First Mention of Cannibalism in Salamandra Algira (Caudata, Salamandridae) Larvae in Algeria with Some Ecological Notes
Animal cannibalism is a practice well known to scientists, it consists of partially or entirely eating a living individual of the same species by necessity or by instinct. Although originally thought to be abnormal behavior, it has now been recorded in over 1300 species across a diverse group of taxa, ranging from protists and invertebrates to humans. This type of behavior exists in amphibians in different forms and it is observed in 12 families of toads, 7 families of salamanders and 1 family of gymnophionas (Lissamphibians) and this phenomenon is known in natural populations of Salamandridae larvae. The only representative of this family and genus Salamandra in North Africa is Salamandra algira with five subspecies, of which only one is reported for Algeria (Salamandra algira algira) and cannibalism among Salamandra algira larvae has been described for Morocco at Jbel Tazzeka in the Middle Atlas where gigantism has also been observed in certain larvae whose size can vary from 25 to 74 mm. In this study we report for the first time the practice of cannibalism in Algeria in S.A. algira and we also observed gigantism in some larvae. Regarding salamanders, there is still a lot to do because their distribution and ecology are not well known.
Introduction
Animal cannibalism is a practice that is beginning to be well known to scientists, it consists of partially or entirely eating a living individual of the same species by necessity or by instinct. Mathematical models and behavioral studies present cannibalism as a way to maximize individual profitability: it is often an adaptation that compensates for food limitation or extreme competition for the same resources. Although originally thought to be abnormal behavior, cannibalism has now been recorded in over 1300 species across a diverse group of taxa, ranging from protists and invertebrates to humans. It would act as an important factor in regulating population dynamics. For Van allen, et al [1]. cannibalism occurs in a majority of both carnivorous and non-carnivorous animal taxa from invertebrates to mammals. Similarly, infectious parasites are ubiquitous in nature. Thus, interactions between cannibalism and disease occur regularly. While some adaptive benefits of cannibalism are clear, the prevailing view is that the risk of parasite transmission due to cannibalism would increase disease spread and, thus, limit the evolutionary extent of cannibalism throughout the animal kingdom. In contrast, surprisingly little attention has been paid to the other half of the interaction between cannibalism and disease, that is, how cannibalism affects parasites. Here we examine the interaction between cannibalism and parasites and show how advances across independent lines of research suggest that cannibalism can also reduce the prevalence of parasites and, thus, infection risk for cannibals.
Alabi, et al. [2] by studying the origin and evolution of cannibalism in animal populations had to analyze this kind of behavior in amphibians. They quote and agree with Elgar, et al. who said amphibians can have different forms of cannibalism which are: AA: adults consuming adults — adults cannibalizing adults; AJ: adults consuming juvenile stages — adults cannibalizing juveniles; AE: adults consuming eggs — adults cannibalizing eggs; JJ: juvenile stages consuming stages juveniles — juveniles cannibalizing juveniles; JE: juvenile stages consuming eggs — juveniles cannibalizing eggs. The causes of cannibalism in amphibians are variable. Cannibalism can be induced by too abundant fertility, with the corollary a small offspring, more vulnerable to predation; competition between individuals induced by a high population density. Ambystoma tigrinum larvae eat each other once the frequency encounter is high [3]. The drying up of egg-laying sites associated with an increase in density and a decrease in resources; polyphenism, i.e. discrete phenotypic differences in morphology between individuals in the population. This morphological difference results in a variation in the rate of growth and behavior between individuals, some grow faster and develop a morphology adapted to cannibalism, others develop normally [4, 5]. The species Ambystoma tigrinum [6]. Hynobius retardatus [7]. Scaphiopus couchii [8, 9], are characterized by large- headed and wide-mouthed cannibalistic individuals compared to to body size. Some individuals could have an intermediate morphology between that of a cannibalistic individual and that of a non-cannibalistic individual [5]. This heterogeneity of growth in batrachians finds its explanation for some through genetic factors resulting in differences in the frequency of allozyme in cannibal and non-cannibal individuals [5, 10], and for others through environmental factors [3]. However, according to Nishihara, Hoffman and Pfennig [11, 12], Nishihara-Takahashi and Kohmatsu, et al. [13, 14], cannibal morphology results from phenotypic plasticity linked to environmental conditions.
