Vaccine Strain of Measles Virus: A Tool for Cancer Therapy
Measles virus, the causative agent of measles, possesses a latently oncotropic character. This character enables the virus to infect, syntialize and lyse the cancer cells. The live attenuated strains, specially the Edmonston strain, recognize CD46 as their receptor. CD46 is overexpressed in adenocarcinoma cells. There is a basal level of expression of CD46 in all nucleated cells. The Edmonton vaccine strain has high affinity towards CD46 receptor but the wild stain has strong affinity towards CD150/ SLAM expressed on the lymphoid cells and epithelial nectin-4. This natural property of the live attenuated vaccine strains of Measles virus is exploited for oncolytic virotherapy. In recent years, virus-mediated oncolytic virotherapy has emerged as most reassuring therapy against carcinoma. Without causing any damage to the neighbouring tissues, the oncolytic Measles virus destroys the cancer cells through self-replication. Genetically modified viruses have been generated to broaden the field of oncolytic virotherapy. Previously non-engineered strains of MeV were used for conducting oncolytic virotherapyhighlighting the need for enhancing efficacy and safety. But with the emergence of reverse genetics system, the viral genome can be manipulated to develop genetically modified strains and this has brought an improvement in the therapeutic index. In this review we will discuss how the genetically engineered MeV strains are generated for oncolytic virotherapy as well as the possible outcomes of this modern therapeutic approach. We will also discuss about MeV mediated oncolytic immunotherapy in this review.
Introduction
Measles virus (MeV) is an enveloped virus with single- stranded, non-segmented, negative-sense RNA as genetic material. It belongs to the Paramyxoviridae family of the order Mononegavirales and grouped into the Morbillivirus genus- the viral neuraminidase is absent in Measles virus. Its RNA genome has a length of 15894 nucleotides. Six structural and two non-structural proteins are encoded by the genome. The structural proteins are Nucleocapsid protein (N), Phosphoprotein (P), Matrix protein (M), Fusion Protein (F), Hemagglutinin Protein (H) an Large Protein which is the RNA-dependent RNA polymerase (RdRp, L). The two non-structural proteins are V and C protein, encoded from the ORF of P protein gene.
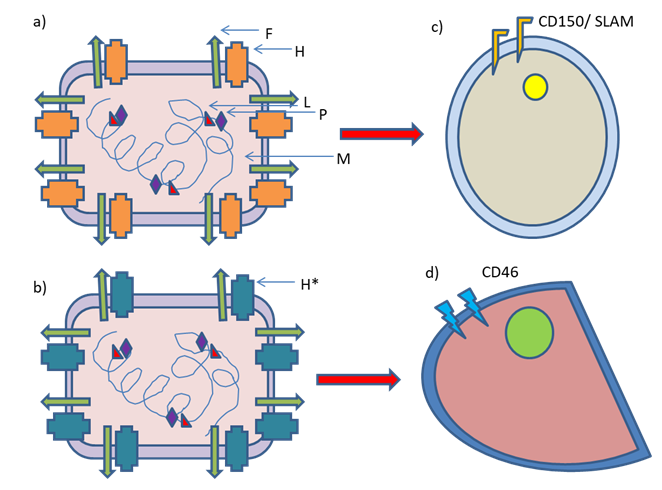
F and H are glycoproteins that are involved in fusion with the plasma membrane and binding to the viral receptors respectively. CD150/SLAM expressed on the lymphoid cells and epithelial nectin-4 serve as receptors for the wild type MeV, while MeV vaccine strains enter the cell mainly via CD46 [1]. This high affinity of H protein of MeV vaccine strain towards CD46 is because of mutations in the gene encoding this protein [2, 3]. MeV infection causes formation of syncytia- a distinctive cytopathic effect. The release of tumour- associated antigens after cell lysis is reported; thereby a tumour microenvironment having immunostimulatory effect is induced [4].
Various case reports have revealed that measles infection is associated with tumour remission. This idea has been exploited for treating cancer affected patients with MeV [5]. The example that was highly cited related to an unnamed boy, who was suffering from Burkitt’s lymphoma [6]. By observing these natural phenomena the idea of utilizing MeV for the treatment of cancer was bloomed, though measles is highly contagious [7]. The wild strain MeV is highly pathogenic and the question of using it in oncolytic virotherapy hardly arises. In 1960 the license to use the live attenuated strains of MeV for cancer treatment was passed and it was proved to be safe [8]. The testing of the derivatives of Edmonston B strain of MeV was commenced after several years. The haematological malignancies were selected as targeted bodies during early studies [9, 10]. The natural lymphotropic property of MeV supported this idea [11]. Wild-type MeV enters various immune cells viz., B and T cells, dendritic cells and macrophages using CD150/SLAM as receptor, thereby suppressing the host immune system [12]. Glioblastoma was reported to be sensitive to oncolysis mediated by MeV, sparing the normal cells [13].
When the replication-competent viruses are considered as therapeutic factors for cancer treatment, a critical importance should be given to tumour specificity for ensuring therapeutic efficacy as well as patient safety. Oncolytic viruses that are greatly effective against a specific carcinoma may be devoid of tumour specificity that results into significant off-target replication leading to toxicity have little or no applicability for clinical purposes. Oncolytic viruses that possess the capability of binding to different cell types or that are sequestered in the liver may not appear in sufficient numbers in tumour cells, thereby limiting their efficiency, especially after systemic administration. Two main approaches were introduced for addressing the matter of tumour specificity- the viruses possessing natural oncotropism were selected and tumour specificity was ensured by genetic modification, thereby generating engineered strains. The cell carriers having tumour-homing potentialities are being utilized for this very purpose as another option [14, 15, 16, 17].
Oncolytic MeV is effective against a wide range of cancers [18]. Various other measles vaccine strains are utilized for therapeutic purposes besides Edmonston B derivatives. They are Edmonston-Zagreb and AIK-C [19], Moraten-Schwarz [20], Leningrad-16 [21] as well as rMV-Hu191 [22] strains. The oncolytic effects of these above mentioned strains have been established in preclinical studies.
Thus, MeV mediated oncolytic virotherapy is proved to be one of the promising oncolytic platforms developed currently for the therapy of cancer. The advantage of using engineered MeV vaccine strains includes an outstanding safety profile lacking genotoxicity and ensuring immunogenicity. The reverse genetics system of MeV offers ensuring engineering possibilities.
Mutations in H Protein
Signalling lymphocyte activation molecule (SLAM) was reported to be a cellular receptor for wild-type MeV. In the recombinant wild-type MeV asparagine at the 481 position of the Hemagglutinin protein was substituted with tyrosine. The N481Y substitution enabled the recombinant wild-type virus to use CD46 as the second receptor. It was reported that its ability to utilize CD46 was comparatively low in CD46+ SLAM- cell lines compared to the recombinant virus having the Hemagglutinin Protein coding gene of the Edmonston laboratory strain. The N481Y substitution is not enough alone for making SLAM-tropic MeVs to utilize CD46 efficiently. This suggests that further substitutions are needed in the Hemagglutinin for their efficient infection and replication in CD46+ cells [3].
It was further reported that three substitutions (E492G and N390I plus T446S or N416D) besides N481Y substitution in the H protein of Ichinose-B (IC-B) strain (pathogenic MeV isolated from a patient suffering from acute measles in 1984) are necessary to utilize CD46 receptor efficiently [23]. The presence of N416D, N390I and T446S substitutions is reported in all strains belonging to Edmonston lineage but E492G substitution is present only in the H protein of cDNAs generated for Edmonston tag strain [2, 23]. In some Edmonston-lineage strain T484N substitution has been reported. Regarding the utilization of CD46 T484N substitution has a similar effect as that created by the E492G substitution [2, 23, 24].
Thus, the mutations in the multiple residues of the H protein have significant roles in the utilization of CD46 as receptor. These mutations enable the vaccine strains to enter into adenocarcinoma cells, keeping the immune cells almost safe- the vaccine strains are antigenic, never immunogenic. The specificities of wild-type and vaccine strain of MeV are depicted in Figure 1.
Genetically Engineered Measles Strains
Cancers, characterized by uncontrolled and abnormal cell division, are classified according to the tissue of origin. Cancer originating from epithelial tissue is termed as carcinoma. Cancer originating from connective tissue is termed as sarcoma. Cancer originating from hematopoietic cells is termed as leukaemia and lymphoma. Cancer originating from pluripotent cells is termed as germ cell tumour. Lastly, cancer originating from embryonic tissue or precursor cells is termed as blastoma [25, 26].
Measles virus was isolated from Edmonston, a patient by Peebles and Enders in 1954 [27]. Today Schwarz and Moraten strains are used- these are developed from further passages [28]. The unmodified Edmonston Vaccine Strain is utilized for the treatment of several cancers viz. blood cancers like leukaemia and myeloma and gynaecological cancers like breast and ovarian cancers. Genetically engineered strains are used for cancer therapy. Table 1 represents the genetically engineered measles virus used to treat cancer [28, 29].
Desired modifications of vaccine strains of MeV for enhancing virotherapy was performed by developing a reverse genetics system after rescuing the modified strains from cloned cDNA [31]. We will discuss this topic vividly in a separate section. In this system transgenes are inserted by added transcription units that are equipped with regulatory sequences of viral polymerase [32]. H protein of MeV is solely responsible for receptor attachment. To target entry, the H protein of MeV vaccine strain is mutated for ablating its natural tropism and targeting moieties such as single-chain variable fragments of antibody are fused to H for redirecting it to proper destination. The insertion is introduced at the C-terminal of H protein. The ligand that is introduced is a single chain antibody of the receptors that are expressed by cancer cells e.g. CD133, CD38, CD20, carcinoembryonic antigen (CEA) and epidermal growth factor receptor (EGFR) [33, 34, 35].
| Cancer Type | Genetically Modified Viral Strains |
| Lung Cancer | rMV-EGFP-SLAMblind, MV-CEA |
| Thyroid Cancer | MV-NIS |
| Blood Cancer | MV-PNP, MV-CD20, MV-LacZ |
| Myeloma | MV-lambda, MV-NIS, MV-CD38 |
| Burkitt's Lymphoma | MV-GMCSF |
| Breast Cancer | MV-CEA, MV-NAP |
| Ovarian Cancer | MV-NIS, MV-CEA, MV-CD38, MV-her/neu |
| Liver Cancer | NV-NIS, MV-CEA, MV-CD133, MV-MMP |
| Pancreatic Cancer | MV-NIS, MV-PNP |
| Prostate Cancer | MV-CEA, MV-NIS |
| Colorectal Cancer | MV-PNP, MV-NIS, MV-CD133 |
| Skin Cancer | MV-alphaPDL1 |
| Fibrosarcoma | MV-CD20, MV-MMP |
| Mesothelioma | MV-GMCSF, MV-NIS |
| Brain Cancer | MV-CD133, MV-CD38 |
Table 1: Genetically engineered viral strains utilized for oncolytic virotherapy [30].
