-lactamases. Biotechnol Genet Eng Rev 12(1): 189-230.
-
Waley SG (1992) β-Lactamase: Mechanism of action. In: The chemistry of β-Lactams. Page MI (ed.), London, pp: 198-228.
-
Mascaretti OA (2003) General mechanisms of bacterial resistance. In: Bacteria versus Antibacterial Agents: An Integrated Approach. In: Mascaretti OA, (Ed.), ASM Press Washington DC, pp: 89-90.
-
Shah AA, Hasan F, Ahmed S, Hameed A (2004) Extended-Spectrum β-Lactamases (ESBLs): Characterization, epidemiology and detection. Crit Rev Microbiol 30(1): 25-32.
-
Ambler R P (1980) The structure of β-Lactamases. Philos Trans R Soc Lond 289(1036): 321-331.
-
Bush K, Jacoby GA, Medeiros AA (1995) A functional classification scheme for β-Lactamases and its correlation with molecular structure. Antimicrob Agents Chemother 39(6): 1211-1233.
-
Bradford PA (2001) Extended spectrum beta- lactamases in the 21st century: Characterization, epidemiology and the detection of this important resistance threat. Clin Microbiol Rev 14(4): 933-951.
-
Pituot JD, Laupland KB (2008) Extended spectrum beta-lactamase-producing Enterobacteriaceae: An emerging public- health concern. Lancet Infect Dis 8(3): 159-166.
-
Beidenbach DJ, Moet GJ, Jones RN (2004) Occurrence and antimicorbial resistance pattern comparisons among bloodstream infection isolates from the SENTRY Antimicrobial Surveillance Program (1997 - 2002). Diagn Microb Infect Dis 50(1): 59-69.
-
Omar M, Badawi H, Helmy H (2001) Comparison of different methods for detection of extended spectrum β-lactamases (ESBLs) and their genetic relatedness among Enterobacteriaceae clinical isolates in a research medical institute. Egypt J Med Microbiol, pp: 365-374.
-
Bouchillon SK, Johnson BM, Hoban DJ, Johnson JL, Dowzicky MJ, et al. (2004) Determining incidence of extended-spectrum ß-lactamase producing Enterobacteriaceae, vancomycin-resistant Enterococcus faecium and methicillin-resistant Staphylococcus aureus in 38 centres from 17 countries: The PEARLS Study 2001-2002. Int J Antimicrob Agents 24(2): 119-124.
-
Bell JM, Turnidge JD, Gales AC, Pfaller MA, Jones RN, et al. (2002) Prevalence of extended spectrum beta- lactamase (ESBL)-producing clinical isolates in the Asia-Pacific region and South Africa: Regional results from SENTRY Antimicrobial Surveillance Program (1998-1999). Diagn Microbiol Infect Dis 42(3): 193- 198.
-
Lee SH, Kim JY, Shin SH, An YJ, Choi YW, et al. (2003) Dissemination of SHV-12 and characterization of new AmpC-type beta-lactamase genes among clinical isolates of Enterobacter species in Korea. J Clin Microbiol 41(6): 2477-2482.
-
El Khizzi NA, Bakheshwain SM (2006) Prevalence of extended spectrum beta-lactamases among Enterobacteriaceae isolated from blood culture in a tertiary care hospital. Saudi Med J 27(1): 37-40.
-
Paterson DL, Bonomo RA (2005) Extended spectrum β-lactamases: A clinical update. Clin Microbiol Rev 18(4): 657-686.
-
Mohamed Al Agamy MH, El Din Ashour MS, Wiegand I (2006) Firstdescription of CTX-M β-lactamase- producing clinical Escherichia coli isolates from Egypt. Int J Antimicrob Agents 27(6): 545-548.
-
Jacoby GA, Munoz Price L (2005) Mechanisms of disease: The new beta-lactamases. N Engl J Med 352(4): 380-391.
-
Paterson DL, Hujer KM, Hujer AM, Yeiser B, Bonomo MD, et al. (2003) Extended-spectrum β-lactamases in Klebsiella pneumoniae bloodstream isolates from seven countries: Dominance and widespread prevalence of SHV- and CTX-M-type β-lactamases. Antimicrob Agents Chemother 47(11): 3554-3560.
-
Karisik E, Ellington MJ, Pike R, Warren RE, Livermore DM, et al. (2007) Molecular characterisation of plasmids from CTX-M-15 producing Escherichia coli strains responsible for outbreaks in the United Kingdom. Fifteenth European Congress of Clinical Microbiology and Infectious Diseases Oxford United Kingdom 58(3): 1134-01-64.
-
Ma L, Ishii Y, Ishiguro M, Matsuzawa H, Yamaguchi K (1998) Cloning and sequencing of the gene encoding Toho-2, a class A ß-lactamase preferentially inhibited by tazobactam. Antimicrob Agents Chemother 42(5): 1181-1186.
-
Poirel L, Girlich D, Naas T, Nordmann P (2001) OXA- 28, an extended-spectrum variant of OXA-10 beta- lactamase from Pseudomonas aeruginosa and its plasmid- and integron- located gene. Antimicrob Agents Chemother 45(2): 447-453.
-
Jiang X, Ni Y, Jiang Y, Yuan F, Han L, et al. (2005) Outbreak of infection caused by Enterobacter cloacae producing the novel VEB-3 beta-lactamase in China. J Clin Microbiol 43(2): 826-831.
-
Castanheira M, Mendes RE, Walsh TR, Gales AC, Jones RN (2004) Emergence of the extended-spectrum beta-lactamase GES-1 in a Pseudomonas aeruginosa strain from Brazil: Report from the SENTRY Antimicrobial Surveillance Program. Antimicrob Agents Chemother 48(6): 2344-2345.
- hMPV: Is It Another Covid-19 Like Situation?
- Streptomyces: Sources of Novel Discoveries in Antibiotic Research to Combat Antimicrobial Resistance
- A Review of Mosquitoes (Diptera: Culicidae) and Their Biodiversity, Medical and Veterinary Importance
- Past and Current Immunotherapy in Cancer
- Hematological Cancer and Viral Infection
- The Growing Threat of Antimicrobial Resistance in India: Challenges and Solutions