According to Crump [4], cannibalism is observed in 12 families of toads, 7 families of salamanders and 1 family of gymnophionas (Lissamphibians). Larval cannibalism is widespread among amphibians, especially in urodeles and anurans including species of the families Ambystomatidae (Ambystoma annulatum), Cryptobranchidae (Andrias japonicus), Dicamptodontidae (Dicamptodon ensatus), Hynobiidae (Hynobius nebulosus) Myobatrachidae (Lechriodus fletcheri) and Pelobatidae (Scaphiopus bombifrons) are representative of the behavior. Cannibalism has already been reported by different authors in different species of salamanders. Pfenning, et al. [15] suggest that pathogen transmission is an important cost of cannibalism on the tiger salamander Ambystoma tigrinum and provide a general explanation for why cannibalism is infrequent in most species and in 1999 Pfenning, et al. [16] mentions that in the larvae cannibalism of this species discriminates kin. This phenomenon has also been reported in Hynobius retardatus by Kishida, et al. [17] who mention that cannibalism plays a role in adaptively accelerating the growth and development of salamander hatchlings.
The genus Salamandra Garsault is a group of terrestrial Urodeles widely distributed across the western Palearctic, comprising six species: Salamandra algira Bedriaga, S.atra Laurenti, S.corsica Savi, S.infraimmaculata Martens, S.lanzai Nascetti, Andreone, Capula and Bullini and S. salamandra [18, 19]. For Degani, et al. [20] the phenomenon of cannibalism is known in natural populations of Salamandra salamandra larvae inhabitating either rain pools in rocks or permanent springs poor in aquatic life. It was found that cannibalism increased under experimental conditions proportionally to the increase in size between the larvae.