In the serum of MV-CEA vaccine treated patients, the level of the soluble peptide, CEA, that is released into the patient’s blood stream following infection of cancer cells with MV-CEA, can be detected. CEA is a reporter gene that is inserted to track MeV replication. Another reporter gene β-human chorionic gonadotropin (HCG) was selected in this context [36]. Thyroidal sodium iodide symporter (NIS), a membrane ion channel, mediates uptake of iodine by thyroid glands. Genetically engineered MeV expressing NIS possesses the capability of inducing symport of iodine, thereby increasing iodine concentration inside cancer cells of thyroid gland. Encoding of NIS to yield MV-NIS enabled to conduct γ-camera imaging of iodine-123 (123I) [37, 38, 39, 40]. For ensuring invasion to spread all over the body, matrix metalloproteinases (MMP) are secreted by cancer cells for degrading extracellular matrix. Genetically engineered MeV harbouring MMP-cleavable sequences replacing furin- cleavable site of F protein ensures selective fusion of the engineered virus with MMP-expressing cancer cells [41].
Here F protein is engineered instead of H protein. In E. coli, Purine Nucleoside Phosphorylase (PNP) was reported to be present. It is a prodrug convertase assisting in converting chemotherapeutic prodrugs i.e., fludarabine into 2-fluoroadenine (reported to be a highly toxic metabolite), that intercalates into DNA, thereby promoting apoptosis. Genetically engineered MeV expressing prodrug convertase was prepared [42]. Genetically modified measles virus that expresses the immunomodulatory neutrophil activating protein (NAP) of H. pylori, induces proinflammatory cytokine release for boosting up immune response against cancer thereby ensuring prolonged survival [43]. Genetically modified MeV harbouring LacZ reporter gene is used for transduction in the tumour cells to quantify tumour development and spread of cancer. The beta-galactosidase enzymatic activity is recorded to be higher in metastatic cancer cells [44, 45]. Recombinant Measles virus armed with Granulocyte macrophage colony stimulating factor (GM-CSF) was reported to up-regulate anti-tumour activity of neutrophil by increasing its infiltration [46]. There is an overexpression of CD274 (programmed death ligand 1) in cancer cells to enable them to escape immune response against tumour. When cancer therapy is conducted in combination with the modulators of immune checkpoint, direct cell lysis reduces the tumour burden and there is a stimulation of anti-tumour immunity. Recombinant measles virus harbouring PDL1 antibody gene is proved to be an efficient lytic agent against melanoma [47]. MV-lambda is a genetically modified MeV strain expressing human lambda light immunoglobulin chain. This modified viral strain is used to treat multiple myeloma effectively. In myeloma cells there is an over-expression of human kappa light immunoglobulin chain. The secretion of a chimeric immunoglobulin (one kappa and one lambda chain) by the infected cells was reported. This modified marker can be easily quantified by using immunological techniques [48]. The consequences of the clinical trials are promising. The formation of syncytia that is mediated by F protein destroys the cancer cells [49].
The artificial riboswitches and microRNA target sites are under investigation for the case to target post-entry of MeV. The global loss in expression of microRNA levels in malignant versus healthy cells is regarded as a hallmark of cancer that is coming to light. This concept has been exploited to increase tumour specificity by engineering oncolytic viruses. The underlying concept is to insert the targeting sequences of the microRNAs that are absent in tumour cells but are exhibited in healthy tissues into the oncoviral genome [50, 51, 52, 53, 54]. It was shown that the above mentioned approach represses engineered MeV in healthy tissues expressing microRNAs, keeping the kinetics of replication as well as ensuing destruction of the cancer cells unchanged [55, 56, 57]. A simple overview of the engineered MeV and its mode of action are described in Figure 2.
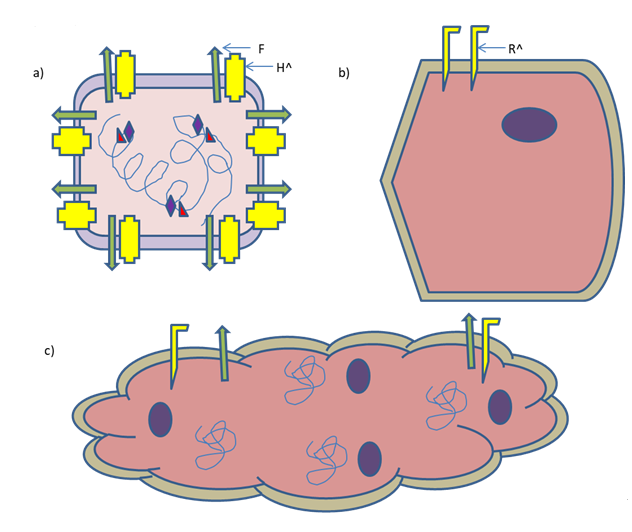
Figure 2: An Engineered MeV killing a cancer cell by syncytia formation. a) Engineered MeV displaying specific ligand of the respective receptor on a cancer cell. H^ indicates insertion of the gene of that specific ligand. b) A cancer cell expressing specific receptor (R^) to be recognized by the engineered virus. c) Engineered MeV killing cancer cell by syncytia formation. N.B. Presence of viral F protein on syncytium.
Combined Therapeutic Approaches
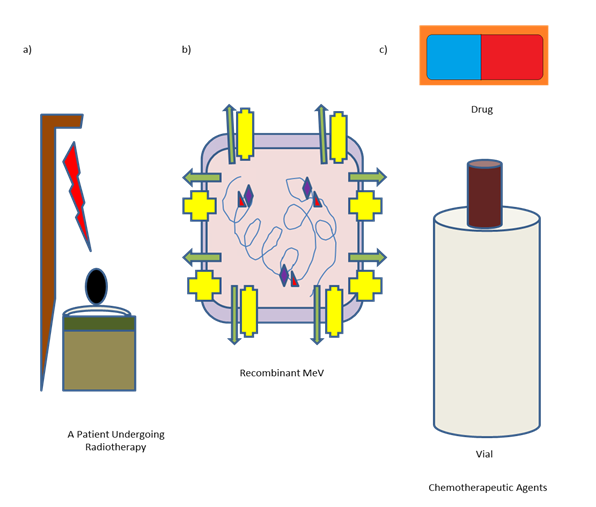
However, for curing advanced stage malignancies, using of oncolytic MeV alone for therapeutic purpose was proved to be insufficient. The combined therapeutic approaches are developed by modern medical oncologists to ensure an effective treatment for advanced stage malignancies. So, other established therapeutic approaches against cancer like chemotherapy and radiation therapy are combined with MeV mediated virotherapy [58].
Synergistic effects of radiotherapy with engineered MeV against Glioblastoma were observed in a Xenograft model as well as in vitro culture [59]. It has been demonstrated in vitro that the combinations of chemotherapies viz., gemcitabine, camptothecin and paclitaxel with engineered MeV were successful [60, 61, 62]. The antitumor efficacy was reported to be increased in laryngeal cancer models when a combination of engineered MeV with nimotuzumab, an anti-epidermal growth factor receptor (EGFR) monoclonal antibody was administered successfully [63]. The compounds that are reported to modulate cellular metabolism are also tested for conducting combined approaches. Using dichloroacetate to block aerobic glycolysis was reported to stimulate cell death when administered with engineered MeV [64]. For promoting MeV oncolysis, induction of autophagy was proposed as a combined approach [65]. The host cell factors are reported to be modulated by several small molecules for enhancing engineered MeV mediated oncolysis. The heat shock protein (Hsp) 70 is upregulated upon MeV infection. Combination treatment of oncolytic MeV with an Hsp 90 inhibitor resulted into an increase in Hsp70 expression ensuring an enhanced apoptosis [66, 67, 68]. MeV replication and syncytia formation are associated with actin cytoskeleton remodelling. If Rho- associated coiled-coil forming kinase (ROCK) is inhibited during the treatment of breast, prostate and Glioblastoma cancer cells with engineered MeV, an increased yield in viral replication spread and killing of tumour cells have been observed [69]. In vitro replication of MeV is enhanced when ruxolitinib, a janus-associated kinase (JAK) inhibitor is used to counteract the IFN response [70]. The efficacy of engineered MeV was reported to be increased upon inhibition of histone deacetylase (HDAC) mediated epigenetic modulation. Here in the case of hepatocellular carcinoma the induction of Interferon and their Stimulated Genes (ISGs) are prevented [71] and the underlying mechanism remained unsolved so far in the case of pancreatic adenocarcinoma [72].
In pancreatic ductal adenocarcinoma (PDAC) the expression of miR-148a is reported to be downregulated, though in numerous healthy tissues present in the gastrointestinal tract it is expressed. The target sequences of the said microRNA are inserted together with cytosine deaminase-uracil phosphoribosyl transferase (CD-UPRT), an E.coli-derived prodrug-converting enzyme, into oncolytic MeV genome. The microRNA target sites are reported to restrict replication and spread of the engineered virus in the cells expressing miR-148a, while it allows unchanged oncolytic efficiency in PDAC cell lines. Non-toxic 5-FC (5-fluorocytosin) is converted into chemotherapeutic drug 5-FU (5-fluorouracil) by CD-UPRT (prodrug convertase), thereby ensuring a localized chemovirotherapy. The superior anti-tumour efficiency of the said engineered virus when combined to 5-FC was demonstrated by the Leber group both in vitro as well as in vivo. The feasibility for generating dually modified oncolytic MeV for enhanced efficacy and safety has been demonstrated by this approach [73].
Bispecific T cell engagers, BiTEs contain two antibody single-chain variable fragments (scFv) that bind to CD3 as well as a tumour surface antigen on T cells to mediate tumour- directed cytotoxicity of T cells. The clinical efficacy of BiTEs has been demonstrated against haematological malignancies. However, problems with delivery as well as toxicities have restricted wide application, also to treat solid tumours, so far. It was reported by the Heidelberg team that the engineered MeV that is encoded with BiTEs has proved to be functional and promotes in vivo recruitment of endogenous T cells. MeV-BiTE has ensured prolonged survival in comparison to parental MeV, BiTE only and engineered MeV that encodes a non-binding BiTE. In Xenograft models derived from patients, the combination of adoptive immune cell transfer with MeV-BiTE has prolonged survival significantly when compared to monotherapies [74].
Lauer group has probed a combinatorial application by employing activated human NK cells (or PBMCs) along with oncolytic MeV-GFP in human sarcoma cell lines. It was demonstrated by the same group in an earlier preclinical study that MeV possesses a potent oncolytic property in paediatric sarcomas [75]. As there were also the presence of sarcoma cell lines showing primary resistance to viral oncolysis, ideas were generated towards the application of combination therapies. It was reported that the above mentioned combination resulted in enhancement of oncolysis of human sarcoma cell lines in comparison to the respective monotherapies. In addition to this, the activation of NK cells when co-cultured with A673 sarcoma cells infected with MeV was reported [76].
The Lauer group also investigated whether the senescent tumour cells induced by gemcitabine can be lysed with much effectively at the time of combinational chemo virotherapy. It was reported that a combination of engineered MeV and gemcitabine more effectively lysed different pancreatic cancer cell lines treated as compared to either of the above two treatments. Furthermore, tumour cell senescence induced by gemcitabine was not impeded by MeV [60].
Super cytosine deaminase (SCD) mediates conversion of 5-fluorocystosine (5-FC), a prodrug into 5-fluorouracil (5- FU), and then to 5-fluorouridine monophosphate (5-FUMP).
This process was reported to occur solely in cancer cells as the expression of SCD is tumour-restricted. In cancer cells the cellular enzymes further process 5-FUMP to produce toxic metabolites that are reported to interfere with DNA, RNA and protein synthesis as well as DNA repair mechanism- this leads to an enhancement in the tumoricidal bystander effects [77]. The Lauer team in Tübingen evaluated the potential efficacy as well as safety of MeV-SCD plus prodrug 5-FC in
Oncolytic Immunotherapy
The early efforts to engineer oncolytic MeV were primarily focussed on augmenting direct killing of tumour cells. In the recent past there has been a shift in strategy from mainly tumour killing to mainly immunotherapeutic therapy. This shift has been encouraged by the progress in cancer immunotherapy that revolutionized the field of medical oncology.
Oncolysis mediated by MeV per se shows pleiotropic effects (a phenomenon of a single gene influencing multiple traits) on the immune response against tumour and every phase of the ‘Cancer Immunity Cycle’ is supported by these effects [79]. The cell death induced by MeV oncolysis is reported to be immunogenic [80], thereby inducing a distinctive immunopeptidome [81] to promote cross- priming of T cell responses against tumour by plasmacytoid and conventional dendritic cells [82]. It was reported that MeV mediated oncolysis enhances tumour necrosis factor-related apoptosis-inducing ligand (TRAIL) induced cytotoxicity by plasmacytoid and myeloid DCs [83] as well as regulating macrophages towards an anticancer phenotype [84]. The activation of neutrophil also occurs that leads to the secretion of tumour necrosis factor (TNF)-α, IFN-α, IL-8, and monocyte chemoattractant protein (MCP)-1; expression of TRAIL and degranulation [85] - depending on the tumour may or may not be useful [86].
These immunotherapeutic effects are ensured by inserting into the MeV genome immunomodulatory transgenes. Engineered MeV is able to serve as a vector for delivering immunomodulators at the site of tumour that is proved to be highly toxic if administered systemically. The granulocyte macrophage colony stimulating factor, GM-CSF was reported to be the first immunomodulatory transgene in context to MeV and other oncolytic viruses. In MeV genome the cytokine granulocyte-macrophage colony-stimulating factor (GM-CSF) was coded for enhancing the function of neutrophil. The severe combined immunodeficient (SCID) mouse is used as a model system to study autoimmune diseases. In the said model system of human lymphoma, the engineered MeV that encodes murine GM-CSF was reported to be more efficiency which correlated with the infiltration of neutrophil in the tumour [87].
The SCID mice are devoid of T- and B- cells; hence the adaptive immune response after virotherapy was not combination with pembrolizumab in patients affected with stage III/IV non-small cell lung cancer (NSCLC). A novel combination therapy is able to enhance the anti-tumoral activity of pembrolizumab further [78]. A cartoon diagram of combined therapy was depicted in Figure 3.
scrutinized. By using an immunocompetent murine colon adenocarcinoma model this were re-examined using MC38cea cells that are reported to be permissive towards redirected MeV-antiCEA. In this study, it was demonstrated that MeV-antiCEA, which is armed with GM-CSF is associated with the complete and durable reemission as well as enhancement of overall survival in one-third population of mice. Mice undergoing treatment with MeV-GM-CSF-antiCEA were reported to reject tumours when re-challenged. This exhibited an adaptive memory immune response having long-lived protection [87, 88].
MC38cea is the first mouse model that is fully immunocompetent- this model was introduced as a prerequisite to investigate immunomodulatory MeV vectors for demonstrating the induction of adaptive immune responses that are tumour-specific [89]. This model contains murine colorectal adenocarcinoma MC38 that is syngeneic to C57BL/6 mice and stably expresses the carcinoembryonic antigen that is reported to be susceptible to CEA-targeted recombinant MeV [87]. In the above mentioned model, the treatment conducted with MV-GM-CSF ensures an extended survival in comparison to the control MeV. The tumour was completely reemitted in forty per cent of treated mice and they were protected subsequently from tumour re- engraftment- this indicates the effect of tumour vaccination. MV-GM-CSF treatment further enhanced an intratumoral infiltration of T cells and their tumour-specific responses. [90].
In Xenograft model of lymphoma, MV-GM-CSF promotes an increased neutrophil infiltration that is associated with tumour regression. It was shown that the immunomodulators further enhances innate immune activation during MeV oncolysis. A MeV vector that encodes IFN-β has got the ability in induction of immune infiltration as well as in the tumour microenvironment remodelling in mesothelioma Xenografts [91]. MeV that encodes the immunomodulatory factor of H. pylori, neutrophil-activating protein (NAP), ensures prolonged survival as well as helps in inducing a useful cytokine response in lung colonization models and breast cancer Xenograft [92].
Another immunocompetent model was developed in C57BL/6 mice. The transduction of B16 melanoma cells was performed to express CD20 surface antigen stably to conduct treatment with engineered MeV targeted against CD20. In this model, the antibodies of immune checkpoint are encoded against programmed cell death-ligand 1 (PD- L1) as well as cytotoxic T lymphocyte-associated-4 (CTLA- 4) by MeV vectors extended survival in comparison to MeV that encodes only the constant region of antibody [93]. The therapeutic standard of this application was demonstrated by systemic administration of the antibodies against PD-L1, PD-1 and CTLA-4 in combination [94].
MeV that encodes IL-12 is associated with complete remission of tumour (90% of treated mice) that was reported to be mediated by CD8+ effector T cell response [95]. MeV encoding IL-12 increases the affluence of CD8+ TILs. This T-cell subtype is essential for its efficiency. Engineered MeV that encodes IL-12 has a superior efficacy against tumour in B16-hCD46 and MC38cea tumour models in comparison to an IL-15-coding MeV variant. Oncolytic MeV vectors that encode an IL-15 super agonist were reported to mediate the activation of NK and T cells, but were not so effective compared to IL-12 encoded MeV [95, 96]. While we compare several immunomodulatory transgenes that included IL-10, IL-12, GM-CSF, a soluble form of CD80 and PD-L1/CTLA-4 antibodies; the engineered viruses armed with IL-12 or PD- L1 antibody were proved to be superior in their respective experiments [96].
This approach was adopted in immunotherapy against cancer. Here tumour-associated antigens are encoded in the MeV vector. MeV vectors that encode ovalbumin (OVA), regarded as model antigen or claudin-6, the tumour antigen in native state or associated with lentivirus-like particles were reported to associate with the induction of antigen- specific cellular and humoral immune responses in CD46- transgenic mice that are IFN-α receptor (IFNAR)-deficient as well as extending survival in B16-derived cancer models [97]. Employing tyrosinase-related protein-2 (TRP-2), a melanoma antigen and OVA, MeV vectors that encodes the full ORFs of antigens or specific immunodominant CD8+ epitope of antigens or epitope variants were produced. The epitope variants were targeted to the proteasome after secretion. Utilizing these MeV vectors, cognate T cell priming mediated by dendritic cell upon their activation was illustrated ex vivo [98].
It was noted that in cutaneous T cell lymphoma lesions there is a shift towards the T cell population that is Th1- biased after viral treatment [99]. In ovarian cancer patients undergone treatment with MV-NIS, IL-4 and IFN-γ responded against IGF binding protein 2 (IGFBP2) and FRα, the tumour antigens- ELISPOT assay was used to detect this response [100]. A rise in IFN-γ counts detected by ELISPOT assay against testis antigens of cancer was also noted in most of the tested multiple myeloma patients undergoing treatment with MV-NIS [101]. Clinical data suggesting that there is a boost in antitumor immunity by engineered MeV mediated virotherapy. The immune effector cells having anticancer property viz., CD8+ NKG2D+ cells or NK cells show the property of adoptive transfer- this property in combination with the oncolytic property of MeV were proved to be effective in conducting immunotherapy [102, 103].
To induce antigen specific T cell responses, MeV is being administered as a highly immunogenic and heterologous vaccine vector [104]. This approach has been utilized for the development of vaccines against diverse pathogens that included emerging SARS CoV-2 [105]. The clinical trials that are published so far tested only the oncolytic MeV, which does not encode any extra immunotherapeutic payloads. An overall therapeutic efficiency can be augmented with novel immunomodulatory vectors.