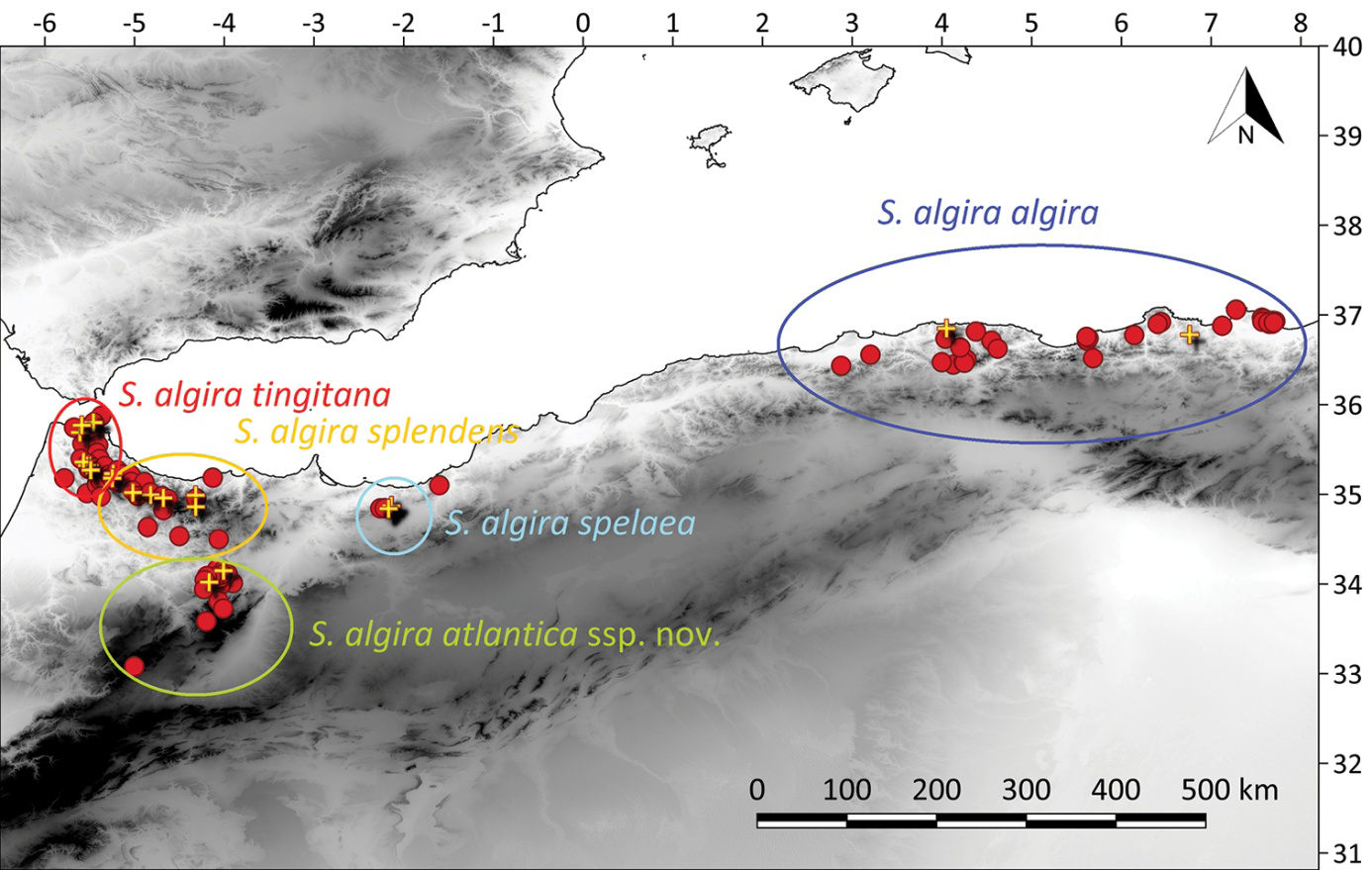
Hernandez and Escoriza [21] stipulate that the sole representative of the genus in north Africa is Salamandra algira [19] and four subspecies are currently recognised [22, 23, 24] S.algira tingitana in the north-western Rif from Ceuta to Moulay Abdessalam ; S.algira splendens, in the central- eastern Rif, from Chefchaouen to Al Hoceima, including isolated populations in the middle Atlas Mountains ; S.algira spelaea restricted to the Beni Snassen massif, and the nominal S.algira.algira in Algeria, located across the pericoastal mountains of Annaba,Collo,Kabylia,and Blida Atlas [23, 25, 26, 27]. Ben Hassine, et al. and Dinis, et al. [28] suggested that the taxonomy of S.algira could require further re-asseeements and S algira splendens includes two distinct allopatric clades separated by 50 km of semi-arid plains and they described one of these clades as a distinct subspecies, based on genetic and morphological evidences (Figure 1).
Cannibalism among Salamandra algira larvae is probably more common in North Africa than we observe. This has been described for Morocco by Escoriza, et al. [29] or in November 2004 they observed this phenomenon at Jbel Tazzeka in the Middle Atlas and they also observed gigantism in certain larvae in which the size can vary from 25 to 74 mm. They observed 1 giant larva which has a length of 74 mm and 18 typical larvae with a length of 41 mm. Bogaerts mentioned in a personal communication in 2023 that he also saw it in Jebel Tazzeka but it has not been described for Algeria where there is still much to study in Algeria, because the distribution nor the ecology do not are not well known.