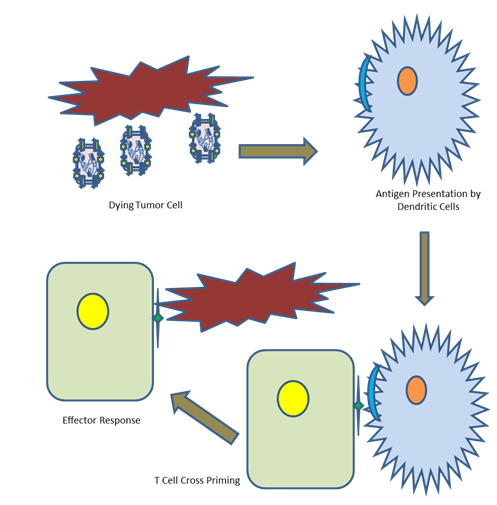
Figure 4: CD8+ cytotoxic T cell cross priming. Recombinant MeV mediated lysis of the cancer cell promotes the activation of dendritic cells, which leads to specific antigen presentation on its surface. This is followed by the cross priming of CD8+ cytotoxic T cells in the lymph nodes. This causes T cell infiltration to the tumour site, followed by an effector response.
Genetically engineered oncolytic viruses are reported to be multifaceted killers of tumour cells. They lyse tumour cells directly and spare normal cells. They are reported to enhance antitumor immunity indirectly by antigen release, thereby activate responses related to inflammation in the tumour microenvironment. There are some limitations too, such as, limited penetration into tumours, brief persistence, and the antiviral immune response by host. These are slowing down the universal translation of oncolytic virotherapy into the health centres. If these challenges are met, combined therapies, such as oncolytic viruses plus chimeric antigen receptor (CAR) T cells, immune checkpoint blockade (ICB), or CAR natural killer (NK) cells, may help provide potential therapeutic platforms in the health centres [106]. (Figure 4) depicts Immunovirotherapy in a nutshell.
Viral Rescue for Recombinant Measles
In reverse genetics approach, a gene of interest is selected, a null mutant of the selected gene is generated and the alternation in phenotype is studied. This approach is totally opposite to forward genetics- there screening of a collection of mutants for a known phenotype is conducted and the genes responsible for that phenotype are identified. By the refinement and development of the reverse genetics approach that allows rescue of MeV from cloned cDNA, the generation of genetically engineered variants of oncolytic MeV was made possible [107, 108].
Raedeke, et al. (1995) plasmids from which phage T7 RNA polymerase transcribes antigenomic RNAs of MeV with the correct termini were constructed. The 293-3-46 helper cells, expressing Measles N and P proteins as well as T7 RNA polymerase constitutively were derived from 293 cell lines. With constructs consisting of cDNA of viral polymerase as well as virus anti-genome that were kept under T7 promoter, these helper cells were co-transfected. Within these helper cells MeV replication and propagation occurred and this led to syncytia formation. To improve effectiveness in viral rescue a modified technique for rescuing the virus was adopted. The transfection media was changed; 14-16 hours after 293-3-46 helper cells were co-transfected with expression plasmids for viral protein. After that for 3 hours a heat shock at 440C was given to those helper cells. The diluted suspensions from these cells were then transferred to a Vero cell monolayer, in a 10 cm dish from each individual well of a six-well dish. The plaques were counted after 3-4 days of transfer. Following that the engineered virus was rescued [109]. The engineered viruses were specifically designed so that the progeny viruses would consist of a hexa-histidine tag at the C-terminal of H protein. This tag was identified by Vero-a – His cells. The Vero cells exhibited a single chain membrane bound antibody- the latter could identify the viral tag. This was named as a ‘pseudoreceptor system’ having use in viral rescue and multiplication [110].
Engineered viruses rescued by employment of the reverse genetics approach can be prepared for (a) infecting directly the cancer cells and lysing them, (b) delivering toxins to ensure the destruction of cancer cells and (c) delivering therapeutic agents at tumour site to target key cancer- specific molecules.
Clinical Trials Of Oncolytic Viruses With Special Focus To Measles Virus
Oncolytic virotherapy is not only confined to MeV but also many other viruses are used for this purpose. In a nut shell the most effective engineered oncolytic viruses used in clinical trials are described below.
The oncolytic virus, Rigvir was the foremost one to get an approval. In 2004 this was based on ECHO-7 virus for treating melanoma in Latvia [111]. H101 (Oncorine) was the second one approved for the treatment of nasopharyngeal carcinoma in China in 2005. This virus is an oncolytic Ad5 having a deletion of E1B-55kD [112]. In a multi-centre, where a controlled and randomized phase III trial was conducted with the combination of chemotherapy with H101 engineered strain, the objective response rate (ORR) was reported to be 78.8% in squamous cell carcinoma of head and neck (SCCHN) in comparison to 39.6% of the control group. This demonstrated an immense anti-tumour efficacy of the above mentioned combination therapy [113]. After it was launched in the market, the potential indications explored on lung cancer, pancreatic cancer, liver cancer and malignant pleural effusion in clinical trials showed certain effectiveness [114]. However, there was a slight impact of H101 on the market that might be the result of the favourable effectiveness of radiotherapy towards nasopharyngeal carcinoma. In recent past, talimogene laherparepvec (T-VEC) was reported to show a notably better ORR in stage IIIB- IV of melanoma that is unresectable. In 2015 the Food and Drug Administration (FDA) fast tracked this [115]. The live attenuated T-VEC is generated by an insertion of a human granulocyte–macrophage colony-stimulating factor (GM- CSF) cassette and deletions of the ICP34.5 and ICP47, in HSV 1 [116].
Early reports registering an oncolytic effect during natural MeV infections in Hodgkin’s disease, leukemia and Burkitt’s lymphoma appeared between the years 1971 and 1981 [117, 118, 119, 120, 121]. It was reported that the University Hospital of Zurich in Switzerland conducted the first clinical trial- the unmodified but attenuated MeV- Edmonston Zagreb (MV-EZ) strain was utilized in studying open-label, single institution, and dose escalation of phase I. Altogether 5 patients (1 female and 4 male) were injected with up to 16 intratumoral injections. The doses were fixed at 100, 500 and 1000 TCID50 (Median Tissue Culture Infectious Dose), following pre- treatment with interferon-α. The therapy was reported to be well tolerated without any dose-limiting toxicity. Five of 6 lesions injected with the above mentioned strain indicated a response to the treatment. One lesion displayed a perfect response; two of 5 distant lesions that were non-injected showed response [122].
From Mayo Clinic conducted clinical trials by utilizing the recombinant MeV strains in multiple myeloma and ovarian cancer [123, 124, 125]. A dose escalating study of phase I was conducted initially in 21 platinum refractory, heavily pre-treated women having recurrent ovarian cancer. They were administered intraperitoneally with MV-CEA having normal pre-treatment CEA levels. No dose-limiting toxicities were reported up to 109 TCID50 with 6 cycles at 4-weekly intervals. Fatigue, fever and abdominal pain were reported- this was grade 2 toxicity or less predominantly [125].
With MV-NIS a phase I/II clinical study was subsequently conducted on 16 women injected intraperitoneally with platinum and taxane refractory recurrent or invasive ovarian carcinoma was monitored. Like the above mentioned study, no dose-limiting toxicities were being reported. Unlike the previous study with MV-CEA that commenced with doses of 103 TCID50, here 108 or 109 TCID50 was received by the women. The most common harmful effects were fatigue, fever, abdominal and neutropenia [124]. In an initial report of a continuing phase I trial by using MV-NIS with or without cyclophosphamide in patients having refractory multiple myeloma (NCT00450814), 2 patients pre-treated heavily, revealed evidences of response to the treatment after they had received a single dose of 1011 TCID50 administered intravenously [123].
A Heidelberg Team chaired by Guy Ungerechts has generated an engineered oncolytic MeV encoding multiple transgene to increase therapeutic effectiveness. One of the top candidates in this context is the oncolytic MeV that encodes the secreted form of IL-12 (MeV-IL12). This engineered strain is currently used in clinical testing. In 2022 a phase I/II trial is reported to be launched in Heidelberg. The spin-off company of Heidelberg University Hospital, Canvirex AG has sponsored this trial to evaluate the immunovirotherapeutic effectiveness of MeV-IL12 on solid tumours [126, 127].
Another similar trial was organized by University Hospital Tübingen. The Lauer team has scheduled this trial in Tübingen. Here the potential efficacy as well as the safety of MV-SCD together with prodrug 5-FC in combination with pembrolizumab is assessed in patients affected with stage III/IV of non-small cell lung cancer (NSCLC) [128].
At the Mayo Clinic, the phase I/II trials in malignant peripheral nerve sheath tumour (NCT02700230), ovarian cancer by using mesenchymal stem cells of adipose tissue for virus delivery (NCT02068794), mesothelioma (NCT01503177), squamous cell cancer of the head and neck and breast cancer (NCT01846091) are on-going.
Delivery Route and Biosafety Measures
The possible cause of failure in treatment is suboptimal delivery. For oncolytic virotherapy numerous routes for delivering the oncolytic virus were examined. The therapeutic efficacy is critically dependent up on the selection of the therapeutic route. The scientists proposed different routes in accordance with their research goals as well as the accessible and vital experimental data.
The principle delivery routes of the engineered virus for oncolytic virotherapy are described below. (a) Intravenous delivery: Here the recombinant viruses are administered into the peripheral vein. The injected viruses reach tumour site in non-specific systems and organs via the vascular system. The engineered measles viruses expressing amino-terminal fragment of the urokinase plasminogen activator (uPA) (of human or mice) to target them against uPAR (MV-uPA) for treating breast cancer were administered intravenously [128, 129, 130, 131]. (b) Intratumoral delivery: Here the recombinant viruses are administered into tumours. The injected viruses are reported to exhibit a direct therapeutic outcome on the tumour lesion. Measles virus echistatin vector (MeV-ERV) was reported to be administered directly into the tumour cells to treat multiple myeloma [132, 133, 134]. (c) Intraperitoneal delivery: Here the recombinant viruses are administered into the peritoneal cavity. The injected viruses are reported to get absorbed into the peritoneal veins. This is followed by either arrival of the viruses in the tumour site of some systems or organs via the vascular system or getting diffused directly at tumour site in the peritoneal cavity. MV-αFV expressing a single-chain antibody (scFv) specific for α-folate receptor was targeted against ovarian cancer and was administered intraperitoneally [135, 136].
The common and severe adverse effects of MeV-mediated oncolysis are described below. MeV-NIS is used in the treatment of non-small cell lung cancer, multiple myeloma, malignant pleural mesothelioma and urothelial carcinoma. The common adverse effects are Grade I-II flulike symptoms, diarrhoea, leukopenia and neutropenia. The severe adverse effects are anaemia, neutropenia and leukopenia. MeV-CEA is used in the treatment of primary peritoneal cancer, ovarian epithelial cancer and fallopian tube cancer. The common adverse effects after virotherapy are flulike symptoms, lack or loss of appetite and abdominal pain. The severe adverse effect is pain in joint [137].
MV-NIS and MV-CEA: For maximizing infection of the engineered viruses in the tumour cells, MV-NIS and MV- CEA were injected intraperitoneally to the patients having recurrent ovarian carcinoma [138, 139]. In IFNARKO CD46 Ge mice that are measles naïve, pre-clinical studies were performed. No evidence of the propagation of virus to the outer surface of peritoneal cavity (heart, brain and skeletal muscle) was recorded [140]. Of the 21 women vaccinated with MeV-CEA the following response patterns were observed. In 4 asymptomatic patients low levels of viral genome were detected in peripheral blood mononuclear cells (PBMCs) by RT-qPCR [138]. In the 16 women administered with MeV- NIS, the viral genome was not detected in the peripheral blood [139]. The viral RNA was absent in saliva and urine [138, 140]. In another phase I clinical study, MV-NIS was administered intravenously with or without prior treatment with cyclophosphamide, a chemotherapeutic agent having an immunosuppressive activity in addition. Therefore, it ensures a potential enhancement and prolonging of viral replication and propagation in the tumour cells [141]. The presence of viral genome in buccal swabs of monkeys was reported post-administration. A comparatively higher level of virus genome was found to be present in the buccal swabs of monkeys treated with cyclophosphamide and viral genome was detectable for a longer time span, thereby indicating that the said drug is associated with the enhancement of the propagation of vectors in monkeys [142]. Intracerebral administration of engineered MeV expressing a single chain antibody recognizing EGFR against Glioblastoma multiforme was also reported [143].
Research on bacteriophage has been revitalized with the increase in antimicrobial resistance rates. These phages, discovered over some hundred years back, predate bacteria naturally. For using phages therapeutically, they should effectively kill the bacterial host, carry on lysis preferably and characterized fully for excluding side effects. To develop therapeutic phages a combined attempt of various stakeholders is needed. Here both naturally occurring as well as synthetic phages are used. A prophage was recently discovered- it was proved to be active against Helicobacter pylori. It can hopefully offer a therapy to be used for targeting the etiologic role of pylori in gastric ulcer as well as cancer.
Biosafety Issues: The term “biosafety” denotes the utilization of definite practices, safety apparatus, and buildings designed especially for ensuring that the environment, the community and the workers should be safeguarded from unintended exposure or accidental release of toxins, infectious agents and biological hazards. Various biosafety issues are required to be taken into consideration with utter care while clinical trials are performed with engineered attenuated MeV vectors. After carrying out a risk assessment extensively of the engineered MeV vectors utilized in the clinical trials presently as oncolytic vector or prophylactic vaccine, it can be concluded that for humans these engineered MeV vectors belong to risk 1 class. In order to boost up risk alleviation and avoid diffusion into the surroundings, manipulation of these engineered MeV vectors should be performed under CL-1 (Contaminant Level).
The record of safe utilization of MeV Schwarz strain and the attenuation in MeV-CHIK (Chikungunya fever virus, CHIKV) and MeV1-F4 (F4: a fusion protein consisting of the viral RT, p24, Nef, Ag p17) vaccine candidates permit handling these engineered viruses in the clinical setting under CL-1. The insertion of the transgenes hardly alters its safety profile. For handling MV-NIS and MV-CEA engineered oncolytic vectors, a CL-1 was approved. Engineered MV-NIS mediated transduction of Mesenchymal Stem Cells (MSC) should be conducted in CL-2. The MSC cells are primary cell cultures that are directly derived either from tissues or from organs. While manipulating those cells the presence of unintended contaminating factors accounts for the primary hazard. As finite life span is the characteristic of those cells, the time allotted for identifying and characterizing the contaminating factors remains restricted. So, the transduction of primary cell cultures obtained from human should be performed in a CL-2 and operated in a class II biosafety cabinet to guarantee the sterility of products. The personnel should wear protective equipment, masks, goggles and gloves. The operations that produce aerosols viz., the rupture of a vial must be avoided or contained strictly at the time of preparation and vaccination of the engineered MV vector. An appropriate disinfectant, 1% sodium hypochlorite must be used to inactivate the spills. Personal protective equipment and contaminated waste must be autoclaved before disposal.
For improving the biosafety of virus-mediated oncolytic therapy, the following three features may be taken into consideration. The first strategy involves the selection of non-infectious viruses that do not harm our normal tissues. The second strategy involves the attenuation of pathogenicity towards normal cells of the said viruses by introducing suitable genetic modifications. The third strategy involves recombining different varieties of oncolytic viruses for cancer treatment. As for example the recombination of Newcastle disease virus (NDV) with vesicular stomatitis virus (VSV) greatly helped in reduction of cytotoxicity in normal and healthy neurons and hepatocytes and was reported to be non-infectious to the embryonated eggs.
Limitations of Measles Mediated Virotherapy
However, one should remember that a lifelong vaccine against wild type measles is yet to be developed. The virus possessing a property to suppress the spread of cancer is still listed as one of the major reasons of death among children.
It was reported that the N protein has pro-apoptotic property. It was shown to interact with protein tyrosine kinase 2 for suppressing the GTPases expression viz., cdc42 and RhoA for checking the progression of cancer. But the P
protein was reported to have an antagonistic property. It was shown to interact with phosphoinositide 3-kinase, thereby the expression of anti-apoptotic markers viz. Bcl-xL and Bcl- 2 was increased. So it will be a cause of concern at the time of virotherapy. Therefore the limitations subsist in oncolytic virotherapy. They are immune responses by the host, both innate and adaptive, the range of hosts as well as the safety risks in using the engineered viruses.
Conclusion
Engineered MeV that was probed extensively in oncolytic virotherapy in the clinical trials was proved to be promising. Some clinical trials are still on-going and the recombinant viruses are showing high efficacy. The vaccine strains having an engineered genome for targeting the recombinant virus effectively to the malignant cells have an excellent safety record. Its progress can also be monitored, thereby making it an exciting as well as powerful candidate. Due to the lymphotropic property, MeV has been used as an attractive option for treating lymphoid malignancies particularly.
Once the vaccinologists were engaged in preparing a productive vaccine against measles but now the engineered vaccine strains of MeV are being used successfully as a potent therapeutic agent for the treatment of cancer. The clinical trials are going on in full-fledge and the feedback is positive. Besides radiotherapy and chemotherapy, oncolytic virotherapy has proved to be an effective tool for cancer therapy. The genetically modified virus possessing the capability of boosting the immune system associated with cancer suppression was developed. When the hepatocellular carcinoma cells are infected with Edmonston vaccine strain of MeV, the anti-cancer activity of CD8+ NKG2D+ cells is augmented and the said cells trigger the extrinsic pathway of apoptosis. Other viruses belonging to the Paramyxoviridae family also bear the same character and their tumour lysing property was examined vividly.
On-going efforts for addressing the subject of pre-existing antibodies against MeV in vaccinated patients have exhibited hopeful results in the pre-clinical studies, whereas the results from clinical studies are still awaited. A combination therapy, where engineered MeV is utilized in combination with added immune therapies, is proven to be interesting undoubtedly and the advancement to understand the cooperation between recombinant oncolytic MeV and the microenvironment of tumour is sure to improve its therapeutic efficacy further. Finally, although lots of mechanisms were proposed for MeV- mediated lysis of the tumour cells, the exact mechanism is yet to be discovered. Developments in this field will prove to be essential for upcoming directions in the utilization of MeV as a lytic agent against carcinoma.
When virotherapy is applied in combination with radiotherapy and chemotherapy, there is a synergistic effect.
References
-
Lin LT, Richardson CD (2016) The Host Cell Receptors for Measles Virus and Their Interaction with the Viral Hemagglutinin (H) Protein. Viruses 8(9): 250.
-
Hsu EC, Sarangi F, Iorio C, Sidhu MS, Udem SA, et al. (1998) Richardson C.D. A single amino acid change in the hemagglutinin protein of measles virus determines its ability to bind CD46 and reveals another receptor on marmoset B cells. J Virol 72(4): 2905-2916.
-
Seki F, Takeda M, Minagawa H, Yanagi Y (2006) Recombinant wild-type measles virus containing a single N481Y substitution in its haemagglutinin cannot use receptor CD46 as efficiently as that having the haemagglutinin of the Edmonston laboratory strain. J Gen Virol 87(6): 1643-1648.
-
Ungerechts G, Engeland CE, Buchholz CJ, Eberle J, Fechner H, et al. (2017) Virotherapy Research in Germany: From Engineering to Translation. Hum Gene Ther 28(10): 800-819.
-
Kelly E, Russell SJ (2007) History of Oncolytic Viruses: Genesis to Genetic Engineering. Mol Ther 15(4): 651- 659.
-
Bluming AZ, Ziegler JL (1971) Regression of burkitt’s lymphoma in association with measles infection. Lancet 2(7715): 105-106.
-
Moss WJ (2017) Measles. Lancet 390(10111): 2490- 2502.
-
Di Pietrantonj C, Rivetti A, Marchione P, Debalini MG, Demicheli V, et al. (2020) Vaccines for measles, mumps, rubella, and varicella in children. Cochrane Database Syst Rev 4(4): CD004407.
-
Peng KW, Donovan KA, Schneider U, Cattaneo R, Lust JA, et al. (2003) Oncolytic measles viruses displaying a single-chain antibody against CD38, a myeloma cell marker. Blood 101(7): 2557-2562.
-
Grote D, Cattaneo R, Fielding AK (2003) Neutrophils contribute to the measles virus-induced antitumor effect: Enhancement by granulocyte macrophage colony- stimulating factor expression. Cancer Res 63(19): 6463- 6468.
-
Peng KW, TenEyck CJ, Galanis E, Kalli KR, Hartmann LC, et al. (2002) Intraperitoneal therapy of ovarian cancer using an engineered measles virus. Cancer Res 62(16): 4656-4662.
-
Tatsuo H, Ono N, Tanaka K, Yanagi Y (2000) SLAM (CDw150) is a cellular receptor for measles virus. Nature 406(6798): 893-897.
-
Phuong LK, Allen C, Peng KW, Giannini C, Greiner S, et al. (2003) Use of a vaccine strain of measles virus genetically engineered to produce carcinoembryonic antigen as a novel therapeutic agent against glioblastoma multiforme. Cancer Res 63(10): 2462-2469.