Material and Methods
On November 8, 2022 while working in the forest, we noticed the presence of a colony of salamander larvae in a fountain in the form of a basin and we undertook to monitor the behavior of the larvae. The basin is located in the forest of Beni Mimoune in the municipality of Tichy and department of Bejaia a coastal town situated at 250 East of Algeirs , at an altitude of 600 meters at a place called Tala Merzouk ( 36.6605091 E – 5.1364423 N). The Beni Mimoune forest is a cork oak forest (Quercus suber) with undergrowth characteristic of coastal areas (Pistacia lentiscus, Rubus The basin is oriented North-East with exposure to the Mediterranean Sea, and it is in the form of two compartments of 120 cm long, 60 cm wide and 80 cm deep. From November 2022 to January 2023 it was not supplied by the spring because it was dry and it was only from the end of January that there was running water, which spilled outward. The pool water had supernatant algae on the surface. We carried out 23 work trips from November 2022 to June 2023. On each trip we counted the number of individuals in the basin and observed the behavior of the larvae.

| Year | 2022 | 2023 | ||||||
|---|---|---|---|---|---|---|---|---|
| Month | Nov | Dec | Jan | Feb | Mars | Apr | May | June |
| Number of work trips | 1 | 6 | 5 | 3 | 5 | 1 | 1 | 1 |
Table 1: Schedule of work in the Tala Merzouk basin.
Results and Discussion
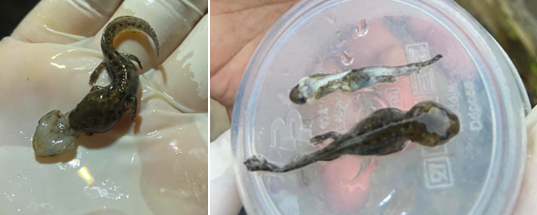
During the first outing in November 2022 we had to notice the presence of 180 larvae, the size of which was identical for the majority and was 25 -30 mm. From January 2023 we had to notice that some larvae grew and reached 40- 45 mm but others remained at their initial size of 25-30 mm. The first observation of the phenomenon of cannibalism was made during the month of January. We noticed two different modes of cannibalism which showed that large larvae (40-45 mm) consumed larvae of the same size and there were also larvae (40-45 mm) which consumed larvae of small size (25- 30 mm).
We also had to observe other phenomena such as the presence of several larvae (40-45 mm) which fed on one and the same individual having the same size. The second phenomenon is that certain larvae when they are stressed start to regurgitate the larva they have eaten, this phenomenon has been observed several times and it has been noticed that the regurgitated larva was emptied of its blood but not of its viscera.
Another observation was made concerning the consumption of Myriapods (undetermined) by a small larva (25-30 mm) and the myriapod had a size of 60 mm (Figure 3).



From the month of February there were only almost half of the individuals left (80-90) we do not know if all the missing individuals were consumed or there were some individuals who were consumed by other predators, this remains to be verified because the larvae in general are in the dond of the basin which is 80 cm deep. We have hypothesized that some individuals were swept away by the waters that overflowed the basin and ended up on the ground and were eaten by other predators. We report that we only saw one Muridae once near the basin.
We also point out that it was from the month of March that we observed the first giant larvae exceeding 70 mm and already possessing the characteristic yellow pigmentations of Salamandra algira, and it was from this month that we found in the basin 3 kinds of larvae which are small larvae (25 -30 mm), medium size larvae (40-45 mm) and giant larvae exceeding 70 mm. It is therefore obvious that in Salamndra algira in Algeria, the larvae do not grow in the same way and this cannot be due to a lack of food availability because there was a constant presence of food.
We did an experiment to find out if cannibalism is due to a lack of food, we took two samples of 3 individuals of different sizes (1 of 25-30 mm, 1 of 40-45 mm and one of 70 mm). The first sample was deposited at the research laboratory in ecology and environment of the University of Bejaia and the second sample was deposited at my home. Both samples did not receive food and the result is that after 10 days the small larva (20-25 mm) died in both samples and we left it until the 20th day when the medium larva size (40- 45 mm) died at the laboratory level and the average larva taken from me died after the 25th day. The giant larvae (70 mm) did not consume the dead larvae in the two samples. This would imply that cannibalism in S. algira in Algeria is not due to lack of food since the giant larva had no direct food but it could consume the larvae that died and it did not. After ten days we took the giant larvae back and put them back in their natural environment at the level of the Tala Merzouk basin.