-
Aref S, Bailey K, Fielding A (2016) Measles to the rescue: a review of oncolytic measles virus. Viruses 8(10): 294.
-
Bellini WJ, Rota JS, Rota PA (1994) Virology of measles virus. J Infect Dis 170(1): 15-23.
-
Ferguson MS, Lemoine NR, Wang Y (2012) Systemic delivery of oncolytic viruses: hopes and hurdles. Adv Virol 2012: 805629.
-
Kuruppu D, Tanabe KK (2005) Viral oncolysis by herpes simplex virus and other viruses. Cancer Biol Ther 4(5): 524-531.
-
Mühlebach MD (2020) Measles virus in cancer therapy. Curr Opin Virol 41: 85-97.
-
Zimmermann M, Armeanu S, Smirnow I, Kupka S, Wagner S, et al. (2009) Human precision-cut liver tumor slices as a tumor patient-individual predictive test system for oncolytic measles vaccine viruses. Int J Oncol 34(5): 1247-1256.
-
Gauvrit A, Brandler S, Sapede-Peroz C, Boisgerault N, Tangy F, et al. (2008) Measles Virus Induces Oncolysis of Mesothelioma Cells and Allows Dendritic Cells to Cross-Prime Tumor-Specific CD8 Response. Cancer Res 68(12): 4882-4892.
-
Ammour Y, Ryabaya O, Shchetinina Y, Prokofeva E, Gavrilova M, et al. (2020) The Susceptibility of Human Melanoma Cells to Infection with the Leningrad-16 Vaccine Strain of Measles Virus. Viruses 12(2): 173.
-
Lv Y, Zhou D, Hao XQ, Zhu MY, Zhang CD, et al. (2019) A recombinant measles virus vaccine strain rmv-hu191 has oncolytic effect against human gastric cancer by inducing apoptotic cell death requiring integrity of lipid raft microdomains. Cancer Lett 460: 108-118.
-
Tahara M, Takeda M, Seki F, Hashiguchi T, Yanagi Y, et al. (2006) Multiple Amino Acid Substitutions in Hemagglutinin Are Necessary for Wild-Type Measles Virus To Acquire the Ability To Use Receptor CD46 Efficiently. J Virol 81(6): 2564-2572.
-
Hsu EC, Sabatinos S, Hoedemaeker FJ, Rose DR, Richardson CD, et al. (1999) Use of site-specific mutagenesis and monoclonal antibodies to map regions of CD46 that interact with measles virus H protein. Virology 258(2): 314-326.
-
Hanahan D, Weinberg RA (2000) The hallmarks of cancer. Cell 100(1): 57-70.
-
Varricchio CG (2004) A cancer source book for nurses (8th Edn.), Jones and Bartlett Publishers. Boston pp: 229.
-
Enders JF, Katz SL, Milovanovic MV, Holloway A (1960) Studies on an attenuated measles-virus vaccine. I. Development and preparations of the vaccine: technics for assay of effects of vaccination. N Engl J Med 263: 153- 159.
-
Bhattacharjee S, Yadava PK (2018) Measles virus: Background and oncolytic virotherapy. Biochem Biophys Rep 13: 58-62.
-
Radecke F, Spielhofer P, Schneider H, Kaelin K, Huber M, et al. (1995) Rescue of measles viruses from cloned DNA. EMBO J 14(23): 5773-5784.
-
Pfaller CK, Cattaneo R, Schnell MJ (2015) Reverse genetics of Mononegavirales: How they work, new vaccines, and new cancer therapeutics. Virology 479-480: 331-344.
-
Bucheit AD, Kumar S, Grote DM, Lin Y, von Messling V, et al. (2003) An oncolytic measles virus engineered to enter cells through the CD20 antigen. Mol Ther 7(1): 62- 72.
-
Hammond AL, Plemper RK, Zhang J, Schneider U, Russell SJ, et al. (2001) Single-chain antibody displayed on a recombinant measles virus confers entry through the tumor-associated carcinoembryonic antigen. J Virol 75(5): 2087-2096.
-
Peng KW, Facteau S, Wegman T, O’Kane D, Russell SJ, et al. (2002) Non-invasive in vivo monitoring of track able viruses expressing soluble marker peptides. Nat Med 8(5): 527-531.
-
Peng KW, Holler PD, Orr BA, Kranz DM, Russell SJ, et al. (2004) Targeting virus entry and membrane fusion through specific peptide/MHC complexes using a high- affinity T-cell receptor. Gene Ther 11(15): 1234-1239.
-
Hadac EM, Peng KW, Nakamura T, Russell SJ (2004) Reengineering paramyxovirus tropism. Virology 329(2): 217-225.
-
Nakamura T, Peng KW, Harvey M, Greiner S, Lorimer IA, et al. (2005) Rescue and propagation of fully retargeted oncolytic measles viruses. Nat Biotechnol 23(2): 209- 214.
-
Watanabe M, Hirano A, Stenglein S, Nelson J, Thomas G, et al. (1995) Engineered serine protease inhibitor prevents furin-catalyzed activation of the fusion glycoprotein and production of infectious measles virus. J Virol 69(5): 3206-3210.
-
Springfeld C, von Messling V, Frenzke M, Ungerechts G, Buchholz CJ, et al. (2006) Oncolytic efficacy and enhanced safety of measles virus activated by tumor- secreted matrix metalloproteinases. Cancer Res 66(15): 7694-7700.
-
Ungerechts G, Springfeld C, Frenzke ME, Lampe J, Johnston PB, et al. (2007) Lymphoma chemovirotherapy: CD20-targeted and convertase-armed measles virus can synergize with fludarabine. Cancer Res 67(22): 10939- 10947.
-
Iankov ID, Allen C, Federspiel MJ, Myers RM, Peng KW, et al. (2012) Expression of immunomodulatory neutrophil- activating protein of helicobacter pylori enhances the antitumor activity of oncolytic measles virus. Mol Ther 20(6): 1139-1147.
-
Waszkiewicz N, Szajda SD, Waszkiewicz M, Wojtulewska- Supron A, Szulc A, et al. (2013) The activity of serum beta-galactosidase in colon cancer patients with a history of alcohol and nicotine dependence: preliminary data. Postepy Hig Med Dosw 67: 896-900.
-
Buller CJ, Zang XP, Howard EW, Pento JT (2003) Measurement of beta-galactosidase tissue levels in a tumor cell xenograft model. Methods Find Exp Clin Pharmacol 25(9): 713-716.
-
Grossardt C, Engeland CE, Bossow S, Halama N, Zaoui K, et al. (2013) Granulocyte-macrophage colony- stimulating factor-armed oncolytic measles virus is an effective therapeutic cancer vaccine. Hum Gene Ther 24(7): 644-654.
-
Engeland CE, Grossardt C, Veinalde R, Bossow S, Lutz D, et al. (2014) CTLA-4 and PD-L1 checkpoint blockade enhances oncolytic measles virus therapy. Mol Ther 22(11): 1949-1959.
-
Allen C, Vongpunsawad S, Nakamura T, James CD, Schroeder M, et al. (2006) Retargeted oncolytic measles strains entering via the EGFRvIII receptor maintain significant antitumor activity against gliomas with increased tumor specificity. Cancer Res 66(24): 11840- 11850.
-
Ylösmäki E, Hakkarainen T, Hemminki A, Visakorpi T, Andino R, et al. (2008) Generation of a conditionally replicating adenovirus based on targeted destruction of E1A mRNA by a cell type-specific MicroRNA. J Virol 82(22): 11009-11015.
-
Edge RE, Falls TJ, Brown CW, Lichty BD, Atkins H, et al. (2008) A let-7 MicroRNA-sensitive vesicular stomatitis virus demonstrates tumor-specific replication. Mol Ther 16(8): 1437-1443.
-
Kelly EJ, Hadac EM, Cullen BR, Russell SJ (2010) MicroRNA antagonism of the picornaviral life cycle: alternative mechanisms of interference. PLoS Pathog 6(3): e1000820.
-
Kelly EJ, Hadac EM, Greiner S, Russell SJ (2008) Engineering microRNA responsiveness to decrease virus pathogenicity. Nat Med 14(11): 1278-1283.
-
Kelly EJ, Nace R, Barber GN, Russell SJ (2010) Attenuation of vesicular stomatitis virus encephalitis through microRNA targeting. J Virol 84(3): 1550-1562.
-
Leber MF, Bossow S, Leonard VH, Zaoui K, Grossardt C, et al. (2011) MicroRNA-sensitive oncolytic measles viruses for cancer-specific vector tropism. Mol Ther 19(6): 1097-1106.
-
Baertsch MA, Leber MF, Bossow S, Singh M, Engeland CE, et al. (2014) MicroRNA-mediated multi-tissue detargeting of oncolytic measles virus. Cancer Gene Ther 21(9): 373-80.
-
Leber MF, Baertsch MA, Anker SC, Henkel L, Singh HM, et al. (2018) Enhanced control of oncolytic measles virus using MicroRNA target sites. Mol Ther Oncolytics 9: 30- 40.
-
Leber MF, Neault S, Jirovec E, Barkley R, Said A, et al. (2020) Engineering and combining oncolytic measles virus for cancer therapy. Cytokine Growth Factor Rev 56: 39-48.
-
Liu C, Sarkaria JN, Petell CA, Paraskevakou G, Zollman PJ, et al. (2007) Combination of measles virus virotherapy and radiation therapy has synergistic activity in the treatment of glioblastoma multiforme. Clin Cancer Res 13(23): 7155-7165.
-
May V, Berchtold S, Berger A, Venturelli S, Burkard M, et al. (2019) Chemovirotherapy for pancreatic cancer: Gemcitabine plus oncolytic measles vaccine virus. Oncol Lett 18(5): 5534-5542.
-
Tai CJ, Liu CH, Pan YC, Wong SH, Tai CJ, et al. (2019) Chemovirotherapeutic Treatment Using Camptothecin Enhances Oncolytic Measles Virus-Mediated Killing of Breast Cancer Cells. Sci Rep 9(1): 6767.
-
Lal G, Rajala MS (2019) Combination of Oncolytic Measles Virus Armed with BNiP3, a Pro-apoptotic Gene and Paclitaxel Induces Breast Cancer Cell Death. Front Oncol 8: 676.
-
Toan NL, Hang NT, Luu NK, VAN Mao C, VAN Ba N, et al. (2019) Combination of Vaccine Strain Measles Virus and Nimotuzumab in the Treatment of Laryngeal Cancer. Anticancer Res 39(7): 3727-3737.
-
Li C, Meng G, Su L, Chen A, Xia M, et al. (2015) Dichloroacetate blocks aerobic glycolytic adaptation to attenuated measles virus and promotes viral replication leading to enhanced oncolysis in glioblastoma. Oncotarget 6(3): 1544-1555.
-
Xia M, Gonzalez P, Li C, Meng G, Jiang A, et al. (2014) Mitophagy enhances oncolytic measles virus replication by mitigating DDX58/RIG-I-like receptor signaling. J Virol 88(9): 5152-5164.
-
Zou J, Guo Y, Guettouche T, Smith DF, Voellmy R, et al. (1998) Repression of heat shock transcription factor HSF1 activation by HSP90 (HSP90 complex) that forms a stress-sensitive complex with HSF1. Cell 94(4): 471-480.
-
Muchowski PJ, Wacker JL (2005) Modulation of neurodegeneration by molecular chaperones. Nat Rev Neurosci 6(1): 11-22.
-
Liu C, Erlichman C, McDonald CJ, Ingle JN, Zollman P, et al. (2008) Heat shock protein inhibitors increase the efficacy of measles virotherapy. Gene Ther 15(14): 1024-1034.
-
Opyrchal M, Allen C, Msaouel P, Iankov I, Galanis E, et al. (2013) Inhibition of Rho-associated coiled-coil-forming kinase increases efficacy of measles virotherapy. Cancer Gene Ther 20(11): 630-637.
-
Stewart CE, Randall RE, Adamson CS (2014) Inhibitors of the interferon response enhance virus replication in vitro. PLoS One 9(11): e112014.
-
Ruf B, Berchtold S, Venturelli S, Burkard M, Smirnow I, et al. (2015) Combination of the oral histone deacetylase inhibitor resminostat with oncolytic measles vaccine virus as a new option for epi-virotherapeutic treatment of hepatocellular carcinoma. Mol Ther Oncolytics 2: 15019.
-
Ellerhoff TP, Berchtold S, Venturelli S, Burkard M, Smirnow I, et al. (2016) Novel epi-virotherapeutic treatment of pancreatic cancer combining the oral histone deacetylase inhibitor resminostat with oncolytic measles vaccine virus. Int J Oncol 49(5): 1931-1944.
-
Singh HM, Leber MF, Bossow S, Engeland CE, Dessila J, et al. (2021) MicroRNA-sensitive Oncolytic Measles Virus for Chemovirotherapy of Pancreatic Cancer. Mol Ther Oncolytics 21: 340-355.
-
Heidbuechel JPW, Engeland CE (2021) Oncolytic viruses encoding bispecific T cell engagers: A blueprint for emerging immunovirotherapies. J Hematol Oncol 14(1): 63.
-
Berchtold S, Lampe J, Weiland T, Smirnow I, Schleicher S, et al. (2013) Innate immune defense defines susceptibility of sarcoma cells to measles vaccine virus- based oncolysis. J Virol 87(6): 3484-3501.
-
Klose C, Berchtold S, Schmidt M, Beil J, Smirnow I, et al. (2019) Biological treatment of pediatric sarcomas by combined virotherapy and NK cell therapy. BMC Cancer 19(1): 1172.
-
Lampe J, Bossow S, Weiland T, Smirnow I, Lehmann R, et al. (2013) An armed oncolytic measles vaccine virus eliminates human hepatoma cells independently of apoptosis. Gene Ther 20(11): 1033-1041.
-
Pidelaserra-Martí G, Engeland CE (2020) Mechanisms of measles virus oncolytic immunotherapy. Cytokine Growth Factor Rev 56: 28-38.
-
Donnelly OG, Errington-Mais F, Steele L, Hadac E, Jennings V, et al. (2013) Measles virus causes immunogenic cell death in human melanoma. Gene Ther 20(1): 7-15.
-
Rajaraman S, Canjuga D, Ghosh M, Codrea MC, Sieger R, et al. (2019) Measles Virus-Based Treatments Trigger a Pro-inflammatory Cascade and a Distinctive Immunopeptidome in Glioblastoma. Mol Ther Oncolytics 12: 147-161.
-
Guillerme JB, Boisgerault N, Roulois D, Ménager J, Combredet C, et al. (2013) Measles virus vaccine-infected tumor cells induce tumor antigen cross-presentation by human plasmacytoid dendritic cells. Clin Cancer Res 19(5): 1147-1158.
-
Achard C, Guillerme JB, Bruni D, Boisgerault N, Combredet C, et al. (2016) Oncolytic measles virus induces tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-mediated cytotoxicity by human myeloid and plasmacytoid dendritic cells. Oncoimmunology 6(1): e1261240.
-
Tan DQ, Zhang L, Ohba K, Ye M, Ichiyama K, et al. (2016) Macrophage response to oncolytic paramyxoviruses potentiates virus-mediated tumor cell killing. Eur J Immunol 46(4): 919-928.
-
Zhang Y, Patel B, Dey A, Ghorani E, Rai L, et al. (2012) Attenuated, oncolytic, but not wild-type measles virus infection has pleiotropic effects on human neutrophil function. J Immunol 188(3): 1002-1010.
-
Dey A, Zhang Y, Castleton AZ, Bailey K, Beaton B, et al. (2016) The Role of Neutrophils in Measles Virus- mediated Oncolysis Differs Between B-cell Malignancies and Is Not Always Enhanced by GCSF. Mol Ther 24(1): 184-192.
-
Grote D, Cattaneo R, Fielding AK (2003) Neutrophils contribute to the measles virus-induced antitumor effect: Enhancement by granulocyte macrophage colony- stimulating factor expression. Cancer Res 63(19): 6463- 6468.
-
Grossardt C, Engeland CE, Bossow S, Halama N, Zaoui K, et al. (2013) Granulocyte-macrophage colony- stimulating factor-armed oncolytic measles virus is an effective therapeutic cancer vaccine. Hum Gene Ther 24(7): 644-654.
-
Ungerechts G, Springfeld C, Frenzke ME, Lampe J, Parker WB, et al. (2007) An immunocompetent murine model for oncolysis with an armed and targeted measles virus. Mol Ther 15(11): 1991-1997.
-
Grossardt C, Engeland CE, Bossow S, Halama N, Zaoui K, et al. (2013) Granulocyte-macrophage colony- stimulating factor-armed oncolytic measles virus is an effective therapeutic cancer vaccine. Hum Gene Ther 24(7): 644-654.
-
Li H, Peng KW, Dingli D, Kratzke RA, Russell SJ, et al. (2010) Oncolytic measles viruses encoding interferon beta and the thyroidal sodium iodide symporter gene for mesothelioma virotherapy. Cancer Gene Ther 17(8): 550-558.
-
Iankov ID, Allen C, Federspiel MJ, Myers RM, Peng KW, et al. (2012) Expression of immunomodulatory neutrophil- activating protein of Helicobacter pylori enhances the antitumor activity of oncolytic measles virus. Mol Ther 20(6): 1139-1147.
-
Engeland CE, Grossardt C, Veinalde R, Bossow S, Lutz D, et al. (2014) CTLA-4 and PD-L1 checkpoint blockade enhances oncolytic measles virus therapy. Mol Ther 22(11): 1949-1959.
-
Hardcastle J, Mills L, Malo CS, Jin F, Kurokawa C, et al. (2017) Immunovirotherapy with measles virus strains in combination with anti-PD-1 antibody blockade enhances antitumor activity in glioblastoma treatment. Neuro Oncol 19(4): 493-502.
-
Backhaus PS, Veinalde R, Hartmann L, Dunder JE, Jeworowski LM, et al. (2019) Immunological Effects and Viral Gene Expression Determine the Efficacy of Oncolytic Measles Vaccines Encoding IL-12 or IL-15 Agonists. Viruses 11(10): 914.
-
Veinalde R, Grossardt C, Hartmann L, Bourgeois- Daigneault MC, Bell JC, et al. (2017) Oncolytic measles virus encoding interleukin-12 mediates potent antitumor effects through T cell activation. Oncoimmunology 6(4): e1285992.
-
Hutzler S, Erbar S, Jabulowsky RA, Hanauer JRH, Schnotz JH, et al. (2017) Antigen-specific oncolytic MV- based tumor vaccines through presentation of selected tumor-associated antigens on infected cells or virus-like particles. Sci Rep 7(1): 16892.
-
Busch E, Kubon KD, Mayer JKM, Pidelaserra-Martí G, Albert J, et al. (2020) Measles Vaccines Designed for Enhanced CD8(+) T Cell Activation. Viruses 12(2): 242.
-
Heinzerling L, Künzi V, Oberholzer PA, Kündig T, Naim H, et al. (2005) Oncolytic measles virus in cutaneous T-cell lymphomas mounts antitumor immune responses in vivo and targets interferon-resistant tumor cells. Blood 106(7): 2287-2294.
-
Galanis E, Atherton PJ, Maurer MJ, Knutson KL, Dowdy SC, et al. (2015) Oncolytic measles virus expressing the sodium iodide symporter to treat drug-resistant ovarian cancer. Cancer Res 75(1): 22-30.
-
Packiriswamy N, Upreti D, Zhou Y, Khan R, Miller A, et al. (2020) Oncolytic measles virus therapy enhances tumor antigen-specific T-cell responses in patients with multiple myeloma. Leukemia 34(12): 3310-3322.
-
Klose C, Berchtold S, Schmidt M, Beil J, Smirnow I, et al. (2019) Biological treatment of pediatric sarcomas by combined virotherapy and NK cell therapy. BMC Cancer 19(1): 1172.
-
Chen A, Zhang Y, Meng G, Jiang D, Zhang H, et al. (2017) Oncolytic measles virus enhances antitumour responses of adoptive CD8(+)NKG2D(+) cells in hepatocellular carcinoma treatment. Sci Rep 7(1): 5170.
-
Mühlebach MD (2017) Vaccine platform recombinant measles virus. Virus Genes 53(5): 733-740.
-
Hörner C, Schürmann C, Auste A, Ebenig A, Muraleedharan S, et al. (2020) A Highly Immunogenic Measles Virus-based Th1-biased COVID-19 Vaccine. Proc Natl Acad Sci U S A 117(51): 32657-32666.
-
Ma R, Li Z, Chiocca EA, Caligiuri MA, Yu J, et al. (2023) The emerging field of oncolytic virus-based cancer immunotherapy. Trends Cancer 9(2): 122-139.
-
Martin A, Staeheli P, Schneider U (2006) RNA polymerase II-controlled expression of antigenomic RNA enhances the rescue efficacies of two different members of the Mononegavirales independently of the site of viral genome replication. J Virol 80(12): 5708-5715.
-
Peng KW, Holler PD, Orr BA, Kranz DM, Russell SJ, et al. (2004) Targeting virus entry and membrane fusion through specific peptide/MHC complexes using a high- affinity T-cell receptor. Gene Ther 11(15): 1234-1239.
-
Alberts P, Tilgase A, Rasa A, Bandere K, Venskus D, et al. (2018) The Advent of Oncolytic Virotherapy in Oncology: The Rigvir® Story. Eur J Pharmacol 837: 117- 126.
-
Liang M (2018) Oncorine, the World First Oncolytic Virus Medicine and Its Update in China. Curr Cancer Drug Targets 18(2): 171-176.
-
Xia ZJ, Chang JH, Zhang L, Jiang WQ, Guan ZZ, et al. (2004) Phase III randomized clinical trial of intratumoral injection of E1B gene-deleted adenovirus (H101) combined with cisplatin-based chemotherapy in treating squamous cell cancer of head and neck or esophagus. Ai Zheng 23(12): 1666-1670.
-
Lin XJ, Li QJ, Lao XM, Yang H, Li SP, et al. (2015) Transarterial Injection of Recombinant Human Type- 5 Adenovirus H101 in Combination with Transarterial Chemoembolization (TACE) Improves Overall and Progressive-Free Survival in Unresectable Hepatocellular Carcinoma (HCC). BMC Cancer 15: 707.
-
Andtbacka RH, Kaufman HL, Collichio F, Amatruda T, Senzer N, et al. (2015) Talimogene Laherparepvec Improves Durable Response Rate in Patients With Advanced Melanoma. J Clin Oncol 33(25): 2780-2788.
-
Liu BL, Robinson M, Han ZQ, Branston RH, English C, et al. (2003) ICP34.5 Deleted Herpes Simplex Virus with Enhanced Oncolytic, Immune Stimulating, and Anti- Tumour Properties. Gene Ther 10(4): 292-303.
-
Pasquinucci G (1971) Possible effect of measles on leukaemia. Lancet 1(7690): 136.
-
Gross S (1971) Measles and leukaemia. Lancet 1(7695): 397-398.
-
Zygiert Z (1971) Hodgkin’s disease: remissions after measles. Lancet 1(7699): 593.
-
Taqi AM, Abdurrahman MB, Yakubu AM, Fleming AF (1981) Regression of Hodgkin’s disease after measles. Lancet 1(8229): 1112.
-
Bluming AZ, Ziegler JL (1971) Regression of Burkitt’s lymphoma in association with measles infection. Lancet 2(7715): 105-106.
-
Heinzerling L, Künzi V, Oberholzer PA, Kündig T, Naim H, et al. (2005) Oncolytic measles virus in cutaneous T-cell lymphomas mounts antitumor immune responses in vivo and targets interferon-resistant tumor cells. Blood 106(7): 2287-2294.
-
Russell SJ, Federspiel MJ, Peng KW, Tong C, Dingli D, et al. (2014) Remission of disseminated cancer after systemic oncolytic virotherapy. Mayo Clin Proc 89(7): 926-933.
-
Galanis E, Hartmann LC, Cliby WA, Long HJ, Peethambaram PP, et al. (2010) Phase I trial of intraperitoneal administration of an oncolytic measles virus strain engineered to express carcinoembryonic antigen for recurrent ovarian cancer. Cancer Res 70(3): 875-882.
-
Samson A, Scott KJ, Taggart D, West EJ, Wilson E, et al. (2018) Intravenous delivery of oncolytic reovirus to brain tumor patients immunologically primes for subsequent checkpoint blockade. Sci Transl Med 10(422): eaam7577.
-
Komorowski M, Tisonczyk J, Kolakowska A, Drozdz R, Kozbor D, et al. (2018) Modulation of the tumor microenvironment by CXCR4 antagonist-armed viral oncotherapy enhances the antitumor efficacy of dendritic cell vaccines against neuroblastoma in syngeneic mice. Viruses 10(9): 455.
-
Tang B, Guo ZS, Bartlett DL, Liu J, McFadden G, et al. (2019) A cautionary note on the selectivity of oncolytic poxviruses. Oncolytic Virother 8: 3-8.
-
Waters AM, Johnston JM, Reddy AT, Fiveash J, Swain AM, et al. (2017) Rationale and design of a phase I clinical trial to evaluate HSV G207 alone or with a single radiation dose in children with progressive or recurrent malignant supratentorial brain tumors. Hum Gene Ther Clin Dev 28(1): 7-16.
-
Selman M, Ou P, Rousso C, Bergeron A, Krishnan R, et al. (2018) Dimethyl fumarate potentiates oncolytic virotherapy through NF-kappaB inhibition. Sci Transl Med 10(425): eaao1613.
-
Xiao X, Liang J, Huang C, Li K, Xing F, et al. (2018) DNA-PK inhibition synergizes with oncolytic virus M1 by inhibiting antiviral response and potentiating DNA damage. Nat Commun 9(1): 4342.
-
Fend L, Yamazaki T, Remy C, Fahrner C, Gantzer M, et al. (2017) Immune checkpoint blockade, immunogenic chemotherapy or IFN-alpha blockade boost the local and abscopal effects of oncolytic virotherapy. Cancer Res 77(15): 4146-4157.
-
Chen CY, Wang PY, Hutzen B, Sprague L, Swain HM, et al. (2017) Cooperation of oncolytic herpes virotherapy and PD-1 blockade in murine rhabdomyosarcoma models. Sci Rep 7(1): 2396.
-
O’Leary MP, Choi AH, Kim SI, Chaurasiya S, Lu J, et al. (2018) Novel oncolytic chimeric orthopoxvirus causes regression of pancreatic cancer xenografts and exhibits abscopal effect at a single low dose. J Transl Med 16(1): 110.
-
Baldo A, Galanis E, Tangy F, Herman P (2016) Biosafety considerations for attenuated measles virus vectors used in virotherapy and vaccination. Hum Vaccin Immunother 12(5): 1102–1116.
-
Peng KW, Frenzke M, Myers R, Soeffker D, Harvey M, et al. (2003) Biodistribution of oncolytic measles virus after intraperitoneal administration into Ifnar-CD46Ge transgenic mice. Hum Gene Ther 14(16): 1565-1577.
-
Kambara H, Saeki Y, Chiocca EA (2005) Cyclophosphamide allows for in vivo dose reduction of a potent oncolytic virus. Cancer Res 65(24): 11255-11258.
-
Myers RM, Greiner SM, Harvey ME, Griesmann G, Kuffel MJ, et al. (2007) Preclinical pharmacology and toxicology of intravenous MV-NIS, an oncolytic measles virus administered with or without cyclophosphamide. Clin Pharmacol Ther 82(6): 700-710.
-
U.S. National Library of Medicine ClinicalTrials.gov (2006) Viral Therapy in Treating Patients With Recurrent Glioblastoma Multiforme. Official title: Phase I trial of Measles virus derivative producing CEA (MV-CEA) in patients with recurrent Glioblastoma Multiforme (GBM).
-
Ferreira R, Sousa C, Gonçalves RFS, Pinheiro AC, Oleastro M, et al. (2022) Characterization and genomic analysis of a new phage infecting Helicobacter pylori. Int J Mol Sci 23(14): 7885.
-
Strathdee SA, Hatfull GF, Mutalik VK, Schooley RT (2023) Phage therapy: From biological mechanisms to future directions. Cell 186(1): 17-31.
-
Baldo A, van den Akker E, Bergmans HE, Lim F, Pauwels K, et al. (2013) General Considerations on the Biosafety of Virus-derived Vectors Used in Gene Therapy and Vaccination. Curr Gene Ther 13(6): 385-394.
-
Public Health Agency of Canada (2011) Laboratory Biosafety and Biosecurity. Pathogen Safety Data Sheets and Risk Assessment-Measles virus.
-
Directives (2009) Directive 2009/41/EC of the European Parliament and of the Council of 6 May 2009 on the contained use of genetically modified micro- organisms (Recast). Official Journal of the European Union, 125: 75-97.
-
Mader EK, Maeyama Y, Lin Y, Butler GW, Russell HM, et al. (2009) Mesenchymal stem cell carriers protect oncolytic measles viruses from antibody neutralization in an orthotopic ovarian cancer therapy model. Clin Cancer Res 15(23): 7246-755.
-
Li L, Liu S, Han D, Tang B, Ma J, et al. (2020) Delivery and Biosafety of Oncolytic Virotherapy. Front Oncol 10: 475.
-
Abdullahi S, Jäkel M, Behrend SJ, Steiger K, Topping G, et al. (2018) A novel chimeric oncolytic virus vector for improved safety and efficacy as a platform for the treatment of hepatocellular carcinoma. J Virol 92(23): e01386-18.
-
Bhaskar A, Bala J, Varshney A, Yadava P (2011) Expression of measles virus nucleoprotein induces apoptosis and modulates diverse functional proteins in cultured mammalian cells. PLoS One 6(4): e18765.
-
Bhattacharjee S, Jaiswal RK, Yadava PK (2017) Measles virus N and P proteins find interacting partners in HeLa cells. Int J Adv Res 5(7): 2691–2698.
-
Davis JJ, Fang B (2005) Oncolytic virotherapy for cancer treatment: challenges and solutions. J Gene Med 7(11): 1380-1389.
-
Chen A, Zhang Y, Meng G, Jiang D, Zhang H, et al. (2017) Oncolytic measles virus enhances antitumour responses of adoptive CD8+NKG2D+ cells in hepatocellular carcinoma treatment. Sci Rep 7(1): 5170.
- hMPV: Is It Another Covid-19 Like Situation?
- Streptomyces: Sources of Novel Discoveries in Antibiotic Research to Combat Antimicrobial Resistance
- A Review of Mosquitoes (Diptera: Culicidae) and Their Biodiversity, Medical and Veterinary Importance
- Past and Current Immunotherapy in Cancer
- Hematological Cancer and Viral Infection
- The Growing Threat of Antimicrobial Resistance in India: Challenges and Solutions