If we stick to the work of Pfennig he shows that in salamanders cannibalism is a form of helping the individual relatives and the research supports the “inclusive fitness theory” which explains why cannibalism is the salamander method to help loved ones. When the salamander becomes cannibalistic and eats non-relatives it removes some of the competition for valuable resources because by eating the non-relatives the relatives have more food available and It suggests that the smell might be the way salamanders discriminate parents from non-parents, and cannibalism is a way to speed development by feeding on protein-rich prey right next to you in the pond. This is probably what we observed in S. algira algira because the larvae we studied come from the same mother.
Conclusion
It is accepted that cannibalism exists in Salamanders and different authors have reported it in different species around the world. In North Africa where there is only one species (Salamandra algira) represented by 5 subspecies, S. algira tingitana, S.a.splendens, S.a.spelaea, S.a.atlantica and S.a.algira. This last subspecies is the only one that lives in Algeria and with our present work we show that it is a subspecies that has the potential to become cannibalistic and it is the first time that we observe this phenomenon in this species in Algeria. Further studies could shed light on this phenomenon on this species in Algeria to allow us to better protect this salamander which is classified as Vulnerable by the IUCN.
References
-
Van Allen BG, Dillemuth FP, Flick AJ, Faldyn MJ, Clark DR, et al. (2017) Cannibalism and infectious disease: friends or foes? The american naturalist. University of Chicago Press 190 (3): 299-312.
-
Alabi T, Patiny S, Verheggen, Francis F, Haugruge E (2009) Origin and evolution of cannibalism in animal populations: why do they eat similar things?. Univ Liege. Biotechnol Agron Soc Environ 13(3): 409-425.
-
Collins J, Cheek J (1983) Effect of food and density on development of typical and cannibalistic salamander larvae in Ambystoma tigrinum nebulosum. Am Zool 23(1): 77-84.
-
Crump ML (1992) Cannibalism in amphibians. In: Elgar M, Crespi B (Edn.), Cannibalism: ecology and evolution among diverse taxa. Oxford: Oxford University Press, UK, pp: 256-276.
-
Wakano JY (2003) Drastic growth effect may explain sympatric cannibalistic polymorphism. J. Theor. Biol 226(1): 69-77.
-
Rose FL, Armentrout D (1976) Adaptive strategies of Ambystoma tigrinum Green inhabiting the Llano Estacado of West Texas. J Anim Ecol 45: 713-729.
-
Wakahara M (1995) Cannibalism and the resulting dimorphism in larvae of a salamander Hynobius retardatus, inhabited in Hokkaido. Jpn Zool Sci 12: 467- 473.
-
Pfennig DW (1990) The adaptive significance of an environmentally cued developmental switch in an anuran tadpole. Oecologia 85: 101-107.
-
Dayton GH, Wapo SD (2002) Cannibalistic behaviour in Scaphiopus couchii: more evidence for larval anuran oophagy. J Herpetol 36: 531-532.
-
Pierce B, Mitton J, Jacobson L, Rose FL (1983) Head shape and size in cannibal and noncannibal larvae of the tiger salamander from West Texas. Copeia pp: 1006-1012.
-
Nishihara A (1996) Effects of density on growth of head size in larvae of the salamander Hynobius retardatus. Copeia pp: 478-483.
-
Hoffman E, Pfennig D (1999) Proximate causes of cannibalistic polyphenism in larval tiger salamanders. Ecology 80: 1076-1080.
-
Nishihara-Takahashi A (1999) Faster growth of head size of prefeeding larvae in a cannibalistic population of the salamander Hynobius retardatus. Zool Sci 16(2): 303- 307.
-
Kohmatsu Y, Nakano S, Yamamura N (2001) Effects of head shape variation on growth, metamorphosis and survivorship in larval salamanders (Hynobius retardatus). Ecol Res 16: 73-83.
-
Pfenning DW, Ho SG, Hoffman EA (1998) Pathogen transmission as a selective force against cannibalism. Animal Behaviour 55(5): 1255-1261.
-
Pfennig DW, Collins JP, Ziemba RE (1999) A test of alternative hypotheses for kin recognition in cannibalistic tiger salamanders. Behavioral Ecology 10(4): 436-443.
-
Kishida O, Tezuka A, Ikeda A, Takatsu K, Michimae H (2014) Adaptive acceleration in growth and development of salamander hatchlings in cannibalistic situations. Functional ecology. British ecological society 29(4): 469- 478.
-
Dubois A, Raffaelli J (2009) A new ergotaxonomy of the family Salamandridae Goldfuss, 1820 (Amphibia, Urodela) Alytes 26(1-4) : 1-85.
-
Escoriza D, Ben Hassine J (2019) Amphibians of North Africa. Academic Press, London, Amphibians of North Africa pp 11-28.
-
Degani G, Goldenberg S, Warburg MR (1980**)** Cannibalistic phenomena in Salamandra salamandra larvae in certain water bodies and under experimental conditions. Hydrobiologia 123-128.
-
Hernandez A, Escoriza D (2019) A new subspecies of African fire salamander Salamandra algira (Urodela, Salamandridae) from the Middle Atlas Mountains, Morocco. Zookeys 893: 143-158.
-
Donaire-Barroso D, Bogaerts S (2003) A new subspecies of Salamandra algira Bedriaga, 1883 from northern Morocco. Podarcis 4: 84-100.
-
Escoriza D, Comas MM (2007) Description of a new subspecies of Salamandra algira Bedriaga, 1883 (Amphibia : Salamandridae) from the Beni Snassen massif (Notheast Morocco). Salamandra 43(2): 77-90.
-
Beukema W, De Pous P, Donaire-Barroso D, Bogaerts S, Garcia-Porta J, et al. (2013) Rewiew of the systematics, distribution, biogeography and natural history of Moroccan amphibians. Zootaxa 3661:1-60.
-
Beukema W, Donaire D, Escoriza D, Bogaerts S, Toxopeus AG, et al. (2010) Biogeography and contemporary climatic differentiation among Moroccan Salamandra algira. Biological Journal of the Linnean Society 101(3): 626-641.
-
Escoriza D, Ben Hassine J (2014) Microclimatic variation in multiple Salamandra algira population along an altitudinal gradient: phenology and reproductive strategies. Acta Herpetologica 9(1): 33-41.
-
Hernandez A, Escoriza D (2017) Easternmost record of Salamandra algira splendens in Morocco. Boletin de la Asociacion Herpetologica Espanola 28(1): 60-61.
-
Ben Hassine J, Gutierrez-Rodriguez J, Escoriza D, Martinez-Solano I (2016) Inferring roles of variance, climate and topography in population differentiation in Salamandra algira (Caudata, Salamandridae). Journal of Zoological Systematics and Evolutionary Research 54(2): 116-126.
-
Escoriza D, Comas MM, Donaire D (2006) Gigantismo larvario en Salamandra Algira Bedriaga 1883. Descripcion de un caso. Butleti de la Societat Catalana d’Herpelogia 17: 59-63.
-
Martinez-Freiria F, Steinfartz S, Vences M, Burgon JD, Elmer KR, et al. (2019) Allopatric diversification and evolutionary melting pot in North African Palearctic relict : the biogeographic history of Salamandra algira. Molecular Phylogenetics and Evolution 130: 81-91.